Evolução inicial do tamanho corporal pequeno no Homo floresiensis
Comunicações da Natureza 15 , Número do artigo: 6381 ( 2024 )
Resumo
Descobertas recentes de Homo floresiensis e H. luzonensis levantam questões sobre como ocorreu a redução extrema do tamanho do corpo em algumas espécies extintas de Homo em ambientes insulares. Investigações anteriores em Mata Menge, Ilha das Flores, Indonésia, sugeriram que os ancestrais do início do Pleistoceno Médio de H. floresiensis tinham mandíbulas e dentes ainda menores. Aqui, relatamos fósseis de hominídeos adicionais dos mesmos depósitos em Mata Menge. Estima-se que um úmero adulto seja 9 a 16% mais curto e mais fino do que o espécime tipo de H. floresiensis datado de ~60.000 anos atrás, e é menor do que qualquer outro úmero de hominídeo adulto do Plio-Pleistoceno relatado até agora. Os dentes recém-recuperados são ambos excepcionalmente pequenos; um deles tem semelhanças morfológicas mais próximas ao H. erectus javanês antigo . A linhagem H. floresiensis provavelmente evoluiu do antigo H. erectus asiático e foi uma linhagem duradoura em Flores, com tamanho corporal marcadamente diminuto, há pelo menos ~700.000 anos.
Conteúdo semelhante sendo visualizado por outros


Introdução
A Bacia de So'a, no centro de Flores, Indonésia, é uma região-chave para elucidar a origem e a evolução do H. floresiensis , uma espécie diminuta de hominídeo conhecida do Pleistoceno Superior em Liang Bua, uma caverna de calcário no oeste de Flores 1 , 2 . Assim como outro Homo de corpo pequeno descoberto em Luzon 3 , a história evolutiva desta espécie insular de hominídeo tem sido objeto de debate prolongado 4 . Estudos de campo anteriores da sequência do Pleistoceno Inferior ao Médio (Calábrico-Chibaniano) da Bacia de So'a recuperaram restos fósseis de fauna endêmica ( Stegodon anão , lagarto-monitor de Komodo, rato gigante, pássaros, crocodilos e tartarugas) 5 , 6 , artefatos de pedra tecnologicamente simples (o mais antigo dos quais data de pelo menos 1,02 ± 0,02 milhões de anos atrás [Ma]) 7 , 8 e, mais importante, uma mandíbula fragmentária e seis dentes isolados de um hominídeo de pequeno porte 9 . Esses fósseis de hominídeos foram escavados de uma camada de arenito de origem fluvial (Camada II) do intervalo superior portador de fósseis no sítio Mata Menge, que é datado entre 0,65 e 0,773 Ma 5 , 6 . Esses fósseis exibem afinidades morfológicas gerais com a série tipo de H. floresiensis de Liang Bua (0,1–0,06 Ma) 10 e com o antigo H. erectus de Java (1,1–0,8 Ma) 11 , mas não apresentam as especializações molares únicas que caracterizam o primeiro e eram substancialmente menores que o último 9 .
No geral, os fósseis de Mata Menge sugerem que eles representam um segmento ancestral da linhagem Liang Bua H. floresiensis , e que os hominídeos de Flores são descendentes anões do H. erectus asiático primitivo de corpo grande 9 . Algumas análises cladísticas/filogenéticas, no entanto, apoiam uma ligação evolutiva direta entre H. floresiensis e Homo basais de corpo menor, como H. habilis ou mesmo Australopithecus 12 , 13 , 14 . É importante resolver essa controvérsia para elucidar o padrão e o momento da evolução do tamanho corporal nos hominídeos de Flores.
Notavelmente, a mandíbula e os dentes de Mata Menge são ligeiramente menores do que os espécimes-tipo de H. floresiensis de Liang Bua. Isso sugere que uma redução dentognática drástica ocorreu em Flores no início do Pleistoceno Médio, mais de 600.000 anos antes da evidência fóssil mais antiga de H. floresiensis em Liang Bua. Até agora, no entanto, a falta de elementos pós-cranianos na assembleia de Mata Menge havia limitado nossa compreensão da evolução do tamanho do corpo em Flores.
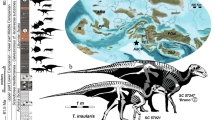
Neste artigo, relatamos a descoberta e a morfologia de um fóssil pós-craniano de hominídeo de Mata Menge, um eixo umeral distal extremamente pequeno (SOA-MM9) (Fig. 1 ). Este espécime e dois dentes de tamanho pequeno (SOA-MM10 e SOA-MM11) foram recuperados como adições ao conjunto de hominídeos existente da Camada II neste local (Tabela 1 ). Nosso exame histomórfico confirma o status adulto do úmero. Também mostramos que a morfologia do eixo é mais semelhante ao Homo de corpo pequeno (por exemplo, LB1 e H. naledi ) do que ao Australopithecus (por exemplo, AL 288-1), e uma coroa molar (SOA-MM11) apresenta semelhanças de forma mais próximas ao H. erectus javanês antigo do que ao Homo africano antigo . A amostra aumentada de fóssil de Mata Menge reforça sua classificação como um representante antigo de H. floresiensis , que provavelmente sofreu uma redução drástica no tamanho do corpo em relação ao H. erectus asiático de grande porte em algum momento entre ~1,0 e 0,7 Ma.

a – f Úmero SOA-MM9 em vistas anterior, lateral, posterior, medial, proximal e distal. g Seção de micro-TC de SOA-MM9 na extremidade distal indicada em ( c ). h e i Úmero LB1 em vistas anterior e medial. Observe a área oca na superfície posterior da extremidade distal (triângulos preenchidos em f e g ), que serve como um marco osteométrico (ponto hOF). Abreviações: ant = anterior, post = posterior, lat = lateral, med = medial, DT = tuberocidade deltoide, NF = forame nutriente, HS = localização para corte histológico. j SOA-MM10 direita d c . Da esquerda para a direita, vistas oclusal, labial e lingual. k SOA-MM11 esquerda M 3 . Da esquerda para a direita, vistas oclusal, bucal e lingual.
Resultados
Contexto e idade geológica
Todos os fósseis de hominídeos se originaram do topo de uma camada de arenito seixo endurecido em forma de fita (Camada II), que foi depositada em um pequeno canal de riacho em um leque aluvial vulcanoclástico há 5 ~ 0,7 Ma. Esta estimativa de idade é baseada na identificação do limite Brunhes-Matuyama 15 datado em 0,773 Ma por medições paleomagnéticas combinadas com uma série de datações de traços de fissão em zircões em estratos tufáceos estratigraficamente 16,5 m abaixo da Camada II 5 , 6 . Uma idade mínima de 0,65 ± 0,02 Ma para a Camada II é fornecida por uma data de 40 Ar/ 39 Ar em cristais de hornblenda simples de uma tefra de queda de ar (PGT-2) ocorrendo estratigraficamente a 14 m acima da Camada II.
A camada II, com espessura máxima de 50 cm, recobre um paleossolo avermelhado (camada III) com um contato erosivo ondulado. Uma série de camadas maciças de fluxo de lama rico em argila tuffácea (camadas Ia-f) selou as camadas II e III posteriormente (Fig. 2 ; Nota complementar 1 ). O fragmento de úmero SOA-MM9 foi recuperado em vários pedaços em uma semana após a abertura da escavação 32 A em 2013, mas foi reconhecido como tal apenas em 2015 após reconstrução em laboratório. O espécime foi danificado no processo de escavação do arenito extremamente compacto da camada II. Um canino decíduo maxilar (d c : SOA-MM10) foi escavado em 2015 a ~5 cm abaixo do limite entre as Camadas I e II, enquanto um terceiro molar mandibular (M 3 : SOA-MM11) foi escavado em 2016 a ~15 cm abaixo do topo da Camada II. Todos os fósseis de hominídeos estão concentrados na parte superior da Camada II, enquanto fósseis de outras faunas tendem a ser distribuídos de forma mais uniforme nesta unidade. Há evidências de transporte fluvial dos fósseis antes do enterramento, com muitos (mas não todos) espécimes fraturados (além de danos de escavação), desgastados e/ou arredondados até certo ponto 16 . No entanto, os três fósseis de hominídeos descritos aqui mostram evidências mínimas/nenhuma de abrasão. Observações tafonômicas e sedimentológicas sugerem que os fósseis de hominídeos foram depositados durante um regime de fluxo de energia moderada a baixa no riacho, após um período relativamente breve na superfície durante o qual os restos foram desarticulados (Nota Suplementar 1 ). Logo após a incorporação dos fósseis no leito do riacho, todo o vale do riacho foi preenchido com uma sequência de fluxos de lama de 6,5 m de espessura. Escavações de campo subsequentes em 2017–2019 e 2023 não produziram mais fósseis de hominídeos deste local.

a Mapa de Elevação Digital (DEM) de Flores mostrando a localização da Bacia de So'a e da caverna Liang Bua. b DEM da Bacia de So'a mostrando a localização das escavações de Mata Menge. c Foto do baulk oeste do setor de escavação sul (Setor 32C) no intervalo superior com fósseis em Mata Menge. A Camada III é um paleossolo arenoso avermelhado, coberto com um contato erosivo por uma camada fluvial arenosa (Camada II). As Camadas II e III são cobertas por uma série de unidades de fluxo de lama de cinzas ricas em argila (Camadas Ia-f). O canino decíduo SOA-MM10 foi recuperado a 5 cm abaixo do topo da Camada II (indicado com o retângulo azul tracejado; o retângulo azul é ampliado em e). Observe também os grandes ossos de Stegodon repousando no topo da Camada II e cobertos pelas unidades de fluxo de lama. d Foto do canto noroeste da escavação no setor 32 A, tirada em 7 de novembro de 2013, quatro semanas após a recuperação do fragmento de úmero de hominídeo SOA-MM9. O fóssil foi escavado do topo da Camada II, com a posição aproximada indicada com o oval azul tracejado. e Detalhe do contato entre a Camada I e a Camada II no local do canino decíduo SOA-MM10. f SOA-MM10 ainda parcialmente embutido no arenito da Camada II. g Grade de escavação de Mata Menge (UTM Zona 51 L) mostrando os quadrantes de 1 × 1 m escavados no final da temporada de campo de 2016 em cinza. As posições dos fósseis de hominídeos descritos neste artigo são indicadas com pontos verdes, aquelas descritas anteriormente 9 com pontos cinza. O sombreamento claro representa a vala em degraus escavada em 2010, que revelou pela primeira vez a presença do leito ósseo do intervalo superior contendo fósseis de Mata Menge. h SOA-MM11 cercado por sua matriz de arenito quando escavado em 2016. Os mapas ( a e b ) criados com GeoMapApp ( www.geomapapp.org ) / CC BY / CC BY (Ref. 67 )”.
Idade de desenvolvimento do úmero (SOA-MM9)
Este espécime é uma metade distal não distorcida do eixo umeral direito que mede 88 mm no comprimento máximo preservado (Fig. 1 ; Nota Suplementar 2 ). Apesar de seu pequeno tamanho, a histomorfologia do osso cortical de SOA-MM9 indica claramente seu status adulto. Examinamos seu estágio de desenvolvimento com base no aumento associado à idade de ósteons e estruturas relacionadas, um método amplamente utilizado para estimativas de idade de hominídeos existentes e fósseis 17 , 18 , 19 , 20 , 21 .
Cortes histológicos foram examinados para amostras corticais retiradas nas hastes médio-posteriores de SOA-MM9 ('HS' na Fig. 1 ) e de uma amostra humana moderna (N = 20, veja Dados Suplementares 1 ). Para permitir a variação regional na formação de ósteons dentro de cada área óssea 21 , os dados foram coletados de dois locais adicionais (próximos) na haste média para todos os espécimes humanos modernos ( H. sapiens ) (Fig. 3a ). Em dois parâmetros indicativos da maturidade óssea, Densidade Populacional de Ósteons (OPD) 21 e Índice do Canal de Havers (HCI), SOA-MM9 apresentou valores distintamente maiores (OPD = 16,5, HCI = 0,85) do que em qualquer um dos úmeros subadultos humanos modernos (0,0–8,9 e 0,0–0,63, respectivamente) (Fig. 3b , Dados Suplementares 1 ). Os valores para o úmero de Mata Menge também são maiores do que as médias de nossas amostras de adultos humanos modernos (13,6 e 0,78, respectivamente), indicando que o indivíduo SOA-MM9 estava bem dentro da idade adulta no momento da morte. Embora as superfícies corticais externas de SOA-MM9 exibam danos microscópicos que podem ter reduzido um dos osteons marginais à metade (~100 mícrons) (Fig. 3c ), tais alterações pós-deposicionais teriam impacto limitado em nossa estimativa de idade. Mesmo se assumirmos abrasão de superfície de 200 mícrons, o valor de OPD para SOA-MM9 cairia apenas ligeiramente para ~15,8. Além disso, a dominância de osteons secundários no córtex externo (Fig. 3c ) indica que o crescimento ósseo subperiosteal durante o período de crescimento já havia sido encerrado neste indivíduo 22 .

a Três locais em uma seção cortical do eixo médio-posterior do úmero usados para as análises histomorfológicas. 'Campo médio' é um círculo desenhado no terço externo da linha sagital média, e o 'campo lateral' e o 'campo medial' são círculos correspondentes a 3 mm de distância dali. b Valores histomorfométricos relacionados à idade para SOA-MM9 e humanos modernos. Os dados dos três locais são plotados para todos os espécimes humanos modernos, enquanto os dados para SOA-MM9 são apenas do campo lateral. As linhas pontilhadas são as médias da subamostra adulta humana moderna. Veja Dados Suplementares 1 para os dados originais. c Seção cortical do eixo médio posterior de SOA-MM9 observada por um microscópio de ultra-alta definição (VHX-7000, Keyence). A imagem original (esquerda) e a mesma imagem colorida para osteons secundários intactos (vermelho), osteons fragmentários (laranja e amarelo) e a superfície interna intacta (azul) (direita). Observe a dominância de osteons secundários ao redor da superfície externa (lado negativo das imagens). Apenas uma seção (mostrada aqui) foi produzida para minimizar os danos ao espécime original. d Crista supracondilar lateral de SOA-MM9 e sua seção de TC (as setas). Observe o desenvolvimento fraco, mas distinto, da crista como uma leve eversão.
Nenhuma evidência de patologia foi encontrada em SOA-MM9. O afinamento do osso cortical e o osso entrelaçado seriam patognômicos de alguns distúrbios metabólicos 17 , mas essas características não são evidentes em SOA-MM9. A espessura relativa do osso cortical de SOA-MM9 (0,07: a razão da espessura do osso cortical em relação à circunferência do eixo umeral) (Dados Suplementares 1 ) é quase idêntica à média para a amostra de adultos humanos modernos (0,069). Pacientes com osteogênese imperfeita, que pode levar à baixa estatura, exibem valores de OPD subnormais 19 , uma tendência que está em oposição à condição SOA-MM9. Além disso, o desenvolvimento fraco, mas distinto, da crista supracondilar lateral de SOA-MM9 (Fig. 3d ) indica desenvolvimento normal do músculo extensor radial longo do carpo.
Tamanho do úmero
Em todas as dimensões disponíveis de diâmetro/circunferência e comprimento do eixo, SOA-MM9 é menor que LB1 ( H. floresiensis ) e quaisquer outros indivíduos adultos de hominídeos fósseis de corpo pequeno ( Australopithecus e H. naledi : Dados Suplementares 2 ). Sua circunferência mínima (46 mm) é menor que UW 101-283 (47,5 mm), BOU-12/1 (52 mm) e os menores úmeros em nossa amostra humana moderna pré-histórica (46,5 mm, N = 1050, veja Dados Suplementares 2 ). O tamanho do centroide na seção transversal de nível ~19% também é o menor em comparação com quaisquer espécimes adultos amostrados de Australopithecus , Paranthropus e Homo , incluindo H. naledi e Liang Bua H. floresiensis (Fig. 4 , Dados Suplementares 3 ). O comprimento do eixo distal medido entre os pontos NF (forame nutriente) e hOF (margem superior da cavidade que leva à fossa do olécrano) do SOA-MM9 (58 mm) é nitidamente mais curto do que o de outros fósseis de hominídeos, incluindo LB1 (64 mm) (Dados Suplementares 2 ), embora a posição vertical do NF seja variável nos úmeros humanos 23 .
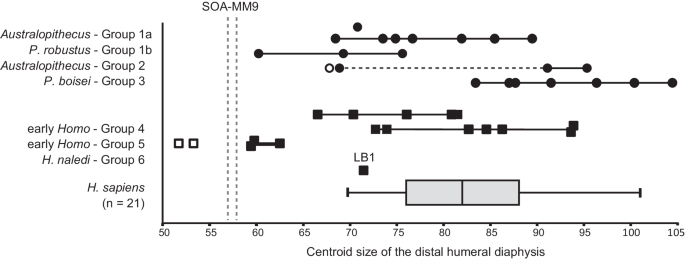
Símbolos: círculo = australopiteco; quadrado = Homo ; preto = adulto (possivelmente incluindo alguns adolescentes tardios); branco = subadulto. Os espécimes em cada grupo morfológico estão listados nos Dados Suplementares 3 junto com seus valores de tamanho. Um gráfico de caixa padrão (a mediana na linha horizontal e os 'bigodes' representando valores mínimos/máximos) é mostrado para uma amostra (N = 21) de humanos modernos. A linha tracejada transversal para o Grupo 2 conecta espécimes pertencentes a Au. sediba e Au. sp. indet . As linhas tracejadas verticais representam os menores (seção proximal) e maiores (seção distal) valores obtidos para SOA-MM9, que é menor do que todos os hominídeos fósseis adultos do Plio-Pleistoceno (incluindo LB1) e semelhante em tamanho aos espécimes de H. naledi . Note que o status totalmente adulto (fusão epifisária proximal ou marcas musculares desenvolvidas no eixo) não pode ser confirmado para alguns espécimes, como os menores indivíduos do Grupo 1b (SKX 10924) e Grupo 4 (SK 2598 e SK 24600). Veja Dados Suplementares 3 para outras notas.
A natureza fragmentária do SOA-MM9 impede uma reconstrução precisa de seu comprimento original, mas pode ser estimado da seguinte forma. Primeiro, a extremidade proximo-posterior preservada do SOA-MM9 está muito próxima do nível de 50% porque essa porção exibe o seguinte conjunto de características dos eixos médios umerais dos hominídeos: 1) O sulco radial (sulco espiral) está presente não na superfície lateral, mas no aspecto posterolateral, visto como uma área achatada nos cortes de TC nos. 1900 e 2000 ('RS' na Fig. 5 ); 2) na vista lateral, a margem anterior exibe uma leve concavidade ao redor do no. 1900 ('AM' na Fig. 5 , veja a imagem renderizada da superfície no lado esquerdo), indicando que essa parte, que está ~13 mm abaixo da extremidade proximo-posterior preservada, leva à tuberosidade deltoide proximalmente. A margem distal da tuberosidade deltoide está situada na superfície anterolateral no nível de 48,4% em nossa média amostral humana moderna (N = 366, intervalo: 43-53%), 51% em LB1 24 e 48% em KNM-WT 15000 25 ; 3) NF está presente 21 mm distal à extremidade proximo-posterior preservada de SOA-MM9. A distância projetada ao longo do eixo do nível de 50% e da margem inferior do NF é de 23 mm em LB1, 1 mm em KNM-WT15000, 15 mm em MH2 ( Au. sediba ) 26 e 21,2 mm em nossa média amostral humana moderna (N = 366, DP = 10,0 mm, intervalo = –2 a 59 mm). Cada um dos três caracteres acima mostra variação substancial, mas sua expressão simultânea no eixo proximal preservado sugere fortemente que a extremidade proximo-dorsal de SOA-MM9 estava muito próxima do nível original de 50%. Este posicionamento é consistente com outras morfologias de eixo exibidas por SOA-MM9 (Nota Suplementar 3 , Fig. 5 , Figs. Suplementares 1 e 2 e Dados Suplementares 4 ).

Esquerda: Imagens renderizadas de superfície. No sentido horário a partir do canto superior esquerdo: vistas anterior, posterior, lateral direita e lateral esquerda. Direita: Seções de TC e ossos corticais reconstruídos no nível do corte indicado pelos numerais (250 − 2183). Os cortes n.º 607, 573 e 639 são estimados em níveis de 19% (a melhor estimativa e intervalo provável: consulte “Métodos”). A espessura do corte desses cortes de TC é de 0,04 mm, de modo que a diferença de 100 corresponde a 4 mm. A reconstrução do osso cortical foi feita com referência aos ossos intactos nos mesmos cortes ou cortes adjacentes. Os ossos corticais preservados estão em rosa, as porções extrapoladas estão em azul e as regiões 'transplantadas e aparadas' de cortes próximos estão em outras cores. O n.º 900 foi reconstruído após uma pequena correção posicional do pequeno osso indicado pela estrela, que está ligeiramente deslocado para dentro na reconstrução atual.
Em seguida, distalmente, a seção CT nº 250 de SOA-MM9 (Fig. 5 ), que é cortada no ponto hOF, corresponde ao nível de 12,5–14% (ou 11,5–15% mais amplamente) do comprimento máximo (conforme explicado abaixo). Devido às relações alométricas observadas de que úmeros humanos modernos mais curtos tendem a ter epífises distais relativamente grandes (Nota Suplementar 3 ), nos referimos às duas amostras a seguir para nos basearmos nas figuras acima. Uma é a população pré-histórica de baixa estatura do Holoceno da Ilha Tanegashima, Japão (N = 13, comprimento máximo do úmero: 245–292 mm), e a outra é um subconjunto de úmeros curtos da população pré-histórica Jomon do Japão (N = 10, comprimento máximo do úmero: 240–250 mm). As médias dos níveis de hOF nessas amostras foram de 12,5% e 14%, respectivamente, com seus intervalos de 11,5 − 13,5% e 13 − 15%, respectivamente. Os valores equivalentes em úmeros fósseis de hominídeos curtos são de 13% em LB1 e AL 288-1.
Com base nas avaliações acima da preservação do eixo umeral SOA-MM9 (proximal, nível de 50%; distal, nível de 12,5 − 14% ou 11,5 − 15%), o comprimento umeral máximo original do SOA-MM9 é estimado em 211–220 mm ou 206–226 mm, respectivamente. Alternativamente, se aplicarmos as razões médias entre o comprimento do NF-hOF e o comprimento umeral máximo em nossas amostras humanas modernas de homens e mulheres (0,29 a 0,30, Dados Suplementares 2 ), o comprimento máximo estimado do SOA-MM9 é de 194–200 mm, mas supomos que isso seja menos confiável, dada a fraca correlação entre as duas medições ( r = 0,37).
Morfologia comparativa do úmero
SOA-MM9 não possui características do Australopithecus distal humeri, como uma crista supracondilar lateral proeminente em forma de flange, uma crista supracondilar medial projetada e curvatura acentuada no plano sagital, embora a expressão dessas características tenda a ser fraca em espécimes comparativamente gráceis deste gênero 26 , 27 , 28 , 29 . Com relação ao formato da seção transversal (nível do eixo distal de 19%), SOA-MM9 é semelhante ao Homo de corpo pequeno ( H. naledi e H. floresiensis ) por ter um perfil estreito mediolateralmente que é incomum entre os grupos comparativos (Fig. 6 ). É diferente, no entanto, de indivíduos Australopithecus de corpo pequeno , como AL 288-1, cujo úmero é apenas ligeiramente mais longo do que o estimado para SOA-MM9 (Fig. 6 ). As distâncias de Procrustes apoiam a variação do formato da seção transversal resumida pelos resultados do PCA. Enquanto as distâncias de SOA-MM9 para agrupar formas médias (Fig. 4a suplementar ) ou espécimes individuais (Fig. 4b suplementar ) de H. naledi e H. floresiensis não excedem o grau de variação dentro da espécie representado por nossa amostra humana moderna, as mesmas distâncias para os outros táxons fósseis são muito maiores.

uma Análise de componentes principais (PCA) da forma diafisária distal entre 40 úmeros fósseis de hominídeos. Cascos convexos definem os grupos fósseis (Grupos 1-3 = australopiteco, Grupo 4 = H. habilis , Grupo 5 = H. erectus sl ., Grupo 6 = H. naledi : veja Dados Suplementares 3 para mais detalhes) e a linha pontilhada conecta LB1 à forma média das três seções amostradas de SOA-MM9. Os dois contornos diafisários retratam a variação da forma exclusivamente ao longo de PC1. SOA-MM9 é extremo ao longo de PC1 e é mais similar em forma geral (com base na distância de Procrustes) a LB1 e a espécimes de H. naledi (Grupo 6). Veja a Fig. Suplementar 3 para uma apresentação bidimensional deste resultado. b Esboços transversais da diáfise distal do úmero de SOA-MM9 e LB1 em comparação com as médias de grupo para humanos modernos e os grupos morfológicos de hominídeos fósseis. As configurações de forma são mostradas em escala pela largura anteroposterior (com anterior em direção ao topo e lateral à direita).
Canino decíduo maxilar (d c : SOA-MM10)
Este dente direito preserva uma coroa completa e um segmento curto e quebrado da raiz (Fig. 1j ). A coroa é extremamente pequena, situada bem abaixo da faixa relatada de H. sapiens (Fig. 7a e Tabela Suplementar 1 ), de forma semelhante ao Mata Menge d c s relatado anteriormente (Fig. 7b ). O espécime tem um ombro distal primitivo e relativamente baixo que se assemelha aos homólogos Australopithecus e Sangiran H. erectus , embora PCAs baseadas em quatro ou cinco medições lineares indiquem que esta morfologia está marginalmente dentro da grande variação vista em H. sapiens (Fig. 5 Suplementar ). O desgaste oclusal expõe uma pequena mancha de dentina na ponta da cúspide e uma linha fina de dentina na margem incisal distal alongada. Este último sugere a presença de uma configuração primitiva e alta de cúspide mesial em seu primeiro molar decíduo oclusivo (dm 1 ), conforme conhecido para um antigo H. erectus javanês dm 1 de Sangiran, S7–67 30 .

Dimensões horizontais da coroa dos caninos decíduos maxilares (d c : a ) e terceiros molares mandibulares (M 3 : c ), bem como os caninos decíduos mandibulares relatados anteriormente (d c : b ). As grandes cruzes em a e b indicam intervalos de 2 DP para a população moderna com os menores dentes, como para d c e d c (Índia 66 ). d Gráficos de pontuações de PC derivadas da Análise Elíptica de Fourier (EFA) normalizada do contorno da coroa M 3. As diferenças de forma ao longo dos eixos de PC são mostradas nos dentes esquerdos para dois desvios-padrão da origem. Símbolos: 'a' (laranja) = Au. afarensis , 'a' (magenta) = Au. africanus , 'h' (azul) = H. habilis sensu lato , 'D' (laranja) = Dmanisi Homo , 'z' (violeta) = Zhoukoudian H. erectus , 'S' (verde) = H. erectus inicial (Sangiran Lower), 's' (verde) = H. erectus inicial (Sangiran Upper), 'L' (vermelho) = Liang Bua, 'S' (vermelho) = Soa (Mata Menge). 'x' = H. sapiens (Japão) para ( a ) e ( b ) H. sapiens (Japão) para ( c ). Os números dos espécimes são indicados para amostras selecionadas em cada símbolo. Os dados de origem são fornecidos como um arquivo de dados de origem.
Terceiro molar inferior (M 3 : SOA-MM11)
A coroa esquerda preservada tem áreas de cúspides distais reduzidas, hipoconulídeo distalmente saliente e nenhuma faceta interproximal distal (Fig. 1k ). O desgaste achatou grande parte da superfície oclusal, exceto pelo metaconídeo que permanece relativamente alto. Os diâmetros da coroa são comparáveis aos do indivíduo menor de Liang Bua (LB6/1) e estão marginalmente dentro da grande variação exibida por nossa amostra global de H. sapiens (Fig. 7c ). Ela tem cinco cúspides principais dispostas em um padrão '+' e é diferente dos terceiros molares mandibulares de Liang Bua H. floresiensis que exibem uma morfologia derivada de quatro cúspides (LB1, LB6/1) 31 , 32 . O contorno da coroa oclusal examinado pela Análise Elíptica de Fourier normalizada mostra que ela se agrupa firmemente com Sangiran H. erectus e marginalmente com H. ergaster , tendo uma coroa curta mesiodistalmente. Está fora do intervalo de variação exibido por H. habilis sensu lato , que é caracterizado principalmente por uma coroa alongada mesiodistalmente e afilada distalmente (Fig. 7d , Fig. 8 , Nota Suplementar 4 , Fig. Suplementar 6 ) com tendência para hipoconulídeos e cúspides acessórias mais bem desenvolvidos 33 , 34 , 35 .

LB1 e LB6/1: Liang Bua H. floresiensis , Sangiran 22: H. erectus javanês antigo , OH13: H. habilis .
Comparação de tamanho com o Liang Bua H. floresiensis
Em todas as medições disponíveis do corpo mandibular, dentes pós-caninos (P 3 , M 1/2 e M 3 ) e úmero distal, os fósseis de Mata Menge relatados aqui ou anteriormente 9 são menores do que os restos de H. floresiensis de Liang Bua (LB1 e LB6/1) em 1–21% (Tabela 2 ). As estimativas de estatura, com base no comprimento do úmero de SOA-MM9 (211−220 mm) e LB1 (243 mm), são 103–108 cm e 121 cm, respectivamente, usando o modelo de pigmeu humano 36 ; ou 93–96 cm e 102 cm, respectivamente, usando o modelo de macaco 37 .
Discussão
Todos os dez restos de hominídeos descobertos até agora em Mata Menge foram escavados de uma área estreita (cerca de 7 m × 20 m) dentro da parte superior da Camada II (Fig. 2 ). Relatamos anteriormente que um fragmento de mandíbula (SOA-MM4) e seis dentes isolados (SOA-MM1, 2, 5, 6, 7 e 8) desta coleção representam pelo menos um adulto e duas crianças 9 (Tabela 1 ). O desgaste limitado na margem incisal mesial do novo d c direito (SOA-MM10) não corresponde ao desgaste extenso na margem incisal distal do d c direito relatado anteriormente (SOA-MM8), mas o grau de desgaste não exclui a possibilidade de que SOA-MM10 e SOA-MM7 (d c esquerdo ) sejam da mesma criança. Enquanto SOA-MM7 foi recuperado por peneiramento, este espécime e SOA-MM10 foram encontrados a uma distância horizontal de 6 m um do outro. O novo molar permanente (SOA-MM11), um M 3 esquerdo moderadamente desgastado , é obviamente um indivíduo diferente de SOA-MM1, um M 1 esquerdo levemente desgastado (ou M 2 ) pertencente a um adolescente (ou um jovem adulto se este fosse um M 2 ). Portanto, o atual conjunto de hominídeos de Mata Menge inclui pelo menos quatro indivíduos, incluindo um adulto, um adolescente/jovem adulto e duas crianças (Tabela 1 ).
A observação de que todos os quatro (ou mais) indivíduos são extremamente diminutos apoia o argumento de que o tamanho corporal pequeno não era um caráter idiossincrático (individual), mas uma característica populacional dos hominídeos do Pleistoceno Médio inicial de Flores. Os dentes decíduos marcadamente pequenos de pelo menos dois indivíduos, que estão quase fora da grande variação de humanos modernos (Fig. 7 ), também indicam que os hominídeos de Mata Menge tinham tamanho dentário diminuto ao nascer. Além disso, o úmero adulto surpreendentemente pequeno (SOA-MM9) relatado aqui demonstra que esse caráter não se limitava aos elementos dentognáticos, mas também se estendia ao tamanho do braço. Nesta nota, vale destacar que os dois ou mais indivíduos adultos/adolescentes de Mata Menge são consistentemente menores do que os dois adultos de Liang Bua H. floresiensis (Tabela 1 ). Isso sugere fortemente que por volta de ~0,7 Ma, os hominídeos de Flores já eram tão pequenos quanto, ou talvez ligeiramente menores do que, o H. floresiensis do Pleistoceno Superior (Nota Suplementar 5 ).
Com base na amostra dentognática recuperada anteriormente, foi sugerido que os fósseis de Mata Menge poderiam ser razoavelmente atribuídos a H. floresiensis 9 . Agora que um novo osso do braço e restos dentários adicionais pertencentes a esse conjunto exibem fortes afinidades com os restos de Liang Bua, podemos classificar com mais confiança esses hominídeos do Pleistoceno Médio inicial em H. floresiensis . Pequenas diferenças notáveis entre as duas formas cronológicas amplamente separadas incluem a falta de especializações morfológicas molares (veja abaixo) e possivelmente os tamanhos menores do corpo e dos dentes no hominídeo Mata Menge anterior.
Este estudo também contribui para o debate sobre a origem e evolução do H. floresiensis . Foi relatado anteriormente que os hominídeos de Mata Menge agora atribuídos ao H. floresiensis eram mais semelhantes ao antigo H. erectus javanês do que ao Australopithecus e ao H. habilis sensu lato na forma do corpo mandibular e no formato M 1 (ou M 2 ) 9 , uma descoberta que contraria hipóteses que assumem uma ligação evolutiva direta entre o H. floresiensis e os hominídeos pré- H. erectus, como o H. habilis 12 , 13 , 14 . O presente estudo indica que a similaridade de forma entre os fósseis de Mata Menge e o H. erectus javanês antigo também se aplica a M 3 , e que os molares de Mata Menge não têm a especialização única vista nas homologias de H. floresiensis de Liang Bua (ou seja, coroas molares de quatro cúspides, encurtadas mesiodistalmente e um tanto distorcidas 9 , 32 , 37 ). Portanto, o H. floresiensis arcaico em Mata Menge provavelmente representa a linhagem anã do H. erectus javanês antigo em um estágio anterior às especializações molares únicas. Alternativamente, se H. habilis sl . fosse ancestral de Mata Menge/Liang Bua H. floresiensis , este último precisaria ter experimentado uma redução substancial do tamanho molar de ~65–60% nos diâmetros das coroas mesiodistal e bucolingual (de H. habilis significa), e isso acompanhado por mudanças de forma comparáveis à condição do H. erectus javanês antigo . Como não há relações alométricas evidentes entre o tamanho e a forma da coroa molar dentro de H. habilis (Nota Suplementar 6 ), a hipótese de que H. floresiensis é um descendente linear direto de H. habilis sl . não é suportada. Em contraste, o tamanho molar foi reduzido das assembleias dentárias de Sangiran Inferior para Superior (Fig. 7c ) sem mudanças significativas na forma (Fig. 7d ), confirmando que tal evolução local poderia ocorrer. Além disso, embora a morfologia do eixo umeral de SOA-MM9 não indique uma afinidade com H. erectus ou H. habilis , sua forma transversal é mais semelhante à dos táxons anões de Homo ( H. floresiensis e H. naledi ) e diferente daquela de corpos pequenosIndivíduos do Australopithecus .
Juntamente com a data de chegada recentemente revisada para H. erectus em Java (~ 1,1 Ma, ou no máximo mais jovem que 1,3–1,5 Ma) 11 e hominídeos em Flores (1,0–1,27 Ma) 6 , bem como as análises craniométricas e odontométricas relatadas que quase unanimemente apoiam fortes afinidades de H. floresiensis com H. erectus (particularmente H. erectus inicial de Java) 37 , 38 , 39 , 40 , 41 , 42 , o seguinte modelo evolutivo emerge. Os primeiros hominídeos de Flores apareceram nesta ilha Wallaceana ~1,0–1,27 Ma, provavelmente não intencionalmente (ou seja, por meio de 'rafting' acidental, talvez em destroços de tsunami) e possivelmente como parte da colonização inicial da região da Plataforma Sunda pelos primeiros H. erectus . Os hominídeos de Flores experimentaram uma redução substancial no tamanho do corpo logo após esse evento (dentro de ~300.000 anos), apesar da presença de predadores de grande porte, como lagartos-monitores de Komodo de ~3 metros de comprimento e crocodilos, desde o registro paleontológico mais antigo (~1,4 Ma) em diante 6 . Isso implica que os répteis gigantes não representavam uma séria ameaça de predação para o H. floresiensis inicial ou seus progenitores. Esse evento evolutivo inicial foi seguido por estabilidade de longo prazo no tamanho do corpo dos hominídeos, possivelmente também em adaptações culturais (por exemplo, tecnologia de pedras 6 , 7 , 8 ) e especialização morfológica menor na dentição. Como o pequeno tamanho do cérebro relatado para o LB1 1 , 43 de ~60.000 anos evoluiu ainda permanece desconhecido. Atualmente, no entanto, os dados fósseis disponíveis sugerem que o tamanho corporal pequeno foi uma adaptação funcional para esses hominídeos insulares durante e um pouco além do Pleistoceno Médio e, de fato, potencialmente até a chegada do H. sapiens em Flores, há cerca de 50.000 anos; um evento que, suspeitamos, precipitou o desaparecimento do H. floresiensis 10 .
Métodos
A permissão para realizar escavações em Mata Menge foi concedida pelo Ministério de Pesquisa e Tecnologia do Estado da Indonésia (licenças RISTEK 300/SIP/FRP/SM/VIII/2013 e 2183/FRP/SM/X/2015), o governo provincial de East Nusa Tenggara em Kupang e a Administração do Distrito de Ngada. A tomografia computadorizada de SOA-MM9 foi conduzida em Tóquio com uma permissão emitida em 2015 pela Agência Geológica de Bandung.
Tomografia computadorizada e medições
Uma microtomografia computadorizada de SOA-MM9 foi feita usando TXS320-ACTIS (Tesco Co.) no Museu Nacional de Natureza e Ciência, Tóquio, com os seguintes parâmetros de varredura: 205 kV e 0,2 mA com um pré-filtro de placa de cobre de 0,5 mm de espessura, uma matriz de 1024 × 1024, tamanho de pixel de 0,04 mm e intervalo de fatia de 0,04 mm (espessura de fatia de 0,043 mm). As microtomografias computadorizadas (tamanho do voxel = 0,156 mm) de 88 japoneses pré-históricos adultos (caçadores-coletores-pescadores do Holoceno do período Jomon) também foram obtidas para a estimativa do comprimento de SOA-MM9 (veja abaixo). Medidas lineares foram feitas usando um paquímetro digital de espalhamento (para o 0,1 mm mais próximo), uma placa osteométrica (para o 1,0 ou 0,5 mm mais próximo) e fita métrica (para o 0,5 mm mais próximo). As propriedades da seção transversal foram calculadas com base em tomografias computadorizadas em nível vertical de 15–50% do eixo, usando o software CT-Rugle (ver. 1.2, Medic Engineering Inc., Japão) e ImageJ (ver. 1.53f51, National Institutes of Health, EUA).
Análises umerais
Amostras comparativas
Para caracterizar sua morfologia umeral como um grupo especializado de hominídeos insulares, o SOA-MM9 foi comparado com uma ampla variedade de úmeros de hominídeos afro-asiáticos do Plioceno e Pleistoceno ( Ardipithecus , Australopithecus , Paranthropus , Homo africano primitivo , Dmanisi Homo , H. erectus / ergaster e H. naledi ), bem como H. floresiensis de Liang Bua (LB1) e uma série de amostras humanas modernas, incluindo algumas populações de baixa estatura. Os espécimes individuais incluídos para comparação métrica linear e análise morfométrica geométrica são mostrados nos Dados Suplementares 2 e 3 , respectivamente. As amostras humanas modernas usadas para a análise métrica linear estão nos Dados Suplementares 2 , enquanto a amostra humana moderna para a análise morfométrica geométrica (Dados Suplementares 3 ) é uma amostra mista de adultos coletados (por JM Plavcan) no Museu Nacional de História Natural Smithsonian (Washington, DC) 44 . Quanto à análise morfométrica geométrica, a amostra fóssil foi dividida em seis grupos morfológicos, cinco dos quais foram estabelecidos por estudos anteriores 29 , 44 , 45 , 46 , enquanto um sexto grupo consiste em cinco espécimes atribuídos ao Homo naledi . Também coletamos dados de contorno de uma varredura do úmero do esqueleto LB1 (ou seja, LB1/50) atribuído ao H. floresiensis 1 . O status adulto/subadulto para cada espécime foi determinado pela fusão epifisária das extremidades proximal e/ou distal, ou outras informações, se disponíveis (por exemplo, desenvolvimento dentário). As comparações métricas lineares se concentram nas amostras adultas, enquanto a análise morfométrica geométrica contém alguns espécimes subadultos, conforme observado na Fig. 4 e Dados Suplementares 3 . Consulte a Nota Suplementar 7 para obter informações adicionais sobre a amostra de H. naledi .
Principais marcos
O SOA-MM9 não possui a maioria dos marcos osteométricos amplamente utilizados, mas os seguintes pontos são utilizáveis:
Forame nutriente (NF): margem distal do forame nutriente na diáfise média.
Ponto hOF: margem proximal da cavidade que leva à fossa do olécrano.
Idade do desenvolvimento
Os ossos humanos sofrem mudanças histomórficas substanciais durante o desenvolvimento e grande parte da vida adulta. Em uma haste óssea de membro, o crescimento ósseo cortical periosteal ocorre como deposição de osso lamelar circunferencial e ósteons primários com canais não-Haversianos. A proporção dessas estruturas primárias diminui à medida que os ósteons secundários (canal Haversiano cercado por anéis concêntricos de osso lamelar) aparecem e aumentam por meio da remodelação óssea. No final da vida adulta, o osso cortical é dominado por ósteons secundários 17 , 47 . Esse processo é demonstrado numericamente pela contagem ou densidade dos elementos do ósteon secundário (ósteons intactos, ósteons fragmentários, canais Haversianos, baía de reabsorção, etc.) 20 , 48 , 49 , 50 . No entanto, como a taxa dessa mudança histomórfica varia consideravelmente dependendo do locus em um osso, as diferenças regionais devem ser consideradas na estimativa da idade histomorfológica 21 , 51 . Como não havia dados histomorfométricos disponíveis na literatura para o eixo médio do úmero humano, coletamos nossos dados referenciais usando úmeros japoneses modernos. Nossa amostra, que consiste em 10 adultos, 6 adolescentes e 4 crianças, foi desenterrada de cemitérios do período Edo (séculos XVII-XIX d.C.) na cidade de Tóquio e está armazenada no Museu Nacional de Natureza e Ciência, Japão. Com base no procedimento padrão de ossificação 52 , categorizamos os úmeros com epífises completamente fundidas como 'adultos', aqueles com epífise proximal não fundida e epífise distal completamente fundida como 'adolescentes' e aqueles com epífises separadas como 'crianças'. A categoria adolescente foi subdividida em 'adolescente inicial' (fusão apenas na epífise distal, N = 2), 'adolescente médio' (fusão na epífise distal e epicôndilo medial, N = 2) e 'adolescente tardio' (epífise proximal parcialmente fundida, N = 2). Nós (JS) cortamos um pequeno pedaço do osso da diáfise médio-posterior do úmero SOA-MM9, para preparar uma amostra seccional ('HS' na Fig. 1c). A localização desta seção é 6,5 mm distal à borda proximal preservada e 14,5 mm proximal à margem distal do forame nutriente, e presume-se que seja ligeiramente distal à tuberosidade deltoide ausente. A seção obtida cobre uma espessura total de suas superfícies externa (periosteal) a interna (intratecal). Após a incorporação em resina, preparamos uma superfície polida para observação com um microscópio de ultra-alta definição (VHX-7000, Keyence). De cada um dos úmeros humanos modernos, preparamos uma seção cortical na superfície posterior 10 mm distal à extremidade inferior da tuberosidade deltoide. Primeiro, cortamos um pequeno pedaço do osso usando um cortador de diamante. Após a incorporação em resina, uma seção transversal de 70 μm de espessura foi feita por um micrótomo (SP-1600, Leica) para observação em um microscópio de luz comum (ImagerA1, Leica). Nós focamos no terço externo do osso cortical, porque a região periosteal é um campo de crescimento ósseo ativo e é útil para estudos de crescimento histomorfométrico 20 . Para permitir a variação regional mencionada acima, nós examinamos três loci adjacentes: um na linha sagital mediana (campo médio) e os outros em ambos os lados (campos lateral e medial) conforme ilustrado na Fig. 3 . A seção preparada para SOA-MM9 corresponde ao campo lateral. Em cada campo de observação, nós contamos o número de osteons secundários intactos (N.On), fragmentos de osteons (N.Fr), baías de reabsorção (N.Re), canais Haversianos dos osteons secundários/fragmentos de osteons (N.Ca) e canais não Haversianos (N.nCa), conforme definido em outro lugar 20 , 53 . Um 'fragmento de osteon' é um osteon secundário erodido por osteons formados posteriormente. Uma estrutura que atravessa a borda do campo de observação foi contada somente se mais da metade dela estivesse dentro. Usamos os três parâmetros independentes de tamanho a seguir como medidas de crescimento e patologia do osso cortical.
1) Densidade populacional de Osteon (OPD): Este índice amplamente utilizado, calculado aqui como (N.On+N.Fr+N.Re) por área (mm2) 21 , monitora o aumento de estruturas secundárias feitas. Um valor maior de OPD reflete estágio avançado de crescimento.
2) Índice de Canal Haversiano (HCI): É uma proporção dos canais secundários feitos. É calculado como (N.Ca/(N.Ca+N.nCa)), e aumenta de 0 a 1 com o crescimento ósseo.
3)Espessura relativa do osso cortical (rCBt): definimos esse índice como a espessura média do osso cortical dividida pela circunferência mínima de cada diáfise umeral. A primeira é a média das três espessuras do osso cortical nos campos medial, médio e lateral na Fig. 3 , que medimos usando um ImageJ de domínio público (US National Institutes of Health, disponível em https://imagej.nih.gov/ij/ ).
Estimativa de comprimento
Os úmeros de hominídeos fósseis e existentes exibem um padrão uniforme de transição na forma transversal (ou seja, achatamento, ângulo do eixo longo, proporção entre as áreas cortical e total e outras características) dos níveis do eixo médio para o distal (Figs. Suplementares 1 e 2 ). Nós (SM e YK) nos referimos a essas informações para reconstruir o comprimento máximo original do úmero (Martin nº 1a) de SOA-MM9.
Propriedades da seção transversal do eixo
Nós (SM) usamos o CT-Rugle 1.2 (Medic Engineering Co.) para calcular as propriedades da seção transversal da diáfise umeral.
Geometria da seção transversal do eixo
Estudos anteriores de úmeros de hominídeos fósseis demonstraram a utilidade taxonômica do formato transversal da diáfise distal amostrada em ~19% do comprimento total (biomecânico) do úmero a partir da extremidade distal 29 , 44 , 46 , 54 , 55 , 56 . O nível de ~19% de SOA-MM9 foi localizado por YK com base em nossa estimativa de seu comprimento máximo (211–220 mm), que foi convertido para o comprimento biomecânico do úmero usando a razão entre os dois (o primeiro é 1,08% maior em média em nossa amostra de humanos modernos pré-históricos de sexo misto (Jomon): N = 88). O nível de 19% assim localizado está dentro das fatias de TC n.º 573–639. Portanto, escolhemos três fatias, n.º 573, 607 (melhor estimativa) e 639 para a presente análise. Coordenadas bidimensionais foram coletadas por ML de todas as três seções do SOA-M9 seguindo o procedimento descrito anteriormente 44 (ou seja, dois marcos do Tipo 2 nos extremos medial e lateral do espécime e 58 semi-marcos deslizantes nas superfícies anterior e posterior). Configurações de marcos brutos foram sobrepostas no mesmo espaço de forma usando a superposição de Procrustes generalizado de mínimos quadrados ortogonais (GPA) 57 , a GPA foi realizada usando o software tpsRelw 58 e os semi-marcos foram permitidos a deslizar ao longo do contorno diafisário usando o critério de energia de flexão minimizada 59 . Subsequentemente à GPA, as relações morfométricas foram avaliadas com o uso de distâncias de Procrustes (Dp) como uma medida de dissimilaridade de forma 60 e análise de componentes principais (PCA) como um meio de resumo visual (via ordenação) da variação de forma entre os espécimes individuais.
Análise odontológica
SOA-MM10 (canino decíduo maxilar: d c ) foi comparado com a amostra disponível de hominídeos fósseis ( Australopithecus e Homo do Pleistoceno Inferior ), bem como uma amostra de H. sapiens (Tabela Suplementar 2 ) por YK Infelizmente, não há dentes decíduos na assembleia existente de H. floresiensis de Liang Bua. SOA-MM11 (terceiro molar mandibular: M 3 ) foi comparado com H. floresiensis de Liang Bua e seus dois principais candidatos ancestrais reivindicados, H. habilis e H. erectus javanês inicial (Tabela Suplementar 2 ). A amostra dentária de H. erectus javanês inicial examinada neste relatório é de Sangiran, Java Central. Dividimos esta amostra em duas subamostras cronológicas, Sangiran Inferior e Sangiran Superior, seguindo o relatório anterior que demonstrou diferenças morfológicas significativas no tamanho do dente, características da mandíbula, capacidade craniana, etc. 61 , 62 . Medições lineares foram tomadas com base nos espécimes originais ou moldes de alta qualidade por YK usando um paquímetro digital (Mitsutoyo Inc.) ou coletados de outra forma da literatura (Tabela Suplementar 2 ). Os contornos da coroa oclusal de SOA-MM11 foram analisados posteriormente por análise elíptica de Fourier normalizada (EFA padronizado por tamanho), usando as amostras comparativas mostradas na Tabela Suplementar 2 e com base nos métodos detalhados em outro lugar 37 . Em resumo, o contorno oclusal de cada espécime foi obtido de uma fotografia do espécime original ou molde de alta qualidade, de uma forma que minimiza o erro derivado do efeito de paralaxe e orientação do dente ou escala. As imagens foram carregadas no software Canvas X (ACD Systems) para extrair o contorno oclusal e, para um dente desgastado, para reconstruir pequenas partes da coroa perdidas pelo desgaste interproximal. Em seguida, a análise elíptica de Fourier normalizada foi conduzida usando o software SHAPE 1.3 63 , após cada contorno da coroa ter sido alinhado ao longo de seu eixo mesiodistal. Não avaliamos o sexo desses materiais devido à pequena amostra e ao baixo dimorfismo sexual relatado em dentes decíduos humanos modernos 64 .
Resumo do relatório
Mais informações sobre o desenho da pesquisa estão disponíveis no Resumo do Relatório de Portfólio da Nature vinculado a este artigo.
Disponibilidade de dados
Todos os dados gerados ou analisados durante este estudo estão incluídos neste artigo publicado (e seus arquivos de informações suplementares) ou como um arquivo de Dados de Origem. O arquivo de Dados de Origem inclui dados brutos usados para as Figuras 6 e 7 e Figuras Suplementares 2 , 3 , 4 e 6. Os fósseis de hominídeos de Mata Menge estão abrigados no Museu Geológico de Bandung. Os dados 3D do úmero SOA-MM9 podem ser compartilhados mediante solicitação a Unggul P. Wibowo (unggul.pw@esdm.go.id). Os dados de origem são fornecidos neste artigo.
Referências
Brown, P. et al. Um novo hominídeo de corpo pequeno do Pleistoceno tardio de Flores, Indonésia. Nature 431 , 1055–1061 (2004).
Morwood, MJ et al. Mais evidências de hominídeos de corpo pequeno do Pleistoceno tardio de Flores, Indonésia. Nature 437 , 1012–1017 (2005).
Detroit, F. et al. Uma nova espécie de Homo do Pleistoceno Tardio das Filipinas. Nature 568 , 181–186 (2019).
Aiello, LC Handbook of Pa lean thropology (eds W. Henke & I. Tattersall) 2281–2297 (Springer, 2014).
Brumm, A. et al. Idade e contexto dos fósseis de hominídeos mais antigos conhecidos de Flores. Nature 534 , 249–253 (2016).
van den Bergh, GD et al. Uma estrutura geocronológica integrativa para a bacia do Pleistoceno So'a (Flores, Indonésia) e suas implicações para a rotatividade faunística e chegada de hominídeos. Quat. Sci. Rev. 294 , 107721 (2022).
Brumm, A. et al. Hominídeos em Flores, Indonésia, há um milhão de anos. Nature 464 , 748–752 (2010).
Brumm, A. et al. Tecnologia de pedra no sítio do Pleistoceno Médio de Mata Menge, Flores, Indonésia. J. Archaeol. Sci. 37 , 451–473 (2010).
van den Bergh, GD et al. Fósseis semelhantes ao Homo floresiensis do início do Pleistoceno Médio de Flores. Nature 534 , 245–248 (2016).
Sutikna, T. et al. Estratigrafia e cronologia revisadas para Homo floresiensis em Liang Bua na Indonésia. Nature 532 , 366–369 (2016).
Matsu'ura, S. et al. Controle de idade do primeiro dado de aparição do Homo erectus javanês na área de Sangiran. Science 367 , 210–214 (2020).
Argue, D., Groves, CP, Lee, MSY & Jungers, WL As afinidades do Homo floresiensis com base em análises filogenéticas de caracteres cranianos, dentais e pós-cranianos. J. Hum. Evol. 107 , 107–133 (2017).
Dembo, M., Matzke, NJ & Mooers, AO & Collard, M. Análise bayesiana de uma supermatriz morfológica lança luz sobre relações controversas entre hominídeos fósseis. Proc. 282 , 20150943 (2015).
Dembo, M. et al. As relações evolutivas e a idade do Homo naledi : uma avaliação usando métodos filogenéticos bayesianos datados. J. Hum. Evol. 97 , 17–26 (2016).
Yurnaldi, D., Setiawan, R. & Patriani, EY A magnetoestratigrafia e a idade da sequência fóssil da bacia de So'a, Flores, Indonésia. Indones. J. Geosci. 5 , 221–234 (2018).
Powley, MJ, Sutisna, I., Mikac, KM, Wibowo, UP e van den Bergh, GD O leito ósseo de estegodonte do sítio arqueológico do Pleistoceno Médio Mata Menge (Flores, Indonésia): agentes tafonômicos na formação do sítio. Quaternário 4 , 31 (2021).
Gocha, TP, Robling, AG & Stout, SD Antropologia Biológica do Esqueleto Humano , 3ª Edição (eds MA Katenberg & AL Grauer) 145–187 (John Wiley & Sons, 2018 ) .
Martínez-Maza, C., Rosas, A. & García-Vargas, S. Paleohistologia óssea e evolução humana. J. Antropol. Sci. 84 , 33–52 (2006).
Wu, K., Schubeck, KE, Frost, HM & Villanueva, A. Taxas de formação óssea de Havers determinadas por um novo método em um mastodonte, e em diabetes mellitus e osteoporose humana. Calcif. Tissue Res. 6 , 204–219 (1970).
Kerley, ER A determinação microscópica da idade em osso humano. Am. J. Phys. Antropol. 23 , 149–163 (1965).
Gocha, TP & Agnew, AM Variação espacial na densidade populacional de osteons no eixo médio do fêmur humano: adaptações histomorfométricas ao ambiente de carga habitual. J. Anat. 228 , 733–745 (2016).
Schultz, M. Tafonomia Forense: O Destino Pós-Morte de Restos Humanos (eds WD Haglund & MH Sorg) 201–222 (CRC Press, 1997).
Carroll, SE Um estudo sobre os forames nutrientes na diáfise umeral. J. Bone Jt. Surg. 45B , 176–181 (1963).
Larson, SG et al. Descrições do esqueleto do membro superior do Homo. floresiensis. J. Hum. Evol. 57 , 555–570 (2009).
Walker, A. & Leakey, R. O esqueleto do Homo erectus Nariokotome (eds A. Walker & R. Leakey) 7 , 95–160 (Harvard University Press, 1993).
Churchill, SE et al. O ombro, braço e antebraço do Australopithecus sediba. PaleoAnthropology 2018 , 234-281 (2018).
Ward, CV, Kimbel, WH, Harmon, EH & Johanson, DC Novos fósseis pós-cranianos de Australopithecus afarensis de Hadar, Etiópia (1990-2007). J. Hum. Evol. 63 , 1–51 (2012).
White, TD et al. Novas descobertas de Australopithecus em Maka na Etiópia. Nature 366 , 261–265 (1993).
Lague, MR et al. Propriedades transversais da diáfise umeral de Paranthropus boisei: Implicações para a função do membro superior. J. Hum. Evol. 126 , 51–70 (2019).
Grine, FE Comparação da dentição decídua de hominídeos africanos e asiáticos. Cour. Forsch. Senckenberg 69 , 69–82 (1984).
Jacob, T. et al. Restos esqueléticos de Homo sapiens pigmoide australomelanésio de Liang Bua, Flores: afinidades populacionais e anormalidades patológicas. Proc. Natl Acad. Sci. Usa. 103 , 13421–13426 (2006).
Kaifu, Y. et al. Descrições dos restos dentários do Homo floresiensis . Antropol. Sci. 123 , 129–145 (2015).
Tobias, PV Olduvai Gorge, 4: os crânios, endomoldes e dentes do Homo habilis . 4 (Cambridge University Press, 1991).
Wood, B. Koobi Fora Projeto de Pesquisa 4: Restos Cranianos de Hominídeos . 4 (Clarendon Press, 1991).
Kaifu, Y., Aziz, F. & Baba, H. Restos mandibulares de hominídeos de Sangiran: coleção de 1952–1986. 128 , 497–519 (2005).
Hens, SM, Konigsberg, LW & Jungers, WL Estimativa de estatura em hominídeos fósseis: qual modelo de regressão e amostra de referência usar? J. Hum. Evol. 38 , 767–784 (2000).
Kaifu, Y. et al. Morfologia dentária única do Homo floresiensis e suas implicações evolutivas. PLoS ONE 10 , e0141614 (2015).
Lyras, GA, Dermitzakis, MD, Van Der Geer, AAE, Van Der Geer, SB & De Vos, J. A origem do Homo floresiensis e sua relação com processos evolutivos sob isolamento. Antropol. Sci. 117 , 33–43 (2009).
Gordon, AD, Nevell, L. & Wood, B. O crânio do Homo floresiensis (LB1): tamanho, escala e afinidades iniciais do Homo . Proc. Natl Acad. Sci. Usa. 105 , 4650–4655 (2008).
Kaifu, Y. et al. Morfologia craniofacial do Homo floresiensis : descrição, afinidades taxonômicas e implicação evolutiva. J. Hum. Evol. 61 , 644–682 (2011).
Baab, KL, McNulty, KP & Harvati, K. Homo floresiensis contextualizado: uma análise morfométrica geométrica comparativa de amostras humanas fósseis e patológicas. PLoS One 8 , e69119 (2013).
Schroeder, L. et al. Diversidade do crânio na linhagem Homo e a posição relativa do Homo . naledi. J. Hum. Evol. 104 , 124–135 (2017).
Kubo, D., Kono, RT & Kaifu, Y. Tamanho do cérebro do Homo floresiensis e suas implicações evolutivas. Proc. R. Soc. B: Biol. Sci. 280 , 20130338 (2013).
Lague, MR Identificação taxonômica de hominídeos fósseis do Pleistoceno Inferior com base na forma transversal diafisária distal do úmero. PeerJ 3 , e1084 (2015).
Lague, MR et al. Anatomia umeral do esqueleto do membro superior KNM-ER 47000 de Ileret, Quênia: Implicações para identificação taxonômica. J. Hum. Evol. 126 , 24–38 (2019).
Lague, MR e Menter, CG Restos pós-cranianos de hominídeos de Sterkfontein, África do Sul (eds CV Ward e B. Zipfel) (Oxford University Press, 2020).
Jee, WSS Biologia Celular e Tecidual , 4ª edição ( ed. L. Weiss) 212–254 (Urban e Schwarzenberg, 1983).
Thompson, DD A técnica central na determinação da idade na morte em esqueletos. J. Forensic Sci. 24 , 902–915 (1979).
Ericksen, MF Estimativa histológica da idade na morte usando o córtex anterior do fêmur. 84 , 171–179 (1991).
Stout, SD & Paine, RR Comunicação breve: estimativa de idade histológica usando costela e clavícula. Am. J. Phys. Antropol. 87 , 111–115 (1992).
Pfeiffer, S. Variabilidade no tamanho do osteon em populações humanas recentes. Am. J. Phys. Antropol. 106 , 219–227 (1998).
Cunningham, C., Scheuer, L. & Black, S. Osteologia Juvenil do Desenvolvimento , 2ª Edição (Academic Press, 2016).
Dempster, DW et al. Nomenclatura padronizada, símbolos e unidades para histomorfometria óssea: uma atualização de 2012 do relatório da histomorfometria ASBMR. Nomenclatura Comm. J. Bone Miner. Res. 28 , 2–17 (2013).
Senut, B. Humeral outlines in some hominoid primates and in polio-Pleistocene hominids. 56 , 275–283 (1981).
Susman, RL, de Ruiter, D. & Brain, CK Restos pós-cranianos recentemente identificados de Paranthropus e Homo primitivo da caverna Swartkrans, África do Sul. J. Hum. Evol. 41 , 607–629 (2001).
De Lumley, MA & Marchal, F. Les sites préhistoriques de la région de Fejej, Sud-Omo, Ethiopie, dans leur contexte estratigraphique et paléontologique (eds H. de Lumley & Y. Beyene) 340–383 (Edições Recherche sur les Civilizations , 2004).
Rohlf, FJ & Slice, DE Extensões do método de Procrustes para a superposição ótima de marcos. 39 , 40–59 (1990) .
Rohlf, FJ tpsRelw, análise de deformações relativas. versão 1.49 ed. Departamento de Ecologia e Evolução ( Universidade Estadual de Nova York em Stony Brook, 2010).
Bookstein, FL Ferramentas morfométricas para dados de referência: geometria e biologia . (Cambridge University Press, 1997).
Rohlf, FJ Estatísticas de forma: superposições de Procrustes e espaços tangentes. J. Classif. 16 , 197–223 (1999).
Kaifu, Y. et al. Afinidades taxonômicas e história evolutiva dos hominídeos do Pleistoceno inicial de Java: evidências dentognáticas. 128 , 709–726 (2005) .
Kaifu, Y., Indriati, E., Aziz, F., Kurniawan, I. & Baba, H. Paleoantropologia asiática: da África à China e além (eds CJ Norton & DR Braun) 143–157 (Springer, 2010).
Iwata, H. & Ukai, Y. SHAPE: um pacote de programa de computador para avaliação quantitativa de formas biológicas com base em descritores elípticos de Fourier. J. Hered. 93 , 384–385 (2002).
Gil-Donoso, E. et al. Dimorfismo sexual de tecidos dentais caninos decíduos dimensões de populações humanas modernas. Antropol. Sci. 131 , 107–115 (2023).
Ubelaker, DH Restos de esqueletos humanos: escavação, análise, interpretação . 2ª ed. (Taraxacum, 1989).
P, S., Bhavyaa, R., M, SM, Nirmal, L. & Patil, SS Dimensões da coroa de dentes primários - Uma revisão sistemática e meta-análise. J. Forensic Sci. 67 , 1348–1356 (2022).
Ryan, WBF et al. Conjunto de dados de síntese de Topografia Multirresolução Global (GMRT). Geochem. Geophys. Geosyst. 10 , Q03014 (2009).
Agradecimentos
O antigo chefe da Agência Geológica, Banding, Dr. Surono, é reconhecido por seu apoio. Registros geodésicos de achados escavados foram conduzidos por Y. Sopyan (2013) e EE Laksmana e A. Rahmadi (2014). Agradecemos às pessoas das aldeias de Mengeruda e Piga por sua participação nas escavações e seu apoio contínuo. Também agradecemos às seguintes pessoas por suas contribuições em campo durante as escavações de Mata Menge de 2013–2016: T. Suryana, E. Sukandar, A. Gunawan, Widji, AT Hascaryo, E. Setiyabudi, AM Saiful, B. Burhan, PD Moi, B. Alloway, B. Pillans, M. Storey, D. Yurnaldi, M. Moore, T. Sutikna, H. Insani, MRPuspaningrum, I. Yoga, HJM Meijer, S. Hayes e F. Aziz. Agradecemos a Takao Sato e Takashi Sano por sua assistência na condução de análises morfológicas, e a JM Plavcan, C. Ward, M. Domínguez-Rodrigo, F. Di Vincenzo e W. Kimbel pelas varreduras 3D e/ou moldes de úmeros fósseis. Somos gratos às seguintes instituições: Pusat Penelitian Arkeologi Nasional, Senckenberg Research Institute e Natural History Museum Frankfurt, American Museum of Natural History, Sapporo Medical University, Niigata University, National Museum of Nature and Science, Tóquio, The University of Tokyo, St. Marianna University School of Medicine, Tahara Municipal Museum, Kyoto University, Kyushu University, Sasebo City Museum Shimanose Art Center, Okinawa Prefectural Museum and Art Museum, Okinawa Prefectural Archeology Center. Aspectos desta pesquisa foram apoiados financeiramente pela bolsa Discovery do Australian Research Council (DP1093342 para o falecido MJM Morwood e AB), Australian Research Council Future Fellowship (FT100100384 para GDvd.B.), Center for Geological Survey Bandung, Indonésia, Geology Museum Bandung, Indonésia (para IK, RS, IS e UPW), bolsa JSPS KAKENHI (22H00421 para YK e 23K17521 para JS) e National Science Foundation (BCS-0647557 para ML).
Informações do autor
Autores e Afiliações
Contribuições
YK, GDvdB e IK conceberam e lideraram o estudo. As escavações foram lideradas por IK e RS, com assistência de ISUPWAB e GDvdB. As análises morfológicas foram conduzidas por YK, SM, JS e ML com assistência de GS, RTK e TSYK, SM, JS e ML escreveram o manuscrito com contribuições editoriais de todos os coautores.