Evidências da pegada da diversidade locomotora dos primeiros hominíneos em Laetoli, Tanzânia
Nature volume 600 , páginas 468–471 ( 2021 )
Resumo
Rastros bípedes descobertos em 1978 no sítio G de Laetoli, na Tanzânia e datados de 3,66 milhões de anos atrás, são amplamente aceitos como a mais antiga evidência inequívoca de bipedalismo obrigatório na linhagem humana 1 , 2 , 3 . Outra trilha descoberta dois anos antes no local próximo A foi parcialmente escavada e atribuída a um hominídeo, mas curiosas afinidades com ursos (ursídeos) marginalizaram sua importância para a comunidade paleoantropológica, e a localização dessas pegadas caiu no esquecimento 3 , 4 , 5 . Em 2019, localizamos, escavamos e limpamos a trilha do local A, produzindo um arquivo digital usando fotogrametria 3D e digitalização a laser.
Aqui, comparamos as pegadas neste local com as de ursos negros, chimpanzés e humanos americanos, e mostramos que elas se parecem mais com as dos hominídeos do que com os ursídeos. Na verdade, a largura estreita do passo corrobora a interpretação original de um hominídeo bípede pequeno e cruzado. No entanto, as proporções inferidas do pé, os parâmetros da marcha e as morfologias 3D das pegadas no local A são facilmente distinguidas daquelas no local G, indicando que um mínimo de dois taxa de hominíneos com pés e andamentos diferentes coexistiram em Laetoli.
Principal
Em 1976, Peter Jones e Philip Leakey descobriram cinco pegadas bípedes consecutivas no sítio A de Laetoli na localidade 7, uma 490 m 2 área de datada de 3,66 milhões de anos atrás (Ma) e com 18.400 pegadas de animais 1 , 2 , 3 (Fig. 1 ) . Mary Leakey sugeriu provisoriamente que a trilha foi feita por um hominídeo 1 . “As pegadas”, escreveu ela, “indicam uma marcha rolante e provavelmente lenta, com os quadris girando a cada passo, em oposição à marcha livre do homem moderno [humanos].” Leakey e Hay 2 classificaram as pegadas como Hominidae, mas com uma ressalva de que “a marcha era um tanto cambaleante, com um pé cruzando na frente do outro”.

a , Um modelo do local A gerado usando fotogrametria mostrando as cinco pegadas de hominídeos. b , Mapa de contorno correspondente do local gerado a partir de uma varredura de superfície 3D com barra de escala. c , Mapa das localidades Laetoli 7 e 8, indicando as posições das vias bípedes A, G e S (redesenhado da ref. 49 ). d , e , Mapas topográficos das duas pegadas A mais bem preservadas, A2 ( d ) e A3 ( e ).
Pegadas de hominídeo inequívocas foram descobertas no local G dois anos depois, lançando dúvidas sobre o status de hominídeo daqueles no local A 3 , 4 , 5 . Os pesquisadores descreveram as pegadas no local A como "muito incomuns", 6 "de formato curioso" 7 e "enigmáticas" 8, mas o consenso era uniforme: elas foram produzidas por um mamífero plantígrado que se movia bípede.
Tuttle 4 avançou três hipóteses para explicar a morfologia das pegadas e da marcha em cruz (isto é, quando um pé de cada lado cruza a linha média antes do toque): (1) distorção do substrato; (2) foram deixados por um urso juvenil (ursid); ou (3) são evidências de outra espécie de hominídeo. Para testar a segunda possibilidade, Tuttle 4 , 9 , 10 , 11 , 12 coletou dados de ursos de circo treinados para andar bípede e descobriu que seus passos curtos e pés relativamente largos eram semelhantes às pegadas do local A, embora os ursos bípedes levem mais largos degraus. Além disso, o quinto dígito é tipicamente o maior em ursídeos, resolvendo o 'problema de cross-stepping', embora Tuttle 4 , 12 tenha notado que os humanos ocasionalmente cross-step. Ele concluiu que “até que estudos biométricos e cinesiológicos naturalísticos detalhados sejam realizados em ursos bípedes e humanos descalços, teremos que adiar a escolha entre as hipóteses de hominídeo e ursídeo no indivíduo Laetoli A” 4 .
Para complicar ainda mais as coisas, a morfologia interna das pegadas do local A nunca foi totalmente limpa do preenchimento da matriz 1 , 2 , 4 , 5 , 12 . White e Suwa 8 argumentaram que “a identificação confiável dessas impressões enigmáticas no local A de Laetoli será impossível até que sejam mais completamente limpas e acompanhadas lateralmente”. Assim, ficamos motivados a realocar e reescavar o local A e realizar uma análise comparativa detalhada das pegadas, bem como da locomoção de ursos ( Ursus americanus ), chimpanzés ( Pan troglodytes ) e humanos para testar se as pegadas no local A eram deixada por um hominídeo ou ursídeo.
Redescoberta do site A
Usando mapas detalhados de Leakey e Harris 3 , identificamos a trilha proboscidiana adjacente às pegadas bípedes. Limpamos a cobertura circundante até que um de nós (KF) encontrou a pegada do A3. A área foi então limpa com escova para expor A1-A5, que não experimentou nenhuma erosão perceptível desde sua descoberta inicial (Fig. 1 , Dados Estendidos Fig. 1 ). Como o tufo da pegada está erodido ao norte, escavamos ao sul (87 cm) e ao leste (54 cm) a partir do calcanhar de A1, mas nenhuma pegada adicional foi encontrada ( informações suplementares ).
Depois de escovar o sedimento de A3, usamos um abaixador de língua de madeira para remover o enchimento de tufo deixado intacto durante as temporadas de campo de 1976–1978. A impressão alucial é claramente definida e tem cerca de 30 mm de largura. Crucialmente, expusemos a impressão do segundo dígito (Extended Data Fig. 2 ). Removemos o enchimento de A2, mas não podíamos fazê-lo completamente sem risco de danos. No entanto, o salto e a impressão alucial são claros. As informações detalhadas das outras pegadas (A1, A4 e A5) são limitadas a estimativas de comprimento, largura e comprimento do passo.
Embora a qualidade da preservação varie dentro e entre A1-A5, não há evidências de que as métricas biologicamente informativas tenham sido afetadas pela distorção do substrato. Rastros adjacentes e misturados de outros animais (variando em tamanho de galinhas d'angola a elefantes) não mostram evidências de distorção em seus perímetros ou morfologias internas. Dado que as superfícies da pista provavelmente representam escalas de tempo de horas a dias 13 , 14 , 15 , é parcimonioso inferir condições semelhantes de substrato e processos tafonômicos durante a formação da impressão e épocas subsequentes.
Avaliação das hipóteses de ursídeos e hominídeos
Gravamos 50,9 h de vídeo do comportamento do urso preto americano selvagem. Postura bípede sem suporte e locomoção ocorreram apenas 0,09% do tempo total de observação, do qual 59% foi postural e 41% foi locomotor (Dados Estendidos Fig. 3 ). Em apenas um caso, um urso deu quatro passos bípedes sem ajuda. Assim, supondo que nossos achados sejam generalizáveis a outros ursídeos, a probabilidade de observar quatro passos bípedes consecutivos é de 0,003%. A baixa frequência desse comportamento e a ausência de passos de transição quadrúpede-bípedes tornam improvável, mas não impossível, que o bipedalismo ursídeo tenha sido preservado no local A. Além disso, Laetoli é desprovido de fósseis ursídeos, apesar da recuperação de mais de 25.000 fósseis atribuído a 85 espécies de mamíferos 16 , 17 . Se estivessem presentes, os ursídeos eram raros na paisagem. Embora os conjuntos de pegadas possam incluir um número surpreendente de rastros de táxons cujos fósseis de esqueletos são raros (por exemplo, frequências relativamente altas de rastros de pássaros em Laetoli 3 e em locais de 1,5 ma perto de Ileret, Quênia 14 , 18 ), não há uma explicação tafonômica clara de por que rastros de ursídeos estariam presentes, mas seus fósseis ausentes.
Além disso, medimos 46 pegadas de quatro ursos negros selvagens jovens bípedes especificamente escolhidos porque o comprimento de seus pés (média = 145,7 mm) estava dentro de 10% do comprimento das pegadas do local A (média = 161,7 mm). Além disso, medimos as pegadas de chimpanzés produzidos durante o quadrupedalismo ( n = 54 de 46 adultos; Ngamba Island Chimpanzee Sanctuary, Uganda) e durante o bipedalismo ( n = 44 de dois subadultos; Stony Brook University, EUA). Comparamos esses dados com pegadas humanas descalças produzidas em três condições: (1) calçados habituais ( n = 654) caminhando em uma esteira plantar de pressão 19 ; (2) habitualmente descalço ou minimamente calçado ( n = 41) caminhar na lama deformável 20 , 21 ; e (3) faixas do Pleistoceno Superior ( n = 113) de Engare Sero, Tanzânia, formadas em cinzas vulcânicas retrabalhadas 22 , 23 (resumo na Tabela Dados Estendidos 1 de ).
Concordamos com outros 4 , 12 que as proporções das dimensões da pegada (por exemplo, largura do calcanhar e antepé) e o comprimento do passo observado no local A estão dentro da faixa ursid (dados estendidos Fig. 4a, b ). No entanto, para essas mesmas medidas, o local A também é semelhante a um chimpanzé e moderadamente semelhante às pegadas de hominídeo definitivas dos locais G e S. Segue-se que o indivíduo do local A estava dando passos curtos - como ocorre quando os humanos andam lentamente ou sobre um substrato escorregadio 24 - não que a marcha fosse semelhante a um ursídeo.
Com o preenchimento adicional removido de A2 e A3, as dimensões do perímetro são decididamente semelhantes a hominídeos, com impressões de calcanhar largas em relação à largura do antepé (Dados estendidos Fig. 4c ). Em contraste, chimpanzés e ursos têm saltos relativamente estreitos. Além disso, com os rastros totalmente escavados e limpos, não encontramos evidências de impressões de garras, embora às vezes estejam ausentes nas pegadas de ursídeo 12 . Aqui, as impressões estavam ausentes em 31% das pegadas ursid. Para testar se A3 foi produzido por um pé esquerdo hominídeo ou um pé direito ursídeo, comparamos a largura do hálux com o segundo dígito nas humanos ( n = 30) e chimpanzés ( n pegadas de = 50), e o quinto dígito com o quarto em urso ( n faixas de = 5). As impressões do dedo do pé A3 correspondem às proporções distintas de humanos e chimpanzés, em vez das dos ursos (dados estendidos Fig. 5 ).
Ao estabelecer que A3 é um pé hominídeo esquerdo, podemos agora confirmar que ocorreu o passo cruzado. Cross-stepping nunca foi observado em nossa amostra comparativa, mas humanos o fazem ocasionalmente 25 , 26 como uma estratégia compensatória para restabelecer o equilíbrio após uma perturbação 27 . Na verdade, sugerimos que a etapa cruzada apóia a hipótese de que as pegadas do local A foram deixadas por um hominídeo. O passo cruzado é improvável, e talvez impossível, quando os ursos ou chimpanzés caminham bípede. Eles produzem grandes excursões médio-laterais de seu centro de massa 28 e andam com quadris altamente abduzidos 29 , resultando em uma alta proporção entre a largura da passada e o comprimento do passo. Por outro lado, o passo cruzado humano é possibilitado por seu centro de massa médio-lateral reduzido e movimentos corporais, quadris aduzidos e ângulo bicondilar (isto é, joelhos valgos), resultando em uma baixa proporção correspondente, conforme expresso em todas as pistas em Laetoli.
The relative step widths of footprints from sites G and S fall squarely within the modern human distribution (Extended Data Figs. 4d, 5). The site A footprints lie outside the distributions of humans, chimpanzees and bears but are most like humans. This result indicates that the maker of the site A footprints had either valgus knees or adducted hips, or both. The presence of either bipedal trait argues for a hominin maker of the site A footprints.
Which hominin?
É geralmente aceito que o Australopithecus afarensis produziu as pegadas nos locais G e S 8 (mas consulte as referências 12 , 30 , 31 , 32 ). Portanto, é tentador atribuir as trilhas do local A a A. afarensis ; no entanto, essa premissa requer um exame da ontogenia do pé e da variação morfológica intraespecífica que leva em consideração a crescente evidência fóssil de diversidade locomotora (e presumivelmente taxonômica) entre os hominíneos do Plioceno (por exemplo, na ref. 33 ).
Com entre 101 e 104 cm de altura (das equações em Dingwall et al. 34 ), o criador das pegadas do site A era menor em altura do que outros trackmakers de Laetoli, que variavam de 111–116 cm (local G1) a 161–168 cm (site S1) 35 . É plausível que as pegadas do local A tenham sido feitas por um juvenil A. afarensis , mas essa hipótese é prejudicada por uma morfologia de pegada distinta das dos locais G ou S.
A proporção entre a largura e o comprimento do pé segue uma trajetória ontogenética diferente em humanos e chimpanzés; os pés humanos são consistentemente mais estreitos do que os pés de chimpanzé (Fig. 2 ). As pegadas feitas por humanos descalços, tanto dos tempos modernos quanto do Pleistoceno, são ligeiramente mais largas do que as feitas por humanos calçados em populações industrializadas. As pegadas não distorcidas dos locais G e S se enquadram na distribuição humana. A3 é mais semelhante a um chimpanzé por ser largo em comparação com seu comprimento (Fig. 2a , Dados Estendidos Fig. 2 ). Em chimpanzés, esta forma de pegada mais ampla é, em parte, impulsionada pela maior divergência do hálux. Assim, medimos a divergência alucial como uma proporção da distância entre o centro da impressão feita pelo primeiro e segundo dígitos e o comprimento da pegada. Para esta métrica, humanos e chimpanzés são claramente distintos. As pegadas do local G mais bem definidas se sobrepõem à distribuição humana, ao passo que a pegada A3 não - possui um hálux ligeiramente mais divergente do que os humanos e o local G, embora não tão divergente quanto as dos chimpanzés (Fig. Dados Estendidos 6 ). Este achado por si só não descarta um juvenil A. afarensis , visto que o pé de Dikika (Afar, Etiópia) tem um hálux ligeiramente mais divergente e móvel do que seus equivalentes adultos 36 .
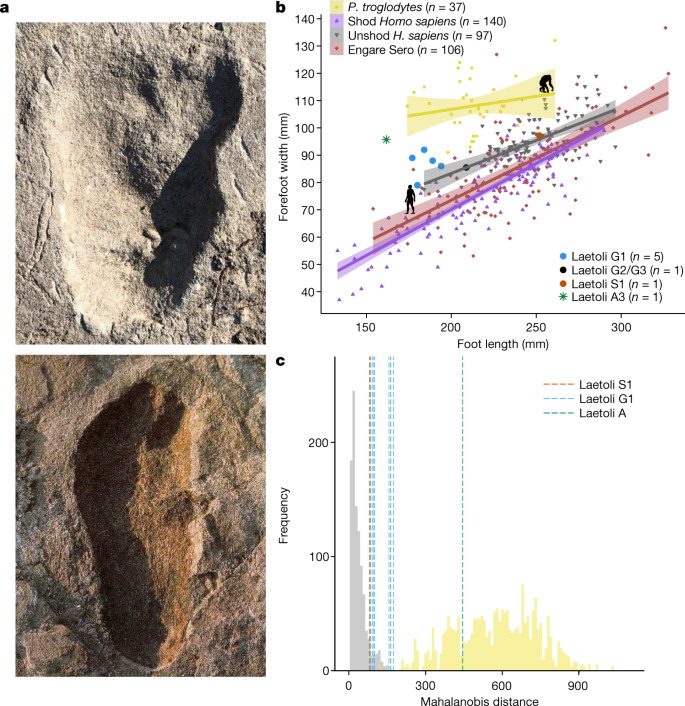
a, Images of Laetoli A3 (top) length matched to Laetoli G1 (bottom). G1 print is reproduced with permission from Science Photo Library. b, Plot comparing foot length to forefoot width in adult and juvenile humans and bipedal chimpanzees, the Pleistocene human footprints at Engare Sero, and the Laetoli trackways. Lines represent ordinary least-squares regression and shaded bands represent 95% confidence interval of the regression. Data were analysed from the total number of individual footprints indicated in the figure legend. Laetoli footprint markers are slightly enlarged for clarity. Data sources match Extended Data Table 1. c, Histogram of Mahalanobis distances between the mean modern human footprint and the averages of two randomly drawn human footprints (grey; n = 245, resampled 1,000 times) and two randomly drawn bipedal chimpanzee footprints (yellow; n = 45, resampled 1,000 times). The blue and orange lines represent the distances of mean Laetoli G1 (n = 5, with 10 unique two-track combinations) and S1 (n = 2) two-track samples, respectively. All Laetoli G1 and S1 samples fall within the human distribution. The green line indicates the distance between the mean Laetoli A track (n = 2) and the human mean, falling squarely within the chimpanzee distribution.
To explore other instructive traits, we compared proportional toe depth ratios as described by Raichlen and Gordon37 (Supplementary Methods). The mean value for site A (−0.191) is distinct from Laetoli G1 and humans using a bent-hip bent-knee gait but overlaps the low end of variation in Laetoli S. Additionally, tracks A1–A3 evince a raised ridge of hardened ash between the heel and lateral forefoot. It is unclear whether this ridge is evidence of substrate shearing38 or midfoot mobility—a characteristic absent from the Laetoli G and S footprints39 (but see ref. 40) and inconsistent with A. afarensis pedal remains41.
Finalmente, testamos se poderíamos amostrar pegadas aleatoriamente com topografia interna semelhante às várias pegadas de Laetoli de humanos ou chimpanzés (seguindo Hatala et al. 42 ). A Fig. 2 ilustra como as faixas dos locais G e S podem ser englobadas na faixa de variação da pegada humana sem ferragem reamostrada, enquanto a morfologia média das faixas A2 e A3 é distinta das pegadas de humanos habitualmente descalços e aquelas nos locais G e S. Na verdade, eles se encaixam confortavelmente na distribuição reamostrada do chimpanzé, sendo tão distintos quanto os rastros dos chimpanzés das morfologias dos rastros humanos descalços. Uma possível explicação para essa morfologia diferente da pegada é que as pegadas do local A foram feitas por um cruzadas A. afarensis em etapas . Testamos essa hipótese comparando as pegadas de humanos ( n = 10) caminhando com sua marcha preferida e depois cruzando os passos. Descobrimos que as pegadas humanas normais e cruzadas diferem minimamente e não correspondem em magnitude ou direção às diferenças entre as impressões do site G e S e as impressões do site A (Extended Data Fig. 7 , Informações Suplementares ).
Concluímos, portanto, que as pegadas do sítio A foram feitas por um hominídeo bípede com um pé distinto e presumivelmente mais primitivo do que A. afarensis . A forma grosseira do pé é semelhante à de um chimpanzé, com ligeira divergência alucial e talvez alguma mobilidade do mediopé. No entanto, o indivíduo no local A estava andando bípede com uma largura de passo estreita indicativa de um joelho valgo, quadril aduzido ou ambos. Esta combinação de morfologia do pé e cinemática da marcha inferida das pegadas preservadas impede que tenham sido feitas por A. afarensis .
Há evidências de diversidade taxonômica em hominídeos durante o Plioceno 43 , 44 , incluindo em Laetoli 45 , 46 , 47 , mas esses hominídeos não andavam com pés morfologicamente idênticos 48 . Por exemplo, o pé BRT-VP-2/73 do local de 3,4 Ma de Woranso-Mille, Etiópia, demonstra que pelo menos duas formas diferentes de pé coexistiram na Depressão Afar durante o Plioceno 33 . Sugerimos que a evidência da pegada para a diversidade locomotora dos hominíneos está igualmente presente em Laetoli, Tanzânia - e tem estado desde a descoberta da pista do local A na década de 1970.
Métodos
Dados comportamentais do urso selvagem
O comportamento do urso negro selvagem foi quantificado usando dados de vídeo gravados por BK ao longo de vários anos em seu local de campo em Lyme, New Hampshire, EUA. Os dados de vídeo capturaram ursos de diferentes idades (filhotes, adolescentes e adultos). Os ursos estiveram presentes na tela por um total de 50 h 55 min 18 s. Para cada incidente bípede terrestre, a duração do evento, a idade aproximada do urso e o número de passos foram registrados. Além disso, as etapas foram avaliadas se foram concluídas de forma independente ou se os indivíduos usaram outros objetos ambientais para o equilíbrio.
Dados cinemáticos comparativos
Dados cinemáticos comparativos foram coletados em três espécies: U. americanus , P. troglodytes e Homo sapiens . Para ursos e chimpanzés, o tamanho da amostra incluiu todos os indivíduos disponíveis alojados em cada local. Para obter informações sobre o tamanho da amostra humana, consulte abaixo. A randomização não foi relevante para nosso estudo, pois estávamos interessados em medir as características da pegada de populações inteiras de amostra, em oposição a comparações dentro dessas populações. O cegamento não foi relevante para os dados coletados nas espécies comparativas não humanas (por exemplo, ursos e chimpanzés) nem para a coleta de dados sobre pegadas fossilizadas. Os participantes humanos desconheciam as faixas do local A em Laetoli e, portanto, não sabiam como os dados obtidos de suas pegadas seriam usados neste estudo.
Ursus americanus
Data were collected on four juvenile semi-wild U. americanus (n = 3 male, 1 female), whose feet were within 10% of the length (average foot length = 145.7 mm) of the recorded footprints of Laetoli site A (average foot length = 161.7 mm). These orphaned, approximately 20-kg bears were located at the Kilham Bear Center (Lyme, NH), awaiting reintroduction to the wild. This study examined the bears between the ages of 5–8 months old. Our protocol was reviewed and approved by the Institutional Animal Care and Use Committee (IACUC) of Dartmouth College. The bears were enticed to independently walk bipedally through a constructed mud trackway for either an applesauce or maple syrup reward (Extended Data Fig. 3). Measurements were collected on the footprints, including foot length, heel width, forefoot width, step length and stride width using the definitions from Tuttle4. For a subset of footprints (n = 5), the width of the impression for the 1st, 2nd, 4th and 5th digits were measured. The presence or absence of claw impressions was also documented.
Pan troglodytes
Data for extant chimpanzees were extracted from three sources to collect all the relevant gait metrics. Two published datasets examined the same two subadult individuals housed at Stony Brook University. The third set were recorded on semi-wild individuals (n = 46), using a plantar pressure mat at the Ngamba Island Chimpanzee Sanctuary (Entebbe, Uganda). While this third data set increases sample size and captures intraspecific variation, we recognize that plantar pressure data do not always align perfectly with footprints made in a deformable substrate20.
Stride width data and step length comparisons
Os dados da largura da passada do chimpanzé foram retirados de Thompson et al. 28 em dois chimpanzés machos subadultos (7,0 ± 0,1 anos de idade; 34,8 ± 1,2 kg) e foram suplementados com dados de comprimento do passo para os mesmos passos. Métodos cinemáticos tridimensionais e cálculo de largura de etapa foram descritos anteriormente 28 . O comprimento do passo foi calculado como a distância entre os marcadores do calcâneo esquerdo e direito no plano sagital durante períodos consecutivos de meia postura dos membros posteriores. Os comprimentos dos passos do chimpanzé são tipicamente assimétricos, então o comprimento do passo foi a média dos dois passos consecutivos que definiram o passo.
Comparações da largura do antepé, largura do calcanhar e comprimento do pé
As dimensões da pegada e os dados do comprimento da passada foram registrados nos mesmos dois chimpanzés machos subadultos acima, embora em uma idade um pouco mais jovem (6,5 e 6,9 anos de idade, 30,7 e 27,8 kg, respectivamente). O projeto experimental é descrito em detalhes em outro lugar 42 . Em suma, os chimpanzés percorriam uma pista, no centro da qual havia uma esteira de pressão (RSScan International) e um contêiner de sedimento hidratado no qual os chimpanzés podiam produzir pegadas. Este sedimento foi retirado diretamente de uma camada que preserva pegadas de hominídeos de 1,5 Ma perto de Ileret, Quênia 50 . Câmeras de vídeo posicionadas lateralmente foram usadas para registrar os chimpanzés enquanto caminhavam ao longo da trilha e pegavam as pegadas. Dois softwares de digitalização, MaxTRAQ Lite + (v. 2.4.0.3) (Innovisions Systems) e ImageJ v.1.47 51 , foram usados para quantificar vários aspectos de seus passos, incluindo o comprimento da passada. Fitas métricas e compassos de calibre digitais foram usados para medir diretamente as dimensões externas dos pés de cada chimpanzé. Fotografias em escala foram tiradas das pegadas produzidas em cada ensaio e, posteriormente, medidas usando o software ImageJ.
Largura do antepé, largura dos dígitos 1 e 2, razão de divergência e comparações de comprimento do pé
Os dados foram coletados pelo EJM no santuário de Chimpanzés da Ilha Ngamba (Entebbe, Uganda) administrado pelo Chimpanzee Sanctuary and Wildlife Conservation Trust (CSWCT) usando procedimentos aprovados pelo Dartmouth College IACUC. O tapete de pressão plantar Tekscan (PPM) foi posicionado dentro de uma passarela conectando o recinto noturno ao habitat da floresta aberta, perto de um portão e sob uma seção transversal sólida para ajudar a evitar que os indivíduos pulassem sobre o tapete usando as barras do teto. Este local foi determinado usando a experiência dos guardiões do santuário. Os animais foram apresentados pela primeira vez a um tapete sem sensores internos, para habituá-los ao novo estímulo. Todos os dados subsequentes foram coletados usando os PPMs vazios e reais, posicionados de forma que cobrissem toda a largura da passarela para forçar os indivíduos a caminharem sobre um dos dois tapetes. Ambas as esteiras foram cobertas com sacos verdes finos para ajudar a disfarçá-los dos chimpanzés e facilitar a remoção mais rápida, se necessário. Determinou-se que a direção sudeste era o caminho preferido para os chimpanzés e o PPM contendo o sensor sempre foi posicionado lá a partir da segunda coleta. Os dados foram coletados duas vezes ao dia; uma vez pela manhã (entre 06:45 e 08:00), quando os chimpanzés se dirigiam à floresta para a primeira alimentação, e uma vez às 18:00, quando os chimpanzés iam para o recinto noturno para dormir e receber sua última alimentação . Os dados foram coletados em 46 chimpanzés adultos (18 machos, 28 fêmeas, com idades entre 12-36 anos). Um subconjunto de 54 registros de pressão dinâmica foi analisado. Usando o software Tekscan PPM associado, Footmat Research (v. 7.10), os registros de pressão foram analisados para determinar o comprimento do pé, a largura do antepé, os dígitos de largura 1 e 2 e a distância linear entre o centro dos dígitos 1 e 2. A divergência a razão foi calculada dividindo-se a distância entre os dígitos 1 e 2 pelo comprimento do pé do indivíduo.
Homo sapiens
Os dados foram extraídos de estudos anteriores de duas populações humanas modernas, a fim de coletar todas as métricas relevantes do pé, pegada e marcha.
Stride width and step length comparisons
Data were taken on 654 participants, recruited through the Living Laboratory at the Boston Museum of Science19. Sample size was determined by museum visitor traffic and willingness to participate in a scientific study. In brief, this dataset included 73 children between the ages of 2 and 7 years old (29 female and 44 male) and 581 individuals (366 female and 215 male) between the ages of 8 and 80 years. A pressure-sensitive gait carpet (6.1 m long × 0.89 m wide) with a spatial resolution of 1.27 cm and collecting data at 120 Hz (GAITRite) was used to collect stride length and stride width. For a subset of 33 adults, additional data were collected using a Tekscan PPM and analysed with FootMat Research (v. 7.10) to calculate foot length, the width of digits 1 and 2, and the linear distance between the centre of digits 1 and 2. These measurements were used to calculate a divergence ratio as described above in ‘P. troglodytes’.
Comparações da largura do antepé, largura do calcanhar e comprimento do pé
Dimensões da pegada e dados de comprimento da passada para 29 adultos Daasanach (15 homens e 14 mulheres, idades de 18 a 47) e 12 crianças (10 homens e 2 mulheres, idades de 4 a 15), que vivem perto da cidade de Ileret, Quênia e cresceram habitualmente descalço ou minimamente calçado foram retirados de Hatala et al. 20 , 21 . Os detalhes do protocolo experimental refletiram amplamente os procedimentos descritos acima. Em resumo, os indivíduos geraram pegadas enquanto caminhavam por uma amostra reidratada dos mesmos sedimentos que preservam rastros de hominídeos de 1,5 Ma perto de Ileret. Câmeras de vídeo foram usadas para registrar os indivíduos à medida que produziam pegadas, e dois pacotes de software de digitalização (MaxTRAQ Lite + v. 2.4.0.3 e ImageJ v.1.47) foram usados para medir o comprimento das passadas e outras variáveis cinemáticas. As dimensões externas dos pés dos sujeitos foram medidas diretamente com fitas métricas e compassos de calibre digitais. Fotografias em escala das pegadas produzidas em cada ensaio foram medidas usando ImageJ.
Experimentos humanos de pegada cruzada
Os experimentos foram realizados por KGH e EMW-H. para investigar se e como a cinemática cross-stepping influencia as dimensões do perímetro e topologias internas das pegadas de um indivíduo. Assim, poderíamos avaliar se o tamanho e a forma dos rastros do local A de Laetoli poderiam ter sido gerados por um hominíneo com pés semelhantes aos que deixaram rastros nos locais G e S, mas durante a travessia. Métodos detalhados são fornecidos nas Informações Suplementares . Resumindo, dez sujeitos adultos (incluindo seis mulheres, três homens e um não binário entre 19 e 52 anos de idade) completaram, cada um, dez testes nos quais produziram rastros em condições sedimentares destinadas a imitar os de Laetoli 37 , 52 . O tamanho da amostra foi determinado pela disponibilidade e vontade de participar do estudo. Cinco ensaios foram concluídos com uma marcha normal, auto-selecionada, e outros cinco foram concluídos com uma marcha cruzada, conforme inferido para o trackmaker do local A de Laetoli. Em cada ensaio, uma pegada focal foi selecionada, medida in situ e fotografada (25-30 fotos por pegada). Os comprimentos e larguras dos degraus que delimitam a pista também foram medidos. As fotografias foram usadas para gerar modelos 3D das trilhas usando o software Agisoft Metashape (v.1.7.3), e as trilhas normais e cruzadas médias foram geradas para cada sujeito usando o DigTrace Pro (v.1.8.1) 53 . Os comprimentos e larguras dessas faixas médias foram medidos usando Geomagic Wrap (v. 2021.0.0) (Sistemas 3D). As profundidades regionais foram medidas e avaliadas usando os mesmos métodos descritos abaixo ('Análises comparativas das formas da pegada de Laetoli'). As comparações dentro dos sujeitos nos permitiram entender como o cross-stepping influenciou as dimensões do perímetro e a topologia interna das pegadas de um sujeito.
Dados e análises de pegadas fósseis
Métricas comparativas foram quantificadas a partir de um conjunto de pegadas humanas modernas do Pleistoceno Superior em Engare Sero, na Tanzânia. Essas pegadas são uma comparação importante com as pegadas de Laetoli, pois foram geradas em uma circunstância semelhante (pegadas em cinzas vulcânicas) e representam uma população primitiva de humanos modernos descalços.
Uma ortofoto 3D em escala do site Engare Sero foi criada por meio de fotogrametria por BZ e CL-P. para visualizar a distribuição de rastros de footprint em todo o site usando o Agisoft Photoscan (agora Agisoft Metashape v. 1.4.4). O modelo foi criado a partir de centenas de fotos tiradas originalmente pelo Programa de Digitalização 3D Smithsonian em 2010. As medições que foram definidas no Tuttle 4 foram tiradas dos rastros fósseis usando o software ImageJ (v. 1.49) e incluíram o comprimento do pé, antepé largura, largura do salto e largura do passo de cada pegada na Engare Sero. Em alguns casos, pegadas parciais foram incluídas para medição, desde que incluíssem os pontos de referência necessários para essas medições. No geral, os dados foram coletados de 151 pegadas no site Engare Sero. Das 151 pegadas, 61 pegadas foram consideradas pegadas parciais e 90 pegadas foram consideradas pegadas completas. Destes, 67 medidas de comprimento do passo e largura da passada e 105 medidas de largura do calcanhar e largura da bola foram incluídos em nossas análises.
Todas as medições para as trilhas de Laetoli G e S foram obtidas de fontes publicadas 4 , 32 , 42 . Box e whisker plots e gráficos bivariados (usando ggplot2 54 ) foram gerados usando R (v. 3.6.1), enquanto a tabela e o gráfico de pizza foram gerados usando Microsoft Excel (v. 2102).
Análises comparativas das formas da pegada de Laetoli
As análises comparativas seguiram métodos semelhantes às análises de reamostragem publicadas anteriormente 42 . Em resumo, a amostra comparativa humana incluiu 245 pegadas produzidas por 29 indivíduos Daasanach adultos e 12 jovens habitualmente sem ferrugem viajando em velocidades de caminhada. A amostra comparativa do chimpanzé incluiu 45 pegadas produzidas por dois indivíduos caminhando bípedes. As amostras de Laetoli incluíram apenas as trilhas mais bem preservadas de cada local, deixando amostras de cinco pegadas do local G que foram descritas por suas escavadeiras originais como livres de danos tafonômicos que obscureceriam a topologia da trilha (G1-25, G1-27, G1-33 , G1-34 e G1-35), e dois do local S (L8-S1-2 e L8-S1-4). Para o site A, incluímos as trilhas A2 e A3, pois essas eram as únicas duas nas quais estávamos relativamente confiantes na identificação de regiões de interesse em toda a trilha. Tamanhos de amostra maiores seriam desejáveis, mas não queríamos sacrificar a qualidade dos dados pela quantidade, incluindo trilhas que foram impressas em excesso ou que não pareciam representar a anatomia completa do pé. Não confiamos em testes estatísticos paramétricos para os quais tamanhos de amostra maiores seriam uma necessidade e, em vez disso, usamos uma abordagem analítica que poderia lidar com conjuntos menores de observações (veja abaixo).
For each experimental and fossil footprint, 3D models were constructed using photogrammetry, through a variety of methods described here for Laetoli site A and elsewhere by the authors for other samples20,21,35,42. Using Geomagic Wrap (v. 2021.0.0) (3D Systems), a best-fit plane was fit to the undisturbed substrate surrounding each track, and this was fixed to the xy plane in world coordinate space. In this orientation, depths of the footprint were measured in the regions of the medial and lateral heel, medial and lateral midfoot, and all five metatarsal heads and toes. Raw depth measurements were normalized, within each footprint, to a scale of 0 to 1 in order to compare the topologies of footprints that may vary in depth. However, a Wilcoxon signed-rank test showed that, overall, human and Laetoli track samples did not differ significantly in their depths (P = 0.08). Within-subject means of the 14 normalized depth measurements were calculated, and a between-subject covariance matrix was created using the subject averages for normalized depths at each of the 14 measured regions. An overall ‘human mean footprint’ was also computed by averaging the within-subject mean normalized depths, and this represented a measure of central tendency as described below.
Para representar a faixa de variação observada na topografia da pegada humana, por 1.000 iterações, amostramos aleatoriamente um sujeito humano e extraímos uma amostra de duas de suas pegadas. Em seguida, calculamos a média das profundidades normalizadas dessas duas pegadas e calculamos a distância de Mahalanobis (usando a matriz de covariância entre sujeitos) entre esta trilha e a média das pegadas de todos os outros sujeitos. Além disso, para 1.000 iterações, selecionamos um chimpanzé aleatório, extraímos uma amostra aleatória de duas de suas pegadas e calculamos a distância de Mahalanobis entre a média dessas pegadas e a pegada humana média geral. Para as trilhas de Laetoli, as amostras do local A e do local S incluíram apenas duas trilhas, então essas foram simplesmente calculadas e a distância de Mahalanobis foi calculada entre cada trilha média e a pegada humana média. Para Laetoli site G, todas as combinações possíveis de duas pistas (dez) foram retiradas da amostra descrita acima, e a distância de Mahalanobis foi calculada entre a pista média de cada combinação e a média humana. Em todos os casos, calculamos distâncias multivariadas usando a matriz de covariância humana entre sujeitos (isto é, tratando o chimpanzé e as trilhas fósseis como se viessem de diferentes sujeitos humanos). Todas as análises descritas acima, e o histograma exibindo distâncias multivariadas (Fig. 2 ), foram gerados usando R (v. 3.6.1), com scripts personalizados e funções dos pacotes dplyr 55 , ggplot2 54 e reshape2 56 .
Fotogrametria
Embora as impressões das pegadas bípedes do site A existissem em um ponto, todas as nossas tentativas de localizá-las (consulte Agradecimentos para uma lista completa) foram malsucedidas. Antes de nosso trabalho de campo em Laetoli em 2019, modelamos a trilha original fotogrametricamente usando a fotografia existente no local. A fotografia original do local A de Laetoli foi tirada por JR. Obtivemos suas fotos da trilha A por meio da Science Photo Library. Todas as fotografias foram tiradas com uma Nikon F2 em filme slide 35 mm Kodachrome. Digitalizações digitais desses slides foram usadas para produzir um modelo 3D das pegadas de Laetoli A. Infelizmente, como as imagens foram tiradas em 1977, elas não foram gravadas com o processamento fotogramétrico moderno em mente. Vários recursos das imagens digitalizadas limitam a construção bem-sucedida e precisa de um modelo 3D. Primeiro, existem apenas quatro imagens das pegadas. Uma dessas imagens tem configurações de exposição visivelmente diferentes que causaram problemas de alinhamento significativos durante o processamento e, portanto, foi excluída. Todas as imagens foram tiradas em ângulos oblíquos, a partir de uma faixa relativamente estreita de posições da câmera. Uma linha amarela definindo a grade do local fica sobre uma das pegadas, obscurecendo parte dela e lançando uma sombra. As imagens foram todas tiradas relativamente no início do dia, portanto, há sombras dentro de cada pegada que criam contrastes fortes. As lâminas foram digitalizadas a 4.000 dpi, mas não foram digitalizadas com equipamento especializado para garantir a precisão geométrica, e isso potencialmente introduziu mais fontes de distorção.
However, despite the limitations of the images, it was possible to extract 3D data for the Laetoli A footprints. All processing was done in Agisoft Photoscan Pro (v. 1.7.1). The standard processing steps (align photos, build dense cloud, build mesh, build texture) were run to produce a 3D model, though the process had to be done iteratively to remove noise, ensure accurate alignment of the photos, scale the model appropriately using published measurements, add manual tie points, and refine the model. A DEM (digital elevation model) and orthophotograph were exported for further visualization and analysis in ArcGIS (v. 10.6.1). The 3D model was also exported to Autodesk Meshmixer (v. 3.5.474) to create a ‘watertight’ 3D volume that could be 3D printed for further visualization (1977 model is hosted on Morphosource, ID: 000390119). Photogrammetric reconstruction was validated using published measurements of the footprints. It is important to note however, that there were no published measurements for the depths of the footprints and that the internal anatomy of this reconstruction is potentially misleading because of the incomplete excavation of the footprints2 , 8 , 12 .
A second, more accurate reconstruction was done using photogrammetry from the re-excavated site A bipedal trackway using 57 images taken in June 2019. The images were captured in a systematic manner using a Nikon D7000 camera and Nikon DX AF-S Nikkor 18–105 mm lens. All photos were taken by hand, from an eye level, while walking a series of transects, across the area of interest. Spacing between shots was kept low to ensure a minimum of approximately 65% overlap between adjacent images. All processing was done using Agisoft Photoscan Pro/Metashape Pro (v. 1.7.1). Standard processing steps (for example, as described15,51) were taken to create a 3D model of the A trail. This included photo alignment, manual editing of the sparse cloud to remove points with high ‘reprojection uncertainty’, building a dense cloud, building a mesh, refining the mesh, then building a texture. During processing, images were checked for sharpness using the ‘image quality’ tool and any images with significantly lower quality were removed. The model was scaled to the real-world using scale bars placed across the region of interest. Finally, an orthophotograph as well as a DEM (digital elevation model) were exported as geotiffs into ArcGIS in an arbitrary local coordinate system for further analysis (2019 model hosted on Morphosource, ID: 000390114).
To generate contour maps, two approaches were used. First, starting with the raw stereolithography scans (.stl file format), Ultimaker Cura software (v.4.8.0) was used to rotate the raw scans and align them with x and y axes. This was a manual process. These rotations were exported to binary-format .stl files. The rotated files were then run through an R script using R version 4.0.3. The R script uses the tidyverse and rgl libraries to load the .stl files into R-friendly dataframes and plot them as contours using ggplot’s geom_contour function. The script is available through GitHub.
Usando uma segunda abordagem, os arquivos .stl foram trazidos para o Cloud Compare (v. 2.11.3) para verificar a orientação do modelo. Se necessário, os modelos eram reorientados para permitir que a superfície do solo local ficasse nivelada usando a ferramenta "nivelar" e, em seguida, os arquivos eram exportados. O modelo corretamente orientado foi importado para o SAGA GIS usando a ferramenta de importação de arquivo de litografia estéreo (STL). Esta ferramenta converte o .stl diretamente em um raster DEM. Os rasters foram verificados no SAGA e um sombreamento gerado com a função de sombreamento analítico usando a configuração da posição do sol padrão de 315 ° azimute e 45 ° de altura. O DEM e a sombra foram então exportados como geotiffs. Esses geotiffs foram importados para ArcGIS para visualização. O DEM foi colorido usando uma rampa de cor vermelho-azul para indicar a profundidade relativa e isso foi colocado em camadas sobre o raster de sombra usando o método de fusão NAGI 57 (dados estendidos Fig. 8 ). A comparação de nuvem foi usada para quantificar as alterações erosivas nas pegadas do local A de 1977 a 2019.
Digitalização de superfície 3-D
Digitalizações tridimensionais de superfície de Laetoli A e moldes de gesso de impressões de ursos foram coletados usando um Creaform Go! Scan 50.
Resumo do relatório
Mais informações sobre o desenho da pesquisa estão disponíveis no Nature Research Reporting Summary, vinculado a este artigo.
Disponibilidade de dados
Dados publicados anteriormente foram obtidos de refs. 4 , 19 , 20 , 21 , 28 , 32 , 42 . Todos os outros dados que apóiam as conclusões deste estudo estão disponíveis no documento e em seus arquivos de informações suplementares . A reconstrução fotogramétrica da trilha de Laetoli A com base em três fotografias originais da expedição de 1977 está disponível no Morphosource ( https://www.morphosource.org ; ID: 000390119). A reconstrução fotogramétrica da trilha de Laetoli A usando 57 fotografias tiradas do local de Laetoli A re-escavado em 2019 está disponível no Morphosource (ID: 000390114). Os dados de origem são fornecidos com este documento.
Referências
- 1.
Leakey, M. Pegadas do Plioceno em Laetoli, norte da Tanzânia. Antiquity 52 , 133 (1978).
- 2.
Leakey, MD & Hay, RL Pegadas do plioceno nos leitos de Laetoli em Laetoli, norte da Tanzânia. Nature 278 , 317-323 (1979).
- 3.
Leakey, MD & Harris, JM (eds) Laetoli, a Pliocene Site in Northern Tanzania (Clarendon, 1987).
- 4.
Tuttle, RH em Laetoli: A Pliocene Site in Northern Tanzania (eds Leakey, MD & Harris, JM) 503-523 (Clarendon, 1987).
- 5.
Day, M. H. in Origine(s) de la Bipedie Chez les Hominides (eds Coppens Y. & Senut B.) 199–213 (Editions du CNRS, 1991).
- 6.
Leakey, M. Disclosing the Past: An Autobiography (Doubleday, 1984).
- 7.
Leakey, M. D. Footprints in the ashes of time. Nat. Geo. 155, 446–457 (1979).
- 8.
White, T. D. & Suwa, G. Hominid footprints at Laetoli: facts and interpretations. Am. J. Phys. Anthropol. 72, 485–514 (1987).
- 9.
Tuttle, R. H. Bear facts and Laetoli impressions. Am. J. Phys. Anthropol. 63, 230–230 (1984).
- 10.
Tuttle, R. H. in Hominid Evolution: Past, Present, and Future 129-133 (Alan R. Liss, 1985).
- 11.
Tuttle, R. H. The pitted pattern of Laetoli feet. J. Nat. Hist. 99, 60–65 (1990).
- 12.
Tuttle, R., Webb, D. & Tuttle, N. in Origine(s) de la Bipedie Chez les Hominides (eds Coppens Y. & Senut B.) 187–198 (Editions du CNRS, 1991).
- 13.
Cohen, A., Lockley, M., Halfpenny, J. & Michel, AE Taphonomy moderno da trilha de vertebrados no Lago Manyara, Tanzânia. Palaios 6 , 371-389 (1991).
- 14.
Roach, NT et al. As pegadas do Pleistoceno mostram o uso intensivo de habitats nas margens do lago por Homo erectus grupos de . Sci Rep. 6 , 26374 (2016).
- 15.
Zimmer, B. et al. Usando fotogrametria diferencial de estrutura de movimento para quantificar a erosão no local da pegada de Engare Sero, na Tanzânia. Quat. Sci. Rev. 198 , 226–241 (2018).
- 16.
Su, DF & Harrison, T. Implicações ecológicas da raridade relativa de hominídeos fósseis em Laetoli. J. Hum. Evol. 55 , 672–681 (2008).
- 17.
Harrison, T. em Paleontologia e Geologia de Laetoli: Evolução Humana no Contexto 1-14 (Springer, 2011).
- 18.
Roach, NT et al. Comunidades animais do Pleistoceno de uma pastagem nas margens de um lago de 1,5 milhão de anos e sua relação com a Homo erectus paleoecologia do . J. Hum. Evol. 122 , 70–83 (2018).
- 19.
DeSilva, JM & Gill, SV Breve comunicação: uma fratura no meio do tarso (mediopé) no pé humano. Sou. J. Phys. Anthropol. 151 , 495–499 (2013).
- 20.
Hatala, KG, Dingwall, HL, Wunderlich, RE e Richmond, BG A relação entre a pressão plantar e a forma da pegada. J. Hum. Evol. 65 , 21–28 (2013).
- 21.
Hatala, KG, Wunderlich, RE, Dingwall, HL & Richmond, BG Interpretando a biomecânica locomotora a partir da morfologia das pegadas humanas. J. Hum. Evol. 90 , 38–48 (2016).
- 22.
Liutkus-Pierce, CM et al. Idade radioisotópica, formação e preservação de pegadas humanas do Pleistoceno Superior em Engare Sero, Tanzânia. Palaeogeogr. Palaeoclimatol. Palaeoecol. 463 , 68–82 (2016).
- 23.
Hatala, K. G. et al. Snapshots of human anatomy, locomotion, and behavior from Late Pleistocene footprints at Engare Sero, Tanzania. Sci. Rep. 10, 7740 (2020).
- 24.
Cham, R. & Redfern, M. S. Changes in gait when anticipating slippery floors. Gait Posture 15, 159–171 (2002).
- 25.
Fukushima, T. Angle and distance between steps. J. Bone. Jt. Surg. 37, 1311 (1955).
- 26.
Murray, M. P., Drought, A. B. & Kory, R. C. Walking patterns of normal men. J. Bone Jt. Surg. 46, 335–360 (1964).
- 27.
Maki, B. E. & McIlroy, W. E. Control of rapid limb movements for balance recovery: age-related changes and implications for fall prevention. Age Ageing 35, ii12–ii18 (2006).
- 28.
Thompson, N. E., O’Neill, M. C., Holowka, N. B. & Demes, B. Step width and frontal plane trunk motion in bipedal chimpanzee and human walking. J. Hum. Evol. 125, 27–37 (2018).
- 29.
O’Neill, M. C., Demes, B., Thompson, N. E. & Umberger, B. R. Three-dimensional kinematics and the origin of the hominin walking stride. J. R. Soc. Interface 15, 20180205 (2018).
- 30.
Tuttle, R. H. Evolution of hominid bipedalism and prehensile capabilities. Philos. Trans. R. Soc. Lond. B. 292, 89–94 (1981).
- 31.
Harcourt-Smith, WE Será que o Australopithecus afarensis fez a trilha da pegada de Laetoli? - Novos insights sobre um problema antigo. Sou. J. Phys. Anthropol. S40 , 116 (2005).
- 32.
Pelissero, AJ A Comparative Analysis of Newly Discovered Pliocene Hominin Footprints from Laetoli, Tanzania (Univ. Colorado at Denver, 2017).
- 33.
Haile-Selassie, Y. et al. Um novo pé hominídeo da Etiópia mostra várias adaptações bípedes do Plioceno. Nature 483 , 565-569 (2012).
- 34.
Dingwall, HL, Hatala, KG, Wunderlich, RE & Richmond, BG Hominin estatura, massa corporal e estimativas de velocidade de caminhada com base em pegadas fósseis de 1,5 milhão de anos em Ileret, Quênia. J. Hum. Evol. 64 , 556–568 (2013).
- 35.
Masao, FT et al. Novas pegadas de Laetoli (Tanzânia) fornecem evidências de uma variação acentuada no tamanho do corpo nos primeiros hominídeos. eLife 5 , e19568 (2016).
- 36.
DeSilva, JM, Gill, CM, Prang, TC, Bredella, MA & Alemseged, Z. Um pé quase completo de Dikika, Etiópia e suas implicações para a ontogenia e função do Australopithecus afarensis . Sci. Adv. 4 , eaar7723 (2018).
- 37.
Raichlen, DA & Gordon, AD A interpretação das pegadas do Site S confirma a biomecânica bípede semelhante à humana em hominíneos Laetoli. J. Hum. Evol. 107 , 134–138 (2017).
- 38.
Bennett, MR & Morse, SA em Human Footprints: Fossilized Locomotion? 101–136 (Springer, 2014).
- 39.
Crompton, RH et al. Função externa do pé semelhante à humana e marcha totalmente ereta, confirmada nas pegadas de hominíneos Laetoli de 3,66 milhões de anos por estatísticas topográficas, formação experimental de pegadas e simulação de computador. JR Soc. Interface 9 , 707–719 (2012).
- 40.
Meldrum, DJ, Lockley, MG, Lucas, SG & Musiba, C. Ichnotaxonomy of the Laetoli trackways: the first hominin footprints. J. Afr. Terra. Sci. 60 , 1–12 (2011).
- 41.
Ward, C. V., Kimbel, W. H. & Johanson, D. C. Complete fourth metatarsal and arches in the foot of Australopithecus afarensis. Science 331, 750–753 (2011).
- 42.
Hatala, K. G., Demes, B. & Richmond, B. G. Laetoli footprints reveal bipedal gait biomechanics different from those of modern humans and chimpanzees. Proc. R. Soc. Lond. B 283, 20160235 (2016).
- 43.
Leakey, M. G. et al. New hominin genus from eastern Africa shows diverse middle Pliocene lineages. Nature 410, 433–440 (2001).
- 44.
Haile-Selassie, Y., et al New species from Ethiopia further expands Middle Pliocene hominin diversity. Nature 521, 483–488 (2015).
- 45.
Musiba, C., et al Newly discovered hominin remains from upper Laetoli beds in northern Tanzania. Abstracts of the PaleoAnthropology Society, 2010 Meetings. PaleoAnthropology 2010, A0001–A0040 (2010).
- 46.
Pelissero, A. J. et al. Photogrammetry imaging: a fresh look at the Laetoli hominin footprints in relation to recent discoveries. Am. J. Phys. Anthropol. 162, 312 (2017).
- 47.
Pelissero, A., Musiba, C., Magori, C., Mwankunda, J. & Bunn, H. Rethinking the Laetoli hominin trackmakers in light of new prints at Site S. In Paleoanthropology Society Meeting, Albuquerque, NM (2019).
- 48.
DeSilva, J., McNutt, E., Benoit, J. & Zipfel, B. One small step: a review of Plio‐Pleistocene hominin foot evolution. Am. J. Phys. Anthropol. 168, 63–140 (2019).
- 49.
Musiba, C. M., Mabula, A., Selvaggio, M. & Magori, C. C. Pliocene animal trackways at Laetoli: research and conservation potential. Ichnos 15, 166–178 (2008).
- 50.
Bennett, MR et al. Morfologia do pé de hominídeo com base em pegadas de 1,5 milhão de anos de Ileret, Quênia. Science 323 , 1197-1201 (2009).
- 51.
Abramoff, MD, Magalhães, PJ & Ram, SJ Processamento de imagens com ImageJ. Biophotonics Int. 11 , 36–42 (2004).
- 52.
D'Aout, K., Meert, L., Van Gheluwe, B., De Clercq, D. & Aerts, P. Pegadas geradas experimentalmente na areia: análise e consequências para a interpretação de pegadas fósseis e forenses. Sou. J. Phys. Anthropol. 141 , 515–525 (2010).
- 53.
Budka, M. et al. DigTrace Pro v.1.8.1 (Bournemouth University, 2016).
- 54.
Wickham, H. ggplot2: Elegant Graphics for Data Analysis (Springer, 2016).
- 55.
Wickham, H., François, R., Henry, L. & Müller, K. dplyr: a grammar of data manipulation. Versão 0.8.3 (2019).
- 56.
Wickham, H. Reshape2: reformular dados de forma flexível: uma reinicialização do pacote Reshape. Versão 1.4.4 (CRAN, 2019).
- 57.
Nagi, R. Manutenção de detalhes e definição de cor ao integrar rasters de cores e tons de cinza usando o método de fusão de intensidade ou de escala de cinza sem alteração (NAGI). Em Proc. 2012 AutoCarto (2012).
Reconhecimentos
Agradecemos a R. Tuttle por sua orientação e inspiração; M. Larkin por sua ajuda na coleta de dados sobre pegadas de ursos; R. Leakey, B. Wood, H. de Lumley, T. White, R. Crompton, R. Clarke, W. Harcourt-Smith, C. Stringer, H. Bonney and the London Natural History Museum, e J. Kibii e ao Museu Nacional do Quênia pela ajuda enquanto tentávamos rastrear fotografias e moldes das pegadas de Laetoli A; J. Sappington para suporte fotográfico; e V. Rossi e A. Metallo pela ajuda com a visualização 3D; ao Diretor Geral da Comissão de Ciência e Tecnologia da Tanzânia (COSTECH) por conceder à nossa equipe as Permissões de Pesquisa COSTECH (2019-370-NA-2019-2016); F. Manongi, J. Mwankunda e M. Mwambungu por nos permitirem acampar e trabalhar em Laetoli. O financiamento para esta pesquisa foi fornecido pela National Geographic (8748-10), Leakey Foundation (71483-001), National Science Foundation (BCS-1128170, DGE-080163, BCE-1730822, GRFP (No. 1840344), o Smithsonian Institution's Programa Origens Humanas, Fundação Terra em Evolução, Clube dos Exploradores e Claire Garber Goodman Fund-Dartmouth College.
Informação sobre o autor
Afiliações
Contribuições
EJM contribuiu com a geração e desenho do estudo, coleta de dados sobre ursos e chimpanzés e análises, e escreveu o manuscrito. KGH contribuiu para a coleta de dados sobre humanos e chimpanzés e análises, e ajudou com o manuscrito. CM contribuiu para a coleta de dados durante o trabalho de campo em Laetoli e análises. JC e ACH contribuíram com a análise da fotogrametria. JA, ACH e SG desenvolveram os mapas de contorno das pegadas. ASD, KF, LDF, JG, EG, SK, BM, AP, SR, RT e CMM contribuíram para a coleta de dados durante o trabalho de campo em Laetoli. NJD contribuiu com o desenho do estudo e auxiliou na elaboração do manuscrito. SVG, JMD, KGH e EMW-H. contribuiu com a coleta de dados em humanos. JMD também contribuiu com a coleta de dados durante o trabalho de campo em Laetoli, desenho do estudo e auxiliou na elaboração do manuscrito. CJ contribuiu com a coleta de dados sobre ursos e análises. BK e PK contribuíram para o acesso aos ursos selvagens e coleta de dados. CL-P. e BZ contribuiu para o acesso e coleta de dados nas pegadas do Engare Sero. EK contribuiu com análises das pegadas do Engare Sero. JR contribuiu com a fotografia original do Site Laetoli. ANET contribuiu para a coleta de dados sobre chimpanzés. Todos os autores contribuíram com comentários editoriais para o manuscrito.
autor correspondente
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Extended data figures and tables
Extended Data Fig. 1 Laetoli print A3 and erosion.
Photographs were taken by J. Reader (left: 1977) and J. DeSilva (right: 2019) at similar overhead angles and times of day (see similarities in shadows cast across the print). Notice the removed matrix infill in the hallucial impression and the presence of the previously unseen second digit impression in 2019 image. Below: comparisons of the photogrammetric meshes of a pothole and footprints A2 and A3 using Cloud Compare15. The impact of erosion on the morphology of the footprints was assessed by quantifying changes to an oval depression located west of A3. Notice that significant changes occurred around the rim of the depression, as would be expected through erosion, that are absent around the rims of the A2 and A3. Instead, significant differences between A2 and A3 are located internally and are a result of a more thorough excavation of the prints.
Extended Data Fig. 2 Details of the best preserved Laetoli A hominin footprint (A3).
Left image shows original 3D scan. Right image highlights the proposed impressions for the toes (blue circles) and matrix infill (green), as well as the potential evidence for midfoot mobility (red).
Extended Data Fig. 3 Incidence of bipedalism in Ursus americanus and Examples of kinematic data collection.
(topo) Gráficos de pizza mostrando a frequência de comportamentos bípedes em selvagem Ursus americanus . O gráfico azul representa o tempo gasto de forma independente bípede (locomotora ou postural) de um total de 50,9 h em que o comportamento do urso foi observado. O gráfico laranja representa a divisão do tempo gasto independentemente bípede em seus componentes postural e locomotor (ou seja, os ursos deram uma ou mais etapas). O gráfico verde representa a frequência de ocorrências em que os ursos caminharam 4 ou mais passos refletindo uma circunstância semelhante à trilha A. de Laetoli (embaixo, à esquerda) A fêmea juvenil caminha bípede, sem ajuda, pela trilha de lama. (canto inferior direito) Exemplo de pegada esquerda de um dos machos jovens.
Dados estendidos Fig. 4 Comparações de pé e marcha em todas as espécies comparativas.
(a) Largura do antepé até o comprimento do passo; (b) largura do salto ao comprimento do passo; (c) largura do calcanhar à largura do antepé; e (d) largura do passo ao comprimento do passo. Observe que para as proporções dos pés (c) e largura da passada (d), Laetoli A é diferente das pegadas produzidas por ursos. Em (d) o valor negativo em Laetoli A representa o fato de que a pista está demonstrando passos cruzados. (ac) Boxplot representa mediana (linha central), quartis superior e inferior (limites de caixa), intervalo (bigodes) e outliers (pontos) e tamanhos de amostra de pegadas individuais para cada espécie são indicados nos painéis de figuras. (ad) Em todas as parcelas, n = 1 para todos os rastros de Laetoli, e os chimpanzés estavam viajando bípede. As fontes de dados correspondem à Tabela dados estendidos 1 de .
Dados estendidos Fig. 5 Evidência que desafia a hipótese ursid.
(a) Razão entre as dimensões de impressão do dedo do pé nas pegadas das espécies comparativas e Laetoli A3. Valores em azul = relação entre a largura do 2º dígito em relação ao hálux. Valores em laranja = razão entre a largura do 4º dígito em relação ao 5º em Ursus americanus . (Inserir imagem) Comparação entre a varredura 3D da pegada ursídica direita (azul) e A3 (laranja). Observe o grande tamanho da 5º impressão do dígito nos ursídeos, mas a diferença geral de formato entre a faixa ursid e A3. (b) Razão entre a largura da passada e o comprimento do passo nas diferentes espécies, incluindo as trilhas bípedes de Laetoli. As definições de boxplot são as da Fig. dados estendidos 4 de e os tamanhos de amostra de pegada individual são indicados nos painéis de figuras. Em 5a, os chimpanzés viajavam quadrúpede; em 5b, os chimpanzés se moviam bípede. As fontes de dados correspondem à Tabela dados estendidos 1 de .
Extensão Fig dados. 6 Boxplots mostrando a relação de divergência entre o primeiro dígito e 2 nd dígitos entre as espécies comparativos e duas pegadas Laetoli.
A razão de divergência foi medida dividindo a distância linear entre o ponto médio dos dígitos um e dois pelo comprimento do pé. Os chimpanzés viajavam quadrúpede e os dados foram obtidos a partir de impressões de pressão plantar. As definições de boxplot são as da Fig. dados estendidos 4 de e os tamanhos de amostra de pegada individual são indicados no painel da figura. As fontes de dados correspondem à Tabela dados estendidos 1 de . Tentamos medir a divergência alucial seguindo Bennett e cols 50 . . mas foram bloqueados pela matriz obscurecendo a região mais profunda da planta do pé.
Dados estendidos Fig. 7 Histograma de distâncias de Mahalanobis entre a pegada humana sem ferradura média e as pegadas humanas sem ferragem reamostradas (cinza) e pegadas de chimpanzé (amarelo).
Como na Figura 3c, as linhas tracejadas em azul, laranja e verde representam amostras de Laetoli G1, S1 e A, respectivamente. Apenas Laetoli A é rotulado, para maior clareza. Tamanhos de amostra para essas amostras como na Fig. 2c também. As linhas pretas pontilhadas e tracejadas foram adicionadas para representar a média das pegadas cruzadas produzidas por 10 humanos adultos habitualmente calçados. Todos se enquadram na distribuição de pegadas humanas descalças (as probabilidades de trilhas de amostragem como essas variam de 0,20 a 0,94) e a uma grande distância da amostra Laetoli A (verde). As pegadas cruzadas humanas tendem a ser ligeiramente mais próximas da média humana do que as amostras de Laetoli S1 e G1, mas sua distribuição se sobrepõe à amostra de Laetoli S1 e a algumas das amostras de Laetoli G1. As pegadas cruzadas caíram, em média, a uma distância de Mahalanobis de 27,2 mais longe do meio humano do que sua contraparte “normal” de caminhada.
Dados estendidos Fig. 8 Varreduras 3D das pegadas de Laetoli A e seus contornos.
Painel esquerdo: varredura completa da trilha Laetoli A com A1 na parte inferior e A5 na parte superior. Lado direito: Imagens ampliadas de pegadas individuais e suas imagens de contorno correspondentes.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.