O registro mais antigo do Pleistoceno de um hominídeo de grande porte do Levante suporta dois eventos de dispersão fora da África
Relatórios Científicos volume 12 , Número do artigo: 1721 ( 2022 )
Resumo
A escassez de fósseis de hominídeos do Pleistoceno na Eurásia dificulta uma discussão aprofundada sobre sua paleobiologia e paleoecologia. Aqui relatamos os primeiros restos de hominídeos de grande porte do corredor levantino: uma vértebra juvenil (UB 10749) do local do início do Pleistoceno de 'Ubeidiya, Israel, descoberto durante uma reanálise dos restos faunísticos. UB 10749 é um corpo vertebral lombar completo completo, com características morfológicas consistentes com Homo sp. Nossa análise indica que UB-10749 era uma criança de 6 a 12 anos no momento da morte, exibindo padrão de ossificação tardia em comparação com humanos modernos. Seu tamanho adulto previsto é comparável a outros hominídeos de grande porte do Pleistoceno da África. As diferenças paleobiológicas entre UB 10749 e outros hominídeos eurasianos primitivos suportam pelo menos dois eventos distintos de dispersão fora da África. Esta observação corresponde a variantes de tradições líticas (Oldowan; Acheulian), bem como vários nichos ecológicos em locais do início do Pleistoceno na Eurásia.
Introdução
A região do Levante, a principal ponte terrestre que liga a África à Eurásia, foi uma rota de dispersão significativa para os hominídeos e a fauna durante o início do Pleistoceno 1 , 2 , 3 . Mas, embora existam numerosos sítios eurasianos do Pleistoceno primitivo, restos fósseis de hominídeos são raros e estão presentes apenas em quatro localidades datadas entre 1,1 e 1,9 Mya 4 , 5 , 6 , 7 , 8 , 9 , 10 , 11 : Dmanisi (Geórgia), Venta Micena (Orce, Granada), Modjokerto e Sangiran (Java, Indonésia) e Sima De Elefante (Atapuerca, Espanha) (Suplemento 2: Tabela 1 ; Fig. 1uma). Em contraste, os sítios da África Oriental do Pleistoceno inicial contendo restos cranianos de Homo são mais abundantes, mas os restos pós-cranianos são mais escassos, e o esqueleto mais bem preservado é o Nariokotome KNM-WT 15000 12 , 13 .

'Localidade do site Uberidya. ( a ) Mapa da África e Eurásia com os principais sítios paleoantropológicos do Pleistoceno. Os círculos pretos denotam locais sem restos osteológicos; círculos vermelhos indicam locais com restos osteológicos humanos. ( b ) A localização do sítio de 'Ubeidiya, ao sul do lago Kineret (mar da Galiléia), nas margens ocidentais do vale do Jordão (círculo vermelho) ( c ) fotografia aérea do plano de escavação de 'Ubeidiya com a localização de camada II-23 onde UB 10749 foi recuperado.
No Levante, o único local deste período de tempo com restos de hominídeos é 'Ubeidiya na escarpa ocidental do Vale do Jordão, que é uma parte do Vale do Rift mais amplo (Suplementar 1: Fig. 1b,c ). Os restos fósseis incluem fragmentos cranianos (UB 1703, 1704, 1705 e 1706), dois incisivos (UB 1700, UB 335) e um molar (UB 1701), identificado como Homo cf. erectus / ergaster 14 , 15 , 16 , 17 , 18 . É importante notar que alguns desses fragmentos foram escavados do solo antes da primeira temporada, enquanto outros são considerados intrusivos e mais jovens do que os depósitos do entorno 17 .
Em 2018, durante uma reanálise das assembleias faunísticas feita por dois dos autores (A. B e MB), foi encontrado um corpo vertebral completo (UB 10749) com características de hominídeos. Este é o primeiro resto pós-craniano de hominídeo encontrado em 'Ubeidiya com segurança atribuído a depósitos do início do Pleistoceno (Ver “ Materiais e métodos ”).
Aqui avaliamos a afinidade taxonômica de UB 10749, sua localização serial ao longo da coluna vertebral, sua idade cronológica e fisiológica no momento da morte, estimamos a altura e o peso do espécime e detectamos quaisquer alterações patológicas ou tafonômicas. Com base em nossas descobertas, exploramos as características únicas de desenvolvimento do UB 10749 dentro do contexto da paleobiologia do Homo primitivo e suas implicações para a dispersão dos hominídeos para fora da África.
Descrição do achado
UB 10749 é um corpo vertebral completo (Fig. 2). A placa superior da vértebra é oval, com superfície irregular, indicando não ossificação da placa terminal vertebral. Da mesma forma, a placa inferior também é oval com bordas póstero-laterais marcadas. Um pequeno buraco é encontrado no centro das placas superior e inferior. A placa inferior é bilateralmente mais larga que a placa superior. As paredes anterior e lateral são lisas e levemente côncavas, ou seja, suas bordas superior e inferior são mais proeminentes que o centro. Não há evidência de fixação das costelas ao corpo nas paredes laterais. A parede posterior pode ser dividida em três partes, os terços central e lateral direito e esquerdo. A parte central é lisa com dois forames nutricionais. Os dois terços laterais estão localizados na junção entre o corpo vertebral e os pedículos. Sua superfície é irregular,2 ). A forma oval do corpo vertebral, a concavidade da placa inferior, a cunha lordótica e a falta de facetas com costelas indicam uma vértebra lombar inferior, ou seja, pré-sacral (PS) 1, PS2 ou PS3 (correspondente a L5, L4 e L3 em humanos modernos).
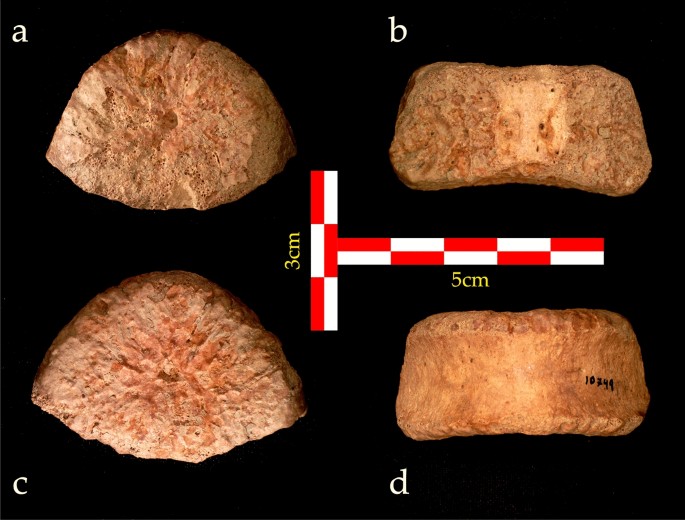
Corpo vertebral UB 10749. ( a ) Vista superior; ( b ) vista posterior; ( c ) vista inferior; ( d ) vista anterior.
Uma varredura de micro-CT (µCT) de UB 10749 (Fig. 3 ) revela um osso cortical bem desenvolvido nas paredes anterior e lateral e na parte central da parede posterior. O osso esponjoso das lâminas superior e inferior é muito fino, assim como o osso dos terços laterais da parede posterior, indicando que estes ainda não estavam ossificados. A µCT também revela canais bem desenvolvidos dentro do corpo vertebral – plexo venoso de Bastons 19 (Fig. 3 c). Uma pequena depressão nas placas superior e inferior é vista nos planos sagital médio e coronal da tomografia computadorizada (Fig. 3 a, b). Uma fina região vertical que aparece em preto no µCT conecta as duas cavidades, indicando que essa área ainda não estava ossificada.

Varredura de µCT de UB 10749. ( a ) Corte sagital médio. ( b ) Corte coronal. ( c ) Seção horizontal.
Identificação taxonômica
Comparamos o UB 10749 com uma variedade de espécies de mamíferos de, mas não limitado a, aqueles presentes em 'Ubeidiya, como carnívoros (por exemplo, Ursus , Hyeana , Panthera ), Artiodactyla (por exemplo, Hippopotamus, Praemegaceros ), Perissodactyla (Rhinocertidae, Equidae) , Proboscidea ( Mamuthus , Elephas ) e Primatas ( Homo , Pongo , Gorilla , Theropithecus e Papio ).
O UB 10749 não possui o recuo interno na parede posterior característico do Ursus e é curto crânio-caudal, em oposição aos corpos vertebrais mais longos dos ungulados.
O tamanho, a grande placa vertebral e o corpo vertebral relativamente curto de UB 10749 indicam que ele pertence a hominoidea. A cunha lordótica e a concavidade da placa inferior sugerem ainda que se trata de uma vértebra hominínea 20 , 21 .
Para estreitar a identificação taxonômica, comparamos UB 10749 a uma variedade de espécies de hominídeos existentes e extintas, e a Pan como um grupo externo (Suplementar 2: Tabela 3 ). A análise revelou que o melhor índice para diferenciar os corpos vertebrais lombares de Homo e Pan é 'comprimento superior à altura posterior' (Fig. 4 ; Suplemento 2: Tabela 4 ). Esse índice também diferencia Homo e Australopithecus 22 . Comparado com as três vértebras pré-sacrais (PS1-PS3) de hominídeos e Pan , UB 10749 está dentro do alcance do Homo e fora do alcance doPan ou Australopithecus . Ele cai perto da posição das vértebras de KNM-WT-15000, um espécime sub-adulto do Pleistoceno do leste da África. Portanto, concluímos que a vértebra em questão provavelmente pertence a um Homo Pleistoceno primitivo .
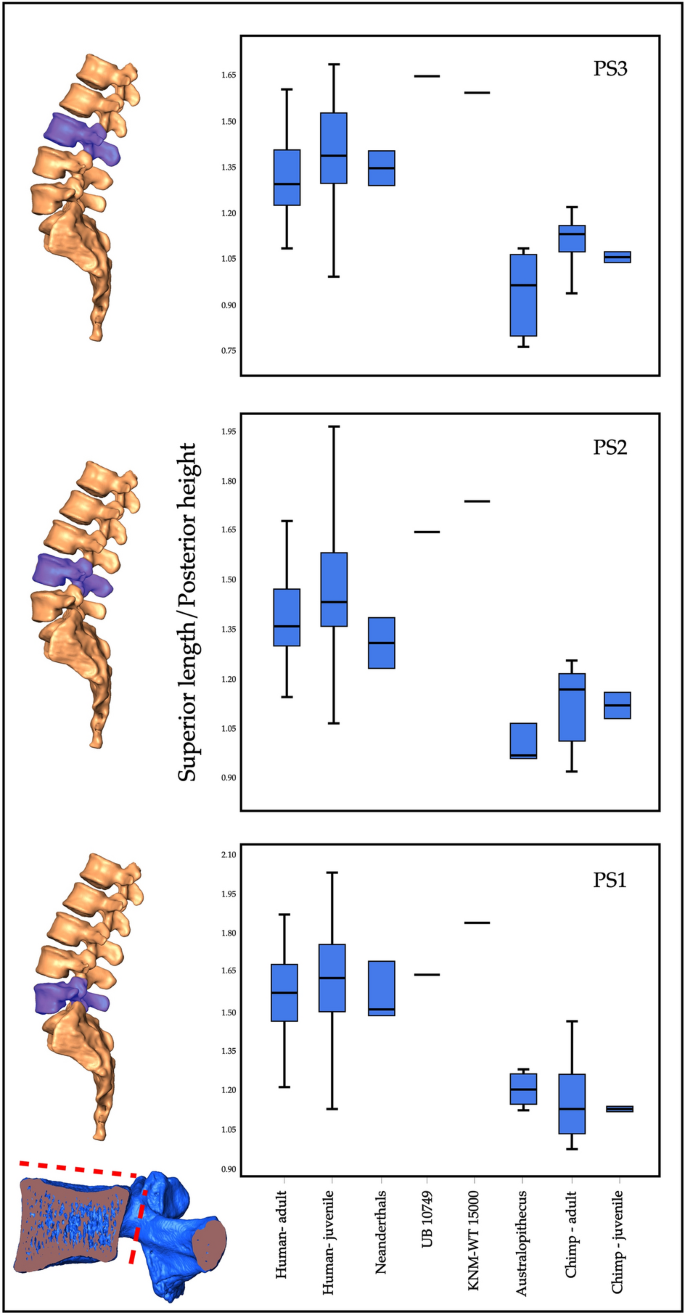
Comparação de UB 10749 com outros hominóides. Proporção do corpo vertebral (comprimento superior para altura posterior) de cada uma das 3 vértebras pré-sacrais inferiores em humanos modernos, neandertais, australopitecos, chimpanzés, KNM-WT 15000 e UB 10749. Observe que UB 10749 está consistentemente dentro da faixa de Homo , e além do alcance dos chimpanzés e australopitecos.
Alocação em série do corpo vertebral
É bem conhecido, principalmente em Hominoidea, que existe uma grande sobreposição na forma dos corpos vertebrais lombares adjacentes 23 . Conduzimos três análises separadas para resolver este problema: (1) Cunha vertebral, ou seja, a razão entre altura posterior/altura anterior que separa significativamente os segmentos vertebrais PS1, PS2 e PS3 de humanos modernos (Suplementar 2: Fig. 1 ; Suplementar 2 : Tabela 4 ), (2) Uma análise de componentes principais (PCA) de índices lineares vertebrais (Fig. 5 a; Suplementar 2: Tabela 4 ) e (3) Análise de forma morfométrica geométrica (GM) (Fig. 5b). A cunha vertebral define UB 10749 como PS2. Os índices lineares vertebrais PCA definem o UB 10749 como PS2 ou PS3, e a análise de forma GM define a vértebra como PS1 ou PS2. Com base nesses resultados, estimamos que a alocação em série de UB 10749 é provavelmente PS2.
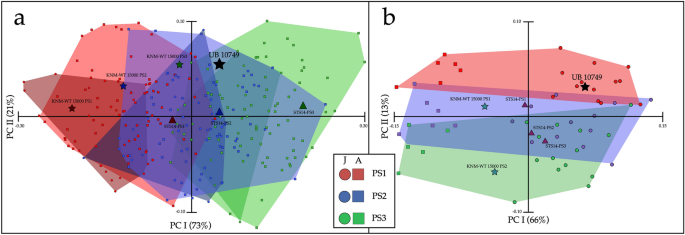
Alocação em série de UB 10749. ( a ) PCA de proporções de corpos vertebrais de humanos modernos, KNM-WT 15000, STS 14 e UB 10749 (ver Tabela Suplementar 4 ). Observe a sobreposição entre adultos e juvenis em cada vértebra pré-sacral. UB 10749 está dentro do alcance de PS2–PS3. Observe que o KNM-WT 15000 e o STS 14 seguem o mesmo padrão dos humanos modernos. ( b ) Resultados de PCA para análise de forma GM. O UB 10749 está dentro do alcance do PS1, mas com proximidade com o PS2. Observe que o KNM-WT 15000 e o STS 14 seguem o mesmo padrão dos humanos modernos. Em ambas as análises: Círculos denota juvenil; Quadrados denota adultos. Vermelho denota PS1; Azul indica PS2; Verde denota PS3.
Idade na morte
A idade à morte é estimada com base no nível de ossificação, tamanho vertebral relativo ou forma vertebral. A falta de ossificação do canal neural em UB 10749 indica uma idade aproximada de 3 a 6 anos em comparação com humanos modernos 24 , 25 (Suplementar 2: Fig. 2 ), embora seja importante notar que vários autores relatam alta variabilidade na ossificação do pedículo, até 16 anos 26 , 27 . A ausência de ossificação da placa terminal vertebral também corrobora a idade jovem de UB 10749, indicando que a vértebra pertence a um indivíduo que não atingiu a puberdade 28 .
Em contraste, com base apenas em seu tamanho, UB 10749 seria atribuído a uma idade mais avançada, provavelmente entre 11 e 15 anos de idade humana moderna (Fig. 6 a: Suplemento 2: Tabela 5 ). No entanto, o tamanho vertebral é altamente variável com a idade, e não podemos descartar uma idade mais jovem ou mais avançada. Finalmente, a análise geométrica da forma do componente principal morfométrico sugere que UB 10749 se enquadra na faixa de humanos modernos de 6 a 10 anos de idade (Fig. 6 b). Isso é confirmado por uma análise discriminante linear que também coloca UB 10749 bem dentro do grupo de 6 a 10 anos (Suplementar 2: Fig. 4 ). Considerando todas as informações acima, estimamos que a idade do óbito para UB 10749 esteja entre 6 e 12 anos.
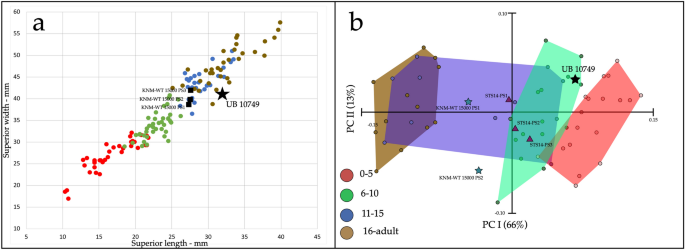
Age at death of UB 10749. (a) Vertebral body size (combined sample of PS1–PS3) of modern humans, KNM-WT 15000 and UB 10749 (see Supplementary 2: Table 5). UB 10749 falls within the range of 11–15 years or the lower end of adults. (b) PCA results for GM shape analysis of modern human, KNM-WT 15000, STS 14, and UB 10749 vertebral bodies. UB 10749 falls within the range of the 6–10 age group. In both analyses: Red, 0–5 years old; Green, 6–10 years old; Blue, 11–15 years old; Brown, 16-adults.
Height and weight estimation
A altura (estatura) e o peso no momento da morte são estimados com base em uma série de equações e gráficos de crescimento para humanos modernos (Suplementar 2: Tabelas 6-8 ). A altura média estimada na morte de UB 10749, aponta para uma altura de 155 cm. Essa altura é comparável a um menino de 13 anos ou a uma menina de 12,5 anos, com base nos gráficos de crescimento do CDC. Uma altura de 155 cm está acima do percentil 95 de 10 anos e acima do percentil 75 para humanos modernos de 12 anos 29 . Como a estimativa de idade para UB 10749 é de 6 a 12 anos, parece que esse indivíduo era alto para sua idade.
O peso é estimado com base na altura ou com base na idade cronológica. Com base na altura, UB 10749 foi de 45 a 55 kg, enquanto com base na idade, o peso de UB 10749 foi de 20 a 43 kg (Suplementar 2: Tabela 7 ). Como a altura é um preditor mais forte para o peso do que a idade de 30 anos , estimamos o peso no momento da morte em cerca de 45-50 kg.
Um único corpo vertebral juvenil não é um preditor definitivo para altura e peso adulto. Ainda mais, o padrão de crescimento dos primeiros hominídeos do Pleistoceno é desconhecido. Assim, para estimar com cautela a altura e o peso do adulto de UB 10749, os cálculos foram baseados em vários métodos: americano moderno (gráficos de crescimento do CDC), população sudanesa moderna 31 e chimpanzés 32 .
Assumindo que UB 10749 tinha de 6 a 12 anos, com base nos gráficos de crescimento dos chimpanzés, teria atingido a altura adulta de 155 a 192 cm e pesava 50 a 101 kg. Com base nos gráficos de crescimento americanos e sudaneses modernos, o UB 10749 apresenta uma faixa de altura entre 168 e 247 cm e um peso entre 62 e 173 kg (Suplementar 2: Tabela 8 ). A predicação média de altura e peso para o tamanho adulto de UB 10749 é de 198 cm e 100 kg. Embora não possamos descartar nenhuma das estimativas, com base em seu tamanho no momento da morte e no tamanho adulto previsto, o UB 10749 era provavelmente um hominídeo de grande porte 33 , 34 , 35 .
Tafonomia
Depósitos fluviais muito finos são evidentes na superfície da vértebra, apesar de serem limpos durante a escavação. Além disso, não há aparente alteração tafonômica ou quebra pós-deposicional.
Patologia
A completude do corpo vertebral e sua simetria bilateral não sugerem processos patológicos ou deformidades de desenvolvimento que possam ter afetado a vértebra, como osteoartrite, hérnia de disco, espondilose, tuberculose, brucelose ou escoliose 36. Entretanto, na ausência do arco vertebral, não podemos descartar o deslizamento anterior do corpo vertebral, ou seja, espondilolistese ou deformidades da articulação facetária. A discrepância entre o tamanho do corpo vertebral e o nível de ossificação é intrigante. O tamanho de UB 10749 é equivalente a um humano moderno de 11 a 15 anos, e o nível de ossificação é equivalente a uma criança humana moderna de 3 a 6 anos. Essa discrepância pode resultar de vários fatores, incluindo condições de desenvolvimento ou patológicas, como: canal notocondral persistente; hipopituitarismo; deficiência androgênica; mutação genética 24 , 37 , 38(ver Suplemento 2 para discussão sobre possível patologia). Embora essas condições sejam raras em humanos modernos, elas não podem ser descartadas. Outra possibilidade é que o UB 10749 apresente um padrão de ossificação diferente do observado em humanos modernos ou grandes símios 25 , 39 .
Discussão
Paleobiologia de UB 10749
A escassez de restos esqueléticos eurasianos do início do Pleistoceno deixa uma lacuna considerável em nossa compreensão da paleobiologia dos hominídeos. A este respeito, o espécime 'Ubeidiya é significativo, fornecendo evidências importantes sobre a diversidade de hominídeos que se dispersaram da África.
Suas características morfológicas indicam que UB 10749 é uma vértebra lombar inferior de um Homo Pleistoceno primitivo . A cunha dorsal do corpo vertebral, o alargamento da placa inferior em comparação com a placa superior, a concavidade da placa inferior e o corpo vertebral alongado ventrodorsalmente em comparação com a altura vertebral são todos traços familiares humanos lombares inferiores 13 , 20 , 21 , 22 .
A afinidade taxonômica dos primeiros hominídeos do Pleistoceno é debatida 40 , e a nomenclatura taxonômica sugerida inclui H. rudolfensis , H. habilis , H. ergaster , H. erectus , H. georgicus e H. antecessor 41 . No início do Pleistoceno, coexistem dois grandes grupos de táxons. Um deles, o hominídeo de corpo pequeno, tradicionalmente representado por H. habilis é caracterizado por corpo pequeno, proporções primitivas de membros, crânio robusto e pequeno volume craniano. Isso se opõe ao hominídeo de corpo grande, representado pelo H. erectus , com proporções de membros semelhantes aos humanos e maior capacidade craniana 42 ,43 . Com base em seu tamanho, o UB 10749 é muito grande para pertencer aos hominídeos de corpo pequeno, como H. habilis sensu lato 33 , 43 , 44 . Nesse sentido, UB 10749 é atribuído ao Homo do Pleistoceno primitivo de corpo grande . Como muito pouco se sabe sobre sua morfologia pós-craniana, uma afinidade taxonômica mais definitiva não é possível.
A altura e o peso estimados ao óbito para UB 10749 se encaixam muito bem com aqueles para KNM-WT 15000 45 . Estima-se que este último seja um indivíduo de 8 anos (com base no desenvolvimento dentário), com uma altura de 159 ± 7 cm (embora veja 46 , 47 para diferentes estimativas) e peso de 49,2 ± 10 kg (com base no bi -largura ilíaca 34 ), comparado a 155 cm e 45–50 kg para UB 10749. O KNM-WT 15000 é um indivíduo alto com nível de ossificação avançado para sua idade cronológica 48 . Em contraste, UB 10749 exibe diferentes padrões ontogenéticos com ossificação atrasada e tamanho aumentado, o que implica que UB 10749 teria crescido mais que KNM-WT 15000.
Comparando KNM-WT 15000 e UB 10749 com o Dmanisi individual D2700/D2735, este último é menor, com 41 kg e 153,1 cm. No entanto, D2700/D2735 quase atingiu seu tamanho adulto, pois espécimes adultos de Dmanisi produziram estimativas gerais entre 145–166 cm e 40–50 kg 49 .
A evidência apresentada aqui demonstra que UB 10749 representa um hominídeo com afinidades compartilhadas com hominídeos de grande porte da África Oriental, como KNM WT 15000, KNM-ER 736 e KNM-ER 1808, MK3(IB7594) 33 , 44 , 50 e, portanto, é a primeira evidência definitiva de um hominídeo de grande porte no corredor levantino.
Implicações paleoecológicas
Estabelecer o ritmo e o modo de dispersão dos hominídeos da África é fundamental na paleoantropologia 51 , 52 , 53 , com pesquisas consideráveis dedicadas à compreensão das forças de empurrar e puxar que influenciam a dispersão dos hominídeos. Embora tenha sido assumido que houve muitos eventos de dispersão 3 , 54 , as dispersões são frequentemente discutidas em termos de preenchimento de um único nicho ecológico para o Homo primitivo sentido lato. Além disso, hipóteses sobre forças intrínsecas ou extrínsecas que podem ter levado ou apoiado essas dispersões são frequentemente testadas em locais que datam de 1,9 a 0,8 milhões de anos, mas analisadas como um único evento de dispersão. No entanto, outros pesquisadores sugerem que a diferença nas assembléias líticas entre Dmanisi (Oldowan) e 'Ubeidiya (Acheulean) refletem eventos de dispersão separados 55 .
Nossa conclusão de que UB 10749 é um hominídeo levantino de grande porte, suporta a ocorrência de várias dispersões do Pleistoceno que não foram apenas separadas no tempo, mas também na ecologia. Dmanisi é reconstruído como pastagem aberta, com condições áridas em comparação com hoje 56 , 57 . Em contraste, o local mais jovem de 'Ubeidiya é reconstruído como mais quente, mas mais úmido do que Dmanisi, com florestas fechadas 58. Segue-se que cada uma das populações de hominídeos associadas às dispersões pode ter exibido adaptações ecológicas e comportamentais únicas. Mais importante ainda, nossa interpretação de que o hominídeo de corpo grande de 'Ubeidiya e o hominídeo de corpo pequeno de Dmanisi não eram da mesma população explica a dificuldade até agora em identificar um único nicho Homo do Pleistoceno inicial 59 , 60. Cronologicamente, Dmanisi antecede 'Ubeidiya por várias centenas de milhares de anos (200-500kya, ver suplementar 1). É possível que tenha havido um único evento de dispersão, seguido de evolução local in-situ que levou a diferentes morfologias de hominídeos em duas regiões. Esta explicação é improvável, pois encontramos hominídeos de corpo pequeno e grande coexistindo na África durante o início do Pleistoceno por quase 500kya 61 . Portanto, a explicação mais parcimoniosa são dois eventos distintos de dispersão “Fora da África”.
No futuro, ao analisar o evento “Out of Africa” do Pleistoceno, devemos estar cientes de que mais de uma única população de hominídeos pode ter se dispersado da África, cada vez com suas próprias diversas adaptações biológicas e culturais.
Materiais e métodos
Materiais
UB 10749 foi encontrado em 1966, da camada II-23. Esta camada foi escavada de 1960 a 1966 por Stekelis, e novamente em 1970 por Bar Yosef 62 , 63 , 64 , 65 (Suplementar 1). UB 10749 foi identificado durante uma reanálise de restos faunísticos, que estão alojados nas Coleções Nacionais de História Natural da Universidade Hebraica de Jerusalém. A vértebra fossilizada apresenta coloração marrom-avermelhada (Fig. 2 ), condizente com a cor do solo da camada II-23 65 .
Observação qualitativa
O UB 10749 foi observado usando três modalidades: descrição macroscópica, microtomografia computadorizada (Triumph® II, triFoil Imaging) e um microscópio digital (microscópio digital Dino-lite edge). Uma amostra comparativa de grandes mamíferos foi estudada no Museu Steinhardt de História Natural da Universidade de Tel Aviv e nas Coleções Nacionais de História Natural da Universidade Hebraica de Jerusalém.
Medidas lineares
Nossa amostra comparativa incluiu adultos e subadultos de humanos modernos (103; 73 adultos, 30 subadultos), hominídeos extintos (13; 10 adultos, três subadultos) e chimpanzés (14; 12 adultos, dois subadultos) (Suplementar 2: Tabela 3 ). Realizamos medições lineares em material ósseo usando o paquímetro Mitotuyu e tomografia computadorizada do banco de dados de imagens do Sheba Medical Center (número de aprovação do comitê de Helsinque: 8266 10 SMC) usando o software Horos (Horosproject.org). Para cada vértebra, realizamos seis medidas lineares: (1) altura anterior, (2) altura posterior, (3) comprimento superior, (4) largura superior, (5) comprimento inferior, (6) largura inferior (Suplemento 2: Tabela 2). Medidas lineares simples podem não indicar taxonomia, idade e serialidade devido à variação significativa no tamanho. Portanto, também calculamos quatro índices para cada vértebra: (1) altura anterior/altura posterior, (2) comprimento superior/largura superior, (3) comprimento inferior/largura inferior e (4) comprimento superior/altura posterior (Suplementar 2 : Tabela 4 ). Para estimativa de idade, comprimento superior e largura superior foram medidos em tomografias computadorizadas de 50 indivíduos adicionais com idade conhecida do banco de dados Sheba Medical Center Imaging (Suplementar 2: Tabela 5 ).
Morfometria geométrica
11 pontos de referência principais foram colocados por um dos autores (AB) nos corpos vertebrais usando dHAL Viewbox (ver. 4.0.1.7, http://www.dhal.com/index.htm ). Nas placas superior e inferior: ponto mais anterior; ponto mais posterior; e dois pontos laterais nas bordas mais externas da vértebra. Três pontos adicionais foram colocados nas paredes anterior e lateral e no ponto médio entre as duas placas. Além disso, oitenta marcas de deslizamento foram automaticamente colocadas no corpo vertebral (Suplemento 2: Fig. 3 ). Para análise do GM, estudamos corpos vertebrais de 20 humanos, variando de 3,5 a 27 anos (PS1–PS3); corpos vertebrais reconstruídos de STS-14 (PS1–PS3) (Suplementar 2: Fig. 5 ); KNM-WT 15000 (PS1–PS2) e UB 10749. No EVAN Toolbox (https://www.evan-society.org/ ), os espécimes foram sobrepostos pelo método de Procrustes. A análise de componentes principais (PCA) foi realizada no conjunto de dados para detectar mudanças relacionadas à forma de serialidade e idade. O primeiro PC (66%) correlaciona-se com as alterações relacionadas à idade e o segundo PC (13%) correlaciona-se com a serialidade vertebral (Figs. 5 e 6 ). A análise discriminante linear foi realizada nos dados sobrepostos (Suplementar 2: Fig. 4 ), para detectar grupos etários humanos modernos e a posição de UB 10749 em relação a esses grupos.
Referências
- 1.
Tchernov, E. A história biogeográfica do sul do Levante. Em The Zoogeography of Israel (eds Yom-Tov, Y. & Tchernov, E.) 159–250 (Dr. Junk Publishers, 1988).
- 2.
Belmaker, M. A presença de um grande cercopithecine (cf. Theropithecus sp.) na formação Ubeidiya (Início do Pleistoceno, Israel). J. Hum. Evoluir 58 , 79-89 (2010).
- 3.
Bar-Yosef, O. & Belfer-Cohen, A. Seguindo os sinais rodoviários do Pleistoceno de dispersões humanas pela Eurásia. Quat. Int. 285 , 30-43 (2013).
- 4.
Gabunia, L. et ai. Os primeiros restos cranianos de hominídeos do Pleistoceno de Dmanisi, República da Geórgia: taxonomia, cenário geológico e idade. Science 288 , 1019-1025 (2000).
- 5.
Gibert, J., Agustí, J. & Moyà, S. Fragmento Cranial atribuido a Homo sp. de Venta Micena (Orce, Granada). Paleontologia e Evolucio (1983).
- 6.
Gilbert, J. et ai. Status hominídeo do fragmento craniano de Orce reafirmado. J. Hum. Evoluir 30 , 203-217 (1998).
- 7.
Palmqvist, PA reavaliação crítica da evidência da presença de hominídeos em tempos do Pleistoceno inferior em Venta Micena, sul da Espanha. J. Hum. Evoluir 33 , 83-89 (1997).
- 8.
Swisher, CC III. et ai. Idade dos primeiros hominídeos conhecidos em Java, Indonésia. Science 263 , 1118-1121 (1994).
- 9.
Huffman, OF Contexto geológico e idade do Perning/Mojokerto Homo erectus, East Java. J. Hum. Evoluir 40 , 353-362 (2001).
- 10.
Larick, R. et ai. Idades 40Ar/39Ar do Pleistoceno Inferior para hominídeos da formação Bapang, Jawa Central, Indonésia. Proc. Nacional Acad. Sci. EUA 98 , 4866-4871 (2001).
- 11.
Lorenzo, C. et ai. Falange da mão humana do Pleistoceno Inferior do sítio da caverna Sima del Elefante (TE) na Serra de Atapuerca (Espanha). J. Hum. Evoluir 78 , 114-121 (2015).
- 12.
Walker, A. & Leaky, R. O Esqueleto Nariokotome Homo Erectus (Harvard University Press, 1993).
- 13.
Meyer, MR & Williams, SA A espinha dorsal do início do Pleistoceno Homo. Em Spinal Evolution: Morphology, Function, and Pathology of the Spine in Hominoid Evolution (eds Been, E. et al. ) 153–184 (Springer, 2019).
- 14.
Stekelis, M. Escavações Arqueológicas em Ubeidiya 1960–1963 (Academia de Ciências de Israel, 1966).
- 15.
Tobias, PV Um Membro do Gênero Homo de Ubeidiya . Academia de Ciências de Israel (1966).
- 16.
Tobias, PV Hominídeos fósseis de Ubeidiya, Israel. Nature 211 , 130-133 (1966).
- 17.
Molleson, TI & Oakley, KP Antiguidade relativa do hominídeo Ubeidiya. Nature 209 , 1268 (1966).
- 18.
Belmaker, M., Tchernov, E., Condemi, S. & Bar-Yosef, O. Novas evidências da presença de hominídeos no Pleistoceno Inferior do Levante Sul. J. Hum. Evoluir 43 , 43-56 (2002).
- 19.
Batson, OV A função das veias vertebrais e seu papel na disseminação de metástases. Ana Surg. 112 , 138-149 (1940).
- 20.
Lakshmanan, P. et ai. Morfologia sagital da placa terminal da coluna lombar inferior. EUR. Coluna vertebral. J. 21 , 160-164 (2012).
- 21.
Plomp, KA, Dobney, K. & Collard, M. Espondilólise e adaptações da coluna vertebral para bipedalismo: A hipótese de superação. Evoluir Med. Saúde Pública 2020 , 35–44 (2020).
- 22.
Been, E. & Rak, Y. The lumbar spine of Australopithecus sediba indicates two hominid taxa. In PaleoAnthropology Society Meeting, Calgary, Canada (2014).
- 23.
Frater, N. Back Problems: Functional, Historical and Paleo-anthropological Aspects (Doctoral dissertation, University of Zurich, 2017).
- 24.
Zhang, H., Sucato, D. J., Nurenberg, P. & McClung, R. Morphometric analysis of neurocentral synchondrosis using magnetic resonance imaging in the normal skeletally immature spine. Spine 35, 76–82 (2010).
- 25.
Rosas, A. et al. The growth pattern of Neandertals, reconstructed from a juvenile skeleton from El Sidrón (Spain). Science 357, 1282–1287 (2017).
- 26.
Blakemore, L., Schwend, R., Akbarnia, BA, Dumas, M. & Schmidt, J. Padrões de crescimento da Sincondrose Neurocentral (NCS) em vértebra cadavérica imatura. J. Pediatra. Ortop. 38 , 181-184 (2018).
- 27.
Rajwani, T. et ai. Características de ressonância magnética da sincondrose neurocentral. Pediatra Radiol. 32 , 811-816 (2002).
- 28.
Martelli, SA A ontogenia espinhal hominóide moderna e fóssil. In Spinal Evolution: Morphology, Function, and Pathology of the Spine in Hominoid Evolution (eds Been, E., Gómez Olivencia, A. & Kramer, PA) 247–281 (Springer, New York, 2010).
- 29.
Kuczmarski, RJ et al. Gráficos de crescimento do CDC para os Estados Unidos: Métodos e desenvolvimento. Estatística de Saúde Vital. 11 (246), 1–190 (2002).
- 30.
Luscombe, MD, Owens, BD & Burke, D. Estimativa de peso em pediatria: uma comparação da fórmula APLS e a fórmula 'Peso = 3 (idade) + 7'. Emerg. Med. J. 28 , 590-593 (2011).
- 31.
Sukkar, MY, Kemm, JR, Ballal, MA & Ahmed, TS Velocidade de crescimento em crianças na zona rural de Cartum, Sudão. Ana Zumbir. Biol. 7 , 473-479 (1980).
- 32.
Ruff, CB & Burgess, ML Quanto mais o KNM-WT 15000 teria crescido?. J. Hum. Evoluir 80 , 74-82 (2015).
- 33.
McHenry, HM Tamanho do corpo e proporções nos primeiros hominídeos. Sou. J. Física. Antropol. 87 , 407-431 (1992).
- 34.
Ruff, C. Previsão do tamanho do corpo de restos esqueléticos juvenis. Sou. J. Física. Antropol. 133 , 698-716 (2007).
- 35.
Ruff, C. Tamanho do corpo e forma do corpo nos primeiros hominídeos – Implicações da pelve de Gona. J. Hum. Evoluir 58 , 166-178 (2010).
- 36.
Boos, N. & Aebi, M. Distúrbios da Coluna - Fundamentos do Diagnóstico e Tratamento (Springer, 2008).
- 37.
Lewis, CP, Lavy, CBD & Harrison, WJ Atraso na maturidade esquelética em crianças do Malawi. J. Cirurgia da Articulação Óssea. Br. 84 , 732-734 (2002).
- 38.
Cotten, A. et ai. Persistência do canal notocordal: filme simples e achados tomográficos. Neurorradiology 37 , 308-310 (1995).
- 39.
Cunningham, DL, Graves, RR, Wescott, DJ & McCarthy, RC O efeito da ontogenia nas estimativas do tamanho do corpo adulto do KNM-WT 15000. J. Hum. Evoluir 121 , 119-127 (2018).
- 40.
Wolpoff, MH Interpretações da evolução multirregional. Science 274 , 704-707 (1996).
- 41.
Wood, B. & Richmond, BG Evolução humana: Taxonomia e paleobiologia. J. Anat. 197 , 19-60 (2000).
- 42.
Dunsworth, H. M. Origin of the genus Homo. Evol. Edu. Outreach 3, 353–366 (2010).
- 43.
Will, M. & Stock, J. T. Spatial and temporal variation of body size among early Homo. J. Hum. Evol. 82, 15–33 (2015).
- 44.
Will, M., Pablos, A. & Stock, J. T. Long-term patterns of body mass and stature evolution within the hominin lineage. R. Soc. Open Sci. B 4, 171339 (2017).
- 45.
Dean, M. C. Measures of maturation in early fossil hominins: Events at the first transition from australopiths to early Homo. Philos. Trans. R. Soc. B. 371, 20150234 (2020).
- 46.
Graves, RR, Lupo, AC, McCarthy, R., Wescott, D. & Cunningham, D. Quão robusto foi o KNM-WT 15000?. J. Hum. Evoluir 59 , 542-54 (2010).
- 47.
Bastir, M. et ai. A anatomia da caixa torácica no Homo erectus sugere uma origem evolutiva recente da forma do corpo humano moderno. Nat. Eco Evoluir 4 , 1178-1187 (2020).
- 48.
Smith, BH A idade fisiológica de KNM-WT 15000. Em: O esqueleto Nariokotome Homo erectus , (Eds Walker, A. & Leakey, R.) 195–220 (Harvard University Press, 1993).
- 49.
Lordkipanidze, D. et ai. Evidência pós-craniana do Homo primitivo de Dmanisi, Geórgia. Natureza 449 , 305-310 (2007).
- 50.
Di Vincenzo, F. et al. The massive fossil humerus from the Oldowan horizon of Gombore I, Melka Kunture (Ethiopia, > 1.39 Ma). Quat. Sci. Rev. 122, 207–221 (2015).
- 51.
Prat, S. First hominin settlements out of Africa Tempo and dispersal mode: Review and perspectives. Comptes Rendus Palevol. 17, 6–16 (2018).
- 52.
Rolland, N. The early Pleistocene human dispersals in the circum-Mediterranean basin and initial peopling of Europe: Single or multiple pathways?. Quat. Int. 316, 59–72 (2013).
- 53.
Dennell, R. Dispersal and colonisation, long and short chronologies: How continuous is the Early Pleistocene record for hominids outside East Africa?. J. Hum. Evol. 45, 421–440 (2003).
- 54.
Shea, J. J. Stone age visiting cards revisited: A strategic perspective on the lithic technology of early hominin dispersal. In Out of Africa 1: The First Hominin Colonization of Eurasia (eds Fleagle, J. G. et al.) 47–64 (Springer, 2010).
- 55.
Bar-Yosef, O. & Belmaker, M. Early and Middle Pleistocene faunal and hominins dispersals through Southwestern Asia. Quat. Sci. Rev. 30, 1318–1337 (2011).
- 56.
Messager, E., Lordkipanidze, D., Kvavadze, E., Ferring, C. & Voinchet, P. Reconstrução paleoambiental do sítio Dmanisi (Geórgia) com base em dados paleobotânicos. Quat. Int. 223 , 20-27 (2010).
- 57.
Blain, HA, Agustí, J., Lordkipanidze, D., Rook, L. & Delfino, M. Contexto paleoclimático e paleoambiental dos hominídeos do Pleistoceno Inferior de Dmanisi (Geórgia, Cáucaso Menor) inferidos da assembleia herpetofaunal. Quat. Sci. Rev. 105 , 136-150 (2014).
- 58.
Belmaker, M. Insights da composição da comunidade carnívora sobre a paleoecologia dos primeiros sítios eurasianos do Pleistoceno: Implicações para a dispersão de hominídeos para fora da África. Quat. Int. 464 , 3-17 (2018).
- 59.
Martínez-Navarro, B. Faunas do Pleistoceno Inferior da Eurásia e dispersões de hominídeos. In: Fora da África I. A Primeira Colonização Hominina da Eurásia (Eds. Fleagle, JG, Shea, JJ, Grine, FE, Baden, AL, Leakey, RE) 207–224 (Springer, 2010).
- 60.
Dennell, R. Migração humana e ocupação da Eurásia. Episódios 31 , 207–210 (2018).
- 61.
Spoor, F. et ai. Implicações de novos fósseis de Homo primitivos de Ileret, a leste do Lago Turkana, no Quênia. Natureza 448 , 688-691 (2007).
- 62.
Picard, L., Baida, U. Relatório Geológico sobre os Depósitos do Pleistoceno Inferior das Escavações de Ubeidiya . Academia de Ciências e Humanidades de Israel (1966).
- 63.
Picard, L., Baida, U. Posição estratigráfica da formação Ubeidiya (Academia de Ciências e Humanidades de Israel, 1966).
- 64.
Bar-Yosef, O., Tchernov, E. Sobre a História Paleoecológica do Sítio de Ubeidiya (Academia de Ciências e Humanidades de Israel, 1972).
- 65.
Bar-Yosef, O. & Goren-Inbar, N. Os conjuntos líticos de Ubeidiya: Um local paleolítico inferior no Vale do Jordão. Qedem 34 , 266 (1993).
Reconhecimentos
Gostaríamos de dedicar este artigo aos falecidos Ofer Bar Yosef e Eitan Tchernov que estiveram envolvidos com as escavações de 'Ubeidiya por mais de 30 anos e contribuíram para nossas discussões sobre o tema de fora da África e a paleobiologia do Homo primitivo. Agradecemos a Rivka Rabinovich das Coleções Nacionais de História Natural da Universidade Hebraica de Jerusalém por fornecer acesso ao UB 10749 e à fauna de mamíferos de 'Ubeidiya; Tamar Dayan e Amos Belmaker, do Museu Steinhart de História Natural da Universidade de Tel Aviv, por fornecerem acesso a amostras comparativas de mamíferos; os Museus Nacionais do Quênia, Emma Mbua, Fredrick Kyalo Manthi e Fred Spoor por fornecer as tomografias computadorizadas do KNM-WT 15000. Este estudo foi financiado pela National Science Foundation para Miriam Belmaker (concessão número 1851613) e pelo Ministério da Ciência espanhol e Inovação para Markus Bastir (CGL2015-63648-P; Ministério da Economia e Competitividade da Espanha e PID2020-115854GB-I00; Ministério da Ciência e Inovação da Espanha) e pelo Programa de Bolsas de Pesquisa da Universidade de Tulsa. Mapas na Fig.1 a, b foram criados por Emil Aladjem e Autoridade de Antiguidades de Israel. A fotografia aérea do local de Ubeidiya na Fig. 1 c foi tirada por Alex Wiegmann, Autoridade de Antiguidades de Israel, como parte deste projeto em andamento.
Informação sobre o autor
Afiliações
Contribuições
AB, EB, MB e OB conceberam e escreveram o artigo. AB e EB coletaram e analisaram os dados. Todos os autores discutiram os resultados e revisaram o artigo. Todos os autores deram aprovação final para publicação.
autor correspondente
Declarações de ética
Interesses competitivos
Os autores declaram não haver interesses conflitantes.
Informação adicional
Nota do editor
A Springer Nature permanece neutra em relação a reivindicações jurisdicionais em mapas publicados e afiliações institucionais.
Informação suplementar
Direitos e permissões
Acesso Aberto Este artigo está licenciado sob uma Licença Creative Commons Atribuição 4.0 Internacional, que permite o uso, compartilhamento, adaptação, distribuição e reprodução em qualquer meio ou formato, desde que você dê os devidos créditos ao(s) autor(es) original(is) e à fonte, fornecer um link para a licença Creative Commons e indicar se foram feitas alterações. As imagens ou outro material de terceiros neste artigo estão incluídos na licença Creative Commons do artigo, salvo indicação em contrário em uma linha de crédito para o material. Se o material não estiver incluído na licença Creative Commons do artigo e seu uso pretendido não for permitido por regulamentação legal ou exceder o uso permitido, você precisará obter permissão diretamente do detentor dos direitos autorais. Para ver uma cópia desta licença, visitehttp://creativecommons.org/licenses/by/4.0/ .
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.