Descrevemos um vampiropode excepcionalmente bem preservado, Syllipsimopodi bideni gen.
et sp. nov., do Carbonífero (Mississipiano) Bear Gulch Lagerstätte de
Montana, EUA. O espécime possui um gládio e dez braços robustos com
fileiras bisseriadas de ventosas; é o único vampyropod conhecido a
manter a condição ancestral de dez braços. Syllipsimopodi é
o mais antigo vampiropode definitivo e coleoide coroa, retrocedendo o
registro fóssil desse grupo em ~81,9 milhões de anos, corroborando as
estimativas do relógio molecular. Usando uma filogenia Bayesiana datada
de cefalópodes neocoleóides fósseis, demonstramos que Syllipsimopodi é
o vampiropode conhecido mais antigo divergente. Isso desafia fortemente
a hipótese comum de que os vampiropodes descenderam de um belemnóide
fragmoteutídeo do Triássico. Já no Mississipiano, os vampiropodes eram
evidentemente caracterizados pela perda do fragmocone com câmaras e do
rostro primordial – traços retidos nos belemnoides e em muitos
decabraquios existentes. Um par de braços pode ter sido alongado, o que,
quando combinado com o longo gládio e as barbatanas terminais, indica
que a morfologia dos primeiros vampiropodes se assemelhava
superficialmente às lulas existentes.
Introdução
Vampyropoda,
o clado que combina octopods, vampyromorphs e seus parentes, é um dos
três principais grupos de cefalópodes coleóides (de casca interna),
sendo os outros dois Decabrachia (lulas, chocos, lulas bobtail e Spirula ) e o extinto Belemnoidea (Fig. . 1 ). Os belemnoides fortemente mineralizados são os coleoides fósseis mais comuns, mas parecem não ter descendentes vivos 1 , 2 e,
portanto, fornecem informações limitadas sobre a evolução dos grupos
existentes (embora alguns tenham proposto que sepiids e/ou espirulids
são belemnoids vivos 3 , 4 , 5 ).
Embora menos comuns, os vampiropodes fósseis não mineralizados são
conhecidos de vários lagerstätten do Mesozóico – sua bioquímica única
parece melhorar o potencial de preservação de seus tecidos moles 6 .
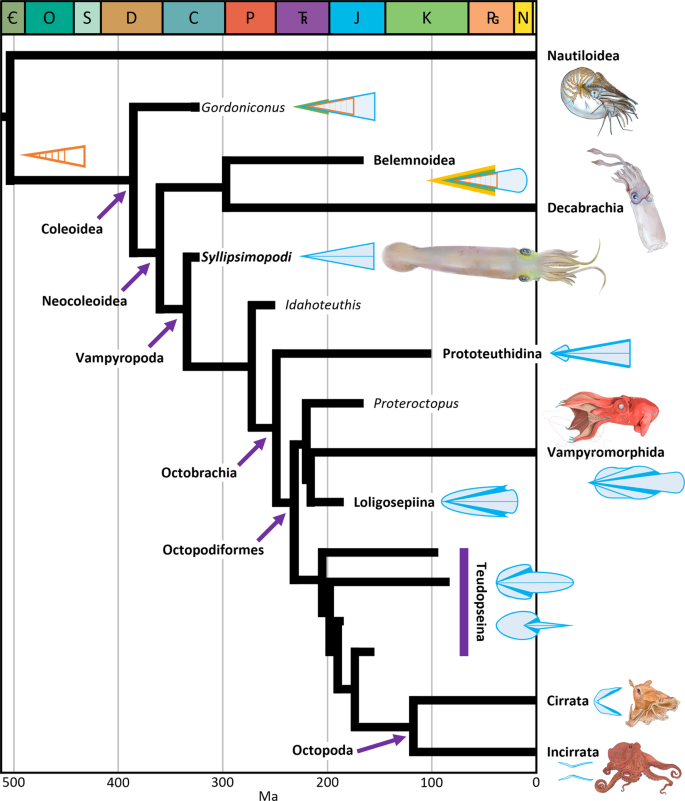
Fig. 1: Visão geral das inter-relações neocoleóides e estimativas de tempo de divergência, mostrando a posição de S yllipsimopodi bideni gen. et sp. novembro
Com base em nossa reconstrução filogenética Bayesiana datada de ponta (Fig. 6 ).
Conchas codificadas por cores: azul = proóstraco/gladius (zonas
hiperbolares e reforços laterais em azul mais escuro), laranja =
fragmocone, verde = rostro primordial, amarelo = rostro. Abreviaturas do
período geológico (cores da Comissão Internacional de Estratigrafia): =
Cambriano (verde escuro), O = Ordoviciano (azul), S = Siluriano (azul
claro), D = Devoniano (marrom), C = Carbonífero (azul), P = Permiano
(laranja vermelho), T R = Triássico (roxo) ), J = Jurássico (ciano), K = Cretáceo (verde), P G =
Paleogeno (laranja), N = Neogene (amarelo), não rotulado = Quaternário
(amarelo pálido). As setas roxas indicam nós nomeados, a barra roxa
indica o grau teudopseid. Representações artísticas criadas por K.
Whalen.
O
gládio é um remanescente de concha achatado encontrado em
vampiromorfos, vampiropodes extintos e muitos decabraquios existentes;
alguns octopods existentes retêm um gladius vestigial na forma de
suportes de barbatanas (cirrates) ou estiletes (incirrates) 2 , 7 , 14 . O gládio é tipicamente composto de três partes (Fig. 2 )
- um campo central mediano (ou raque) é lateralmente flanqueado por
zonas hiperbolares (ou palhetas) que são elas próprias flanqueadas por
campos laterais (ou asas); além disso, pode haver bandeiras de cone
lateral aos campos posteriores (Fig. 2 ) 2 , 14 .
As zonas são frequentemente distinguidas pelas orientações das linhas
de crescimento: campos medianos anteriormente convexos e campos laterais
anterolateralmente convexos são separados por zonas hiperbolares
anteriormente côncavas 2 (Fig. 2 ).
A maior parte do gládio está situada dorsalmente, mas a parte posterior
geralmente tem um componente ventral e lateral muito mais curto,
estendendo-se anteriormente do ápice em forma de taça ou cone; este é o
cone 2 .
Fig. 2: Desenho idealizado de vampyropod gladius (baseado em Vampyroteuthis ).
Mostrando
campo mediano, zonas hiperbolares, campos laterais e bandeiras de cone,
com exemplos de linhas de crescimento. Assíntotas denotam bordas de
zonas hiperbolares.
O mais antigo vampiropode definitivo conhecido foi considerado um espécime fragmentado e mal preservado do prototeutídeo Germanoteuthis do Triássico Médio (Ladiniano, ~241,5–237,0 Ma) 7 , 8 .
No entanto, estimativas de tempo de divergência molecular sugerem que
os vampiropodes divergiram dos decabraquios no Carbonífero-Permiano
(289,7 ± 37,6 Ma) 9 . Pohlsepia 10 foi
proposto como um octopod cirrato carbonífero, mas isso é muito
controverso e parece improvável que o fóssil seja um cefalópode ou
molusco 7 , 9 , 11 , 12 , 13 .
Aqui descrevemos o vampiropode divergente mais antigo e mais antigo, Syllipsimopodi bideni gen. et sp. novembro (Figs. 2 , 3 ), do Mississipiano (Serpukhovian, ~330,3–323,4 Ma) 8 Bear
Gulch Lagerstätte de Montana, EUA. Possui um gládio, barbatanas, 10
braços e ventosas - evidência de uma afinidade coleoide. O fóssil
demonstra que os vampiropodes se originaram no Paleozóico, corroborando
as estimativas do relógio molecular 9 e
estendendo a faixa estratigráfica de vampiropodes fósseis conhecidos em
~81,9 milhões de anos. Nós revisamos e expandimos substancialmente o
Sutton et al. 1 , 2 táxons de caracteres morfológicos neocoleóides para colocar Syllipsimopodi em uma estrutura filogenética Bayesiana com datação de ponta usando o modelo Fossilized Birth-Death (FBD) (Dados Suplementares 1 ). Todos os principais grupos neocoleóides são cobertos, com ênfase nas espécies fósseis portadoras de gladius.
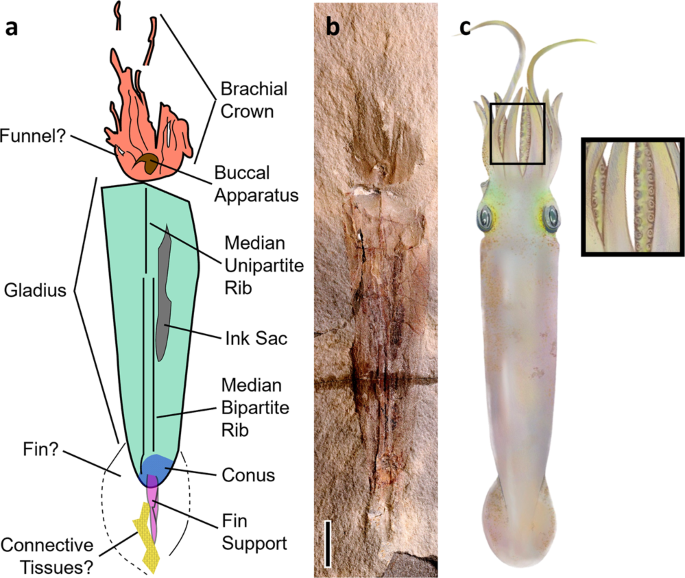
FIG. 3: Syllipsimopodi bideni gen. et sp. nov., holótipo ROMIP 64897.
um desenho esquemático de Syllipsimopodi bideni gen.
et sp. novembro; cerceta = gladius, laranja = cabeça (incluindo
braços), marrom = aparelho bucal, cinza = saco de tinta, azul = cone,
magenta = suporte da nadadeira, amarelo estampado = manchas semelhantes a
escamas (possível remanescente de tecido conjuntivo). b Imagem de cores falsas de contraste aumentado de Syllipsimopodi , holótipo ROMIP 64897. Escala = 1 cm. c Reconstrução artística mostrando ventosas (criada por K. Whalen).
Etimologia .
O nome do gênero é derivado do grego συλλήψιμος (syllípsimos) para
preênsil e πόδι (pódi) para pé. O nome preênsil-pé é escolhido porque
este é o cefalópode mais antigo conhecido a desenvolver ventosas,
permitindo que os braços, que são modificações do pé do molusco, agarrem
melhor presas e outros objetos. O nome da espécie é para celebrar o
recém-inaugurado (no momento da submissão) 46º Presidente dos Estados Unidos, Joseph R. Biden.
Holótipo. ROMIP 64897 (Museu Real de Ontário).
Material. O
tipo e único espécime foi doado ao Royal Ontario Museum por B. Hawes em
1988; número de acesso 88-72717. Não há contrapartida.
Localidade. Calcário Bear Gulch, Heath Formation, Big Snowy Group, Fergus County, Montana, EUA 18 .
O calcário de Bear Gulch é um plattenkalk, ou calcário litográfico,
semelhante ao mais famoso calcário jurássico Solnhofen da Alemanha 19 . A deposição ocorreu em uma baía marinha rasa de baixa latitude sujeita a condições semiáridas e tropicais oscilantes 18 . A preservação excepcional é provavelmente resultado de microturbiditos depositados por monções sazonais 18 .
As chuvas de monção teriam introduzido rapidamente sedimentos
terrestres e biomatéria na baía, alimentando florações de algas que
criaram zonas anóxicas de curta duração simultâneas com a instabilidade
salina causada pela rápida injeção de volumosa água doce 18 . Bear Gulch é talvez mais conhecido pela fauna pelágica da bacia central e foz da baía - uma gama diversificada de vertebrados 20 , 21 , 22 (especialmente condrichthyes 22 , 23 , 24 , 25 e celacantiformes 22 , 26 ), malacostracans 19 , 27 , poliquetas 22 e cefalópodes 13 , 28 , 29 , 30 , que são preservados em detalhes tão requintados que a vascularização às vezes pode ser distinguida 31 . Fósseis bentônicos são muito raros na bacia central 18 , 22 ,
mas fácies marginais preservam gastrópodes, vermes, asterozoários e
esponjas abundantes, que atuaram como substrato para diversos
braquiópodes, bivalves e conularídeos 18 , 19 , 22 . Crinóides, blastóides, briozoários e corais estão quase ausentes; algas (especialmente dasyclads) são comuns em todo 18 , 22 .
Diagnóstico. (Figs. 3 , 4 )
Coleóide com gládio simples, quase triangular, com cone em forma de
funil e campo mediano com costela mediana, mas sem zonas hiperbolares,
bandeiras de cone ou reforços laterais; campos laterais improváveis.
Ausência de fragmocone com câmara, rostro primordial ou rostro. Dez
braços com fileiras bisseriadas de ventosas, mas sem ganchos ou cirros;
dois braços podem ser alongados (embora isso possa ser tafonômico). Saco
de tinta presente. Suporte de aleta mediana terminal e um par de aletas
presentes.
Fig. 4: Syllipsimopodi bideni gen. et sp. nov., holótipo ROMIP 64897, mostrando a coroa do braço.
a – d Escala = 1 cm. um fóssil de corpo completo. b – d Mostrando a coroa do braço; c traços
do braço em azul, roxo indica que o braço está sobreposto abaixo de
dois outros braços, verde indica que o braço está sobreposto acima de si
mesmo; d círculos vermelhos e amarelos marcam as ventosas individuais. e – g = 5 mm; closeup de braços mostrando otários, selecione otários indicados com setas brancas.
Descrição .
O campo mediano do gládio é simples; é mais largo no extremo anterior
com bordas laterais retas e uma borda anterior plana (não
arredondada/pontiaguda) (Figs. 3a , 4 ).
O ângulo assíntota mediano é ~13,8°. A proporção entre o comprimento e a
largura do gládio é de 3,17; o campo mediano tem ~6,5 cm de
comprimento, que é ~55% do comprimento total do corpo. O campo mediano
apresenta uma costela mediana proeminente, que é diageneticamente
distorcida e quebrada em vários lugares (Fig. 2 ). A costela parece ser bipartida posteriormente e unipartida anteriormente (Figs. 2 , 3 ). As linhas de crescimento do gladius são mal preservadas e claramente visíveis apenas em um local na borda lateral (Fig. 3 ). O cone em forma de funil tem ~6,8 mm de comprimento (Figs. 2 , 3 ).
Um suporte de barbatana central em forma de charuto medindo ~13,1 mm de comprimento é preservado posteriormente ao gládio (Fig. 3 suplementar 4 );
uma composição originalmente cartilaginosa parece mais provável. É
possível que este suporte de barbatana seja um vestígio do fragmocone,
mas consideramos esta alternativa improvável porque não há evidência de
um sifúnculo ou septo. É improvável que estes tenham sido dissolvidos
sem deixar vestígios, uma vez que os septos podem ser claramente
observados no coleóide co-ocorrente Gordoniconus 13 . Os septos são dissolvidos em amonóides Bear Gulch 30 ; mas como Gordoniconus é
um coleóide, nós o consideramos um comparador tafonômico melhor. Além
disso, o suporte da nadadeira é posterior e, portanto, externo ao cone; o
fragmocone deve ser interno ao cone (Fig. 5 ).
Consideramos uma identidade de tribuna primordial improvável porque
parece duvidoso que uma tribuna primordial (ou tribuna) estaria presente
na ausência de um fragmocone. O suporte da nadadeira está associado a
manchas de um mineral fibroso brilhante (Fig. 3 ; suplementar 4 ),
presumivelmente remanescentes de tecido conjuntivo. O holótipo parece
preservar um esboço fraco de um único par de barbatanas lobadas
terminais curtas medindo ~2,3 cm de comprimento anteroposterior e ~1,7
cm lateralmente na posição mais larga (Figs. 3 , 4 ).
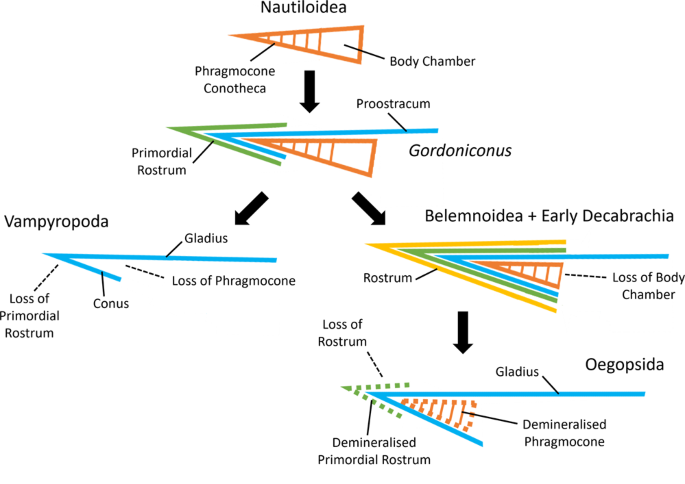
Fig. 5: Visão geral da evolução da concha coleóide, mostrando nossas interpretações do gládio/proóstraco.
Coleóides primitivos, como Gordoniconus 13 , acrescentam o rostro primordial 85 e o proóstraco 14 ; os vampiropodes perdem o fragmocone e o rostro primordial, o proóstraco passa a ser um gládio 14 ; os belemnoides e os primeiros decabraquios perdem a câmara do corpo 86 e acrescentam o rostro 85 ; os oegopsídeos perdem o rostro, alguns retêm um rostro primordial desmineralizado 85 e o fragmocone 71 , o proóstraco passa a ser um gládio 14 .
Estruturas rotuladas apenas quando aparecem (linha preta sólida) ou são
perdidas (linha preta tracejada). Tecidos da concha: laranja =
fragmocone + câmara do corpo (tracejado = desmineralizado), azul =
proóstraco/gládio, verde = rostro primordial (tracejado =
desmineralizado), amarelo = rostro.
Dez braços com ventosas são preservados, medindo ~2,1–2,4 mm de largura no comprimento médio (Fig. 4b, c ).
Evidências de desenvolvimento e inferência filogenética têm sugerido há
muito tempo que a condição de dez braços é ancestral para cefalópodes e
vampiropodes 33 , 34 , 35 , mas nenhum fóssil de dez braços foi documentado fora do clado decabraquiano-belemnóide antes da descoberta de Syllipsimopodi . Syllipsimopodi é
o primeiro e único vampiropode conhecido a possuir dez apêndices
robustos e funcionais; todos os outros vampiropodes conhecidos reduziram
o par de braços II a filamentos (ou seja, Prototeuthidina 7 , Loligosepiina 7 , 36 , Teudopseina 7 , Vampyromorphida), ou perderam o par de braços inteiramente (ou seja, Octopoda).
Dois braços,
medindo ~4,0 e ~4,1 cm de comprimento (~27% do comprimento total do
corpo), podem ter sido alongados em relação aos outros oito braços (Fig.
4b, c ).
Esses braços alongados não têm nenhuma mão óbvia, como nos tentáculos
decabraquianos, e não são significativamente mais finos do que os braços
não modificados, como nos filamentos de vampiromorfos (Fig. 4b, c ).
Dos braços mais curtos, os três mais bem preservados medem ~1,7, ~1,9 e
~1,9 cm de comprimento, respectivamente (~13% do comprimento total do
corpo); os cinco braços restantes estão incompletos ou preservados em
uma orientação contorcida (Fig. 4b, c ). Todos os braços parecem ter ventosas ao longo da base e do comprimento médio (Fig. 4d ).
Os braços mais curtos mais bem preservados parecem mostrar ventosas
distais; não está claro se os braços alongados suportam ventosas
distalmente (Fig. 4d ).
As ventosas têm geralmente entre ~0,31 e ~0,62 mm de diâmetro. As
ventosas parecem ter sido bisseriadas onde quer que estejam presentes,
mas ambas as fileiras nem sempre são preservadas ao longo de todo o
comprimento do braço. As fileiras de ventosas são separadas lateralmente
por ~0,5 mm (Fig. 4e-g ). Dentro de uma fileira, as ventosas são imediatamente adjacentes proximodistalmente ou separadas por até ~0,4 mm (Fig. 4e–g ). Não há evidências de que os otários foram perseguidos.
A maioria
dos braços está incompletamente preservada, então é possível que o
alongamento aparente de dois dos braços seja um artefato tafonômico.
Consideramos isso improvável porque exatamente dois braços são alongados
e os braços alongados têm comprimentos aproximadamente semelhantes,
sugerindo que são do mesmo par de braços. Além disso, os braços mais
curtos mais bem preservados têm comprimentos aproximadamente
semelhantes. A probabilidade sugere que um artefato de preservação
resultaria em um número diferente de dois de braços desigualmente
alongados associados a braços encurtados de comprimentos diferentes.
Espécimes fósseis adicionais serão necessários para testar essa
hipótese. Com base na afinidade filogenética de Syllipsimopodi ,
nós os interpretamos como par de braços II. No entanto, identidades de
armas precisas não podem ser determinadas; isso é uma inferência e não
uma observação. Syllipsimopodi bideni é
codificado de forma conservadora desconhecida na análise filogenética
para todos os caracteres relacionados à modificação/eliminação do par de
braços II (Dados Suplementares 1 ).
Um possível funil medindo ~2,4 mm de comprimento é preservado na borda lateral da cabeça (Fig. 3 ). Uma identificação de funil é defendida porque ventosas parecem estar ausentes na estrutura (Fig. 4d ), e não podemos conectá-lo facilmente a um dos dez braços identificados (Fig. 4b, c ).
O aparelho
bucal é preservado como uma mancha retangular escura dentro de uma
mancha circular clara que é distinta dos tecidos circundantes do
braço/cabeça (Fig. 3 Suplementar 5 ).
A estrutura escura (~3,5 × ~1,3 mm) parece ser texturalmente distinta
dos outros tecidos preservados, sugerindo que poderia ser um
remanescente do bico. Uma intrigante banda em forma de S medindo ~0,33
mm de largura é preservada dentro do aparelho bucal. É possível que esta
banda seja um remanescente da rádula, mas suspeitamos que seja mais
provável que seja um braço com ventosa sobreposta por causa do
comprimento relativamente longo (em comparação com a mancha retangular
escura) e as aparentes formas circulares que revestem as partes da
banda.
Uma
estrutura escura, contígua, anteroposteriormente alongada, semelhante a
um saco, é preservada lateralmente deslocada da crista mediana central
do gládio. Interpretamos isso como o saco de tinta. O saco de tinta mede
~2,6 cm de comprimento anteroposterior e ~0,3 cm de largura
lateralmente no ponto mais largo (Figs. 3 , 4 ).
Interpretações ecológicas .
Os tecidos moles preservados e o gládio sugerem um corpo em forma de
torpedo que lembra as lulas existentes. As barbatanas parecem ter sido
grandes o suficiente para funcionar potencialmente como um suplemento
viável para natação a jato, mas sua aparente forma circular e posição
terminal parecem sugerir que um papel estabilizador pode ter sido mais
importante. Se os braços são preservados aproximadamente em seus
comprimentos totais, então um par de braços era consideravelmente mais
longo do que os outros quatro pares, que eram mais curtos (relação
braço-comprimento do corpo) do que na maioria dos octobráquios
existentes 37 .
Parece provável que os braços alongados capturassem presas para serem
confinadas e manipuladas pelos braços mais curtos, análogos aos
decabrachianos existentes. O tipo não preserva o conteúdo intestinal
identificável, então a dieta é desconhecida. Enquanto Vampyroteuthis continua
sendo o melhor análogo vivo para entender vampiropodes extintos (porque
é o vampiropode existente mais plesiomórfico), essas observações
sugerem que Syllipsimopodi pode
ter preenchido um nicho mais semelhante às lulas costeiras existentes,
ou seja, um predador nectico de nível médio. Não é inconcebível que Syllipsimopodi pode
ter usado seus braços carregados de ventosas para arrancar pequenos
amonóides de suas conchas, ou se aventurar mais na costa para extrair de
forma semelhante pequenos braquiópodes, bivalves e/ou conularídeos 18 , 22 ; isso é especulação embora.
Observações .
Há um desacordo considerável sobre a terminologia adequada para o grupo
que combina vampiromorfos, octopods e seus ancestrais - Octobrachia,
Octopodiformes e Vampyropoda são os nomes mais comuns. O Tratado de Paleontologia de Invertebrados 7 recomenda
Octobrachia como uma superordem para todos os coleóides que perderam o
par de apêndices II ou o modificaram em filamentos (ou seja,
Prototeuthidina, Loligosepiina, Teudopseina, Vampyromorphida e
Octopoda). Seguimos esta recomendação (Fig. 1 )
e, portanto, consideramos Octobrachia como um táxon definido por
apomorfia. Sugerimos que o nome Octopodiformes seja mantido para o grupo
coroa (Fig. 1 ),
uma vez que este parece ser o termo mais popular na literatura
neontológica. O nome Vampyropoda, que é popular entre os paleontólogos e
foi formalmente classificado acima de Octobrachia 38 , deve ser mantido para o grupo total (Fig. 1 ).
Isso mantém os três termos mais comuns precisos como normalmente usados
na literatura e fornece nomes de clado necessários para maior
precisão nomenclatural, sem inventar termos adicionais que confundiriam
ainda mais a discussão.
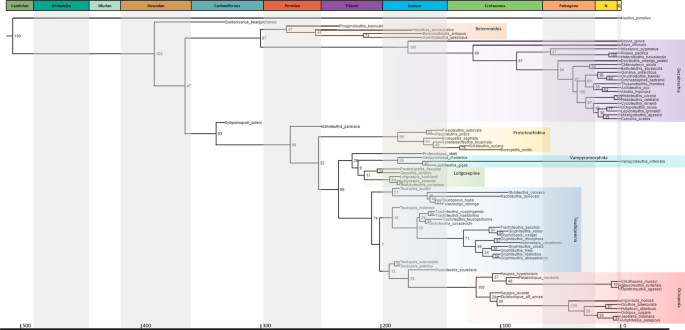
A análise Bayesiana FBD (Fossilized Birth-Death) reconstrói o Syllipsimopodi bideni gen. et sp. novembro como o primeiro vampiropode divergente; o nó é bem suportado com uma probabilidade posterior de 93% (Fig. 6 ).
Esta colocação é suportada pela perda do fragmocone, perda do rostro
primordial, presença de uma interrupção ventral mediana no
gládio/proóstraco e concha dorsal (Fig. 8 , Notas Suplementares ). do Triássico Idahoteuthis , que havia sido descrito como uma possível lula miopsídeo 39 , também é recuperado como um vampiropode inicial; o nó tem uma probabilidade posterior de 99% (Fig. 6 ). Além dos caracteres compartilhados com Syllipsimopodi , a posição de Idahoteuthis é sustentada pela presença de campos laterais e pela forma do campo mediano anterior (Fig. 8 , Notas Suplementares ). A prototeutidina é recuperada como o primeiro clado octobraquiano divergente (Figs. 1 , 6 ), ao contrário das análises de parcimônia anteriores, que reconstruíram os prototeutídeos como loligosepiídeos derivados 1 , 2 ou octopods do caule 2 .
Esses resultados concordam melhor com argumentos não cladísticos
recentes e com o registro estratigráfico – os mais antigos octobráquios
conhecidos são os prototeutídeos do Triássico Germanoteuthis e Reitneriteuthis 7 . Em vez do octobrachiano 1 Jurássico Proteroctopus é recuperado como o vampiromorfo do caule mais basal (Figs. 1 , 6 ). Loligosepiina é recuperada como um clado irmão de Vampyromorphida (Figs. 1 , 6 ); cladogramas anteriores reconstruíram os loligosepiids como um grau de octopodiformes de caule 1 ou um grau de octopods de caule 2 . Vampyronassa e Leptoteuthis (anteriormente um loligosepiid 7 mas nunca cladisticamente posicionado dentro do grupo 1 , 2 ) são recuperados como vampiromorfos (Fig. 6 ).
Fig.
6: Filogenia morfológica Bayesiana datada de FBD (Fossilized
Birth-Death) de cefalópodes neocoleóides, mostrando a posição de Syllipsimopodi bideni gen. et sp. novembro
Os
números nos nós indicam probabilidades posteriores (porcentagem). Dicas
datadas da primeira aparição do membro mais velho da linhagem relevante
no registro fóssil (ver “Métodos” e Informações Complementares ). Mostrando escala de tempo geológico datado usando Gradstein et al. 2 ,
datas em Ma e cores da Comissão Internacional de Estratigrafia; Q =
Quaternário (amarelo pálido). Táxons importantes destacados: laranja =
Belemnoidea, roxo = Decabrachia, amarelo = Prototeuthidina, verde =
Loligosepiina, ciano = Vampyromorphida, azul = grau 'teudopseid',
vermelho = Octopoda. A árvore não apresenta designações taxonômicas
revisadas: Teudopsis bollensis = Briggsiteuthis bollensis gen. e pente. nov., Teudopsis jeletzkyi = Fuchsiteuthis jeletzkyi gen. e pente. nov., Teudopsis subcostata = Suttoniteuthis subcostata gen. e pente. nov., Glyphiteuthis rhinophora = Justinianiteuthis rhinophora gen. e pente. nov., Glyphiteuthis minor = Fisheriteuthis minor gen. e pente. nov., e Trachyteuthis bacchiai = Edmunditeuthis bacchiai geração
e pente. novembro Árvore extraída do arquivo de saída MrBayes TRE
usando icytree.org. Os dados de origem são fornecidos como um dados de origem .
Teudopseina é recuperada como um grau parafilético de octopods de caule (Figs. 1 , 6 ). Os cladogramas anteriores os reconstruíram como vampiromorfos de grupo total 2 , ou como uma combinação de octopodiformes de caule e octopods de caule 1 ; nenhuma filogenia recuperou uma Teudopseina monofilética, Teudopseidae ou Teudopsis 1 , 2 , 40 . Nós estabelecemos novos gêneros para Teudopsis bollensis ( Briggsiteuthis gen. nov.), Teudopsis jeletzkyi ( Fuchsiteuthis gen. nov.) e Teudopsis subcostata ( Suttoniteuthis gen. nov.), que foram consistentemente demonstrados filogeneticamente isolados da espécie-tipo , Teudopsis bunelii 1 , 2 , 38 (Fig. 6 ). T. jeletzkyi e T. subcostata não são atribuídos ao mesmo gênero porque seu clado (Fig. não é encontrado em outras filogenias 1,2 . 6 )
Teudopseina poderia ser mantida como um grupo monofilético de garupa,
restringindo a subordem a Teudopseidae e Palaeololiginidae (Fig. 6 ) - este clado foi recuperado em todas as filogenias 1 , 2 . Todas as análises também sugerem que Teudopseidae é totalmente (sem grupo de garupa) parafilético 1 , 2 ou polifilética (Fig. 6 ) em relação a Palaeololiginidae, que possui antiguidade nomenclatural 7 ,
tornando Teudopseidae um sinônimo júnior. A subordem de alcatra
Teudopseina consistiria, assim, em uma família, Paleololiginidae; porém,
declinamos a revisão subordinada até que as inter-relações sejam mais
estáveis. Ao contrário de estudos anteriores 1 , 2 , recuperamos um Trachyteuthidae monofilético (Fig. 6 ). Estabelecemos novos gêneros para os traquiteutídeos Glyphiteuthis rhinophora ( Justinianiteuthis gen. nov.), Glyphiteuthis minor ( Fisheriteuthis gen. nov.) e Trachyteuthis bacchiai ( Edmunditeuthis gen. nov.) porque cada espécie foi consistentemente encontrada isolada de seu respectivo gênero 1 , 2 (Fig. 6 ). As duas últimas espécies não foram reatribuídas a Glyphidopsis porque esse clado (Fig. 6 ) não foi recuperado em outras filogenias 1 , 2 . A Discussão Suplementar inclui detalhes taxonômicos dos novos gêneros.
Recuperamos a monofilia de belemnoides (Fig. 6 ), como em análises anteriores 1 , 2 ; no entanto, nossos resultados colocam Belemnoidea irmã de Decabrachia (Fig. 1 ) ao invés de Decabrachia, irmã de Sepiida (chocos) e Sepiolida (lulas bobtail) 1 , 2 .
Nossa topologia concorda melhor com o registro estratigráfico e as
estimativas de tempo de divergência molecular. O choco fóssil definitivo
mais antigo é o Maastrichtiano Ceratisepia 41 mas o belemnóide definitivo mais antigo é o fragmoteutídeo Changhsingian Permoteuthis 42 . Tanto a nossa árvore morfológica FBD (Fig. 6 ) quanto as estimativas do relógio molecular sugerem que os chocos se separaram da Spirula no Jurássico 43 . Para que os belemnoides fossem incluídos em Decabrachia, a divisão sepiid-spirulid precisaria ocorrer no Paleozóico.
Os
fragmoteutídeos há muito são vistos como um provável precursor dos
coleóides portadores de gládios e, portanto, dos vampiropodes 44 , 45 , 46 , mas essa narrativa foi rejeitada por todas as análises cladísticas 1 , 2 (Fig. 6 ).
Em vez disso, os fragmoteutídeos se agrupam consistentemente com os
outros belemnoides e os belemnoides com os decabrachians. No entanto, a
ideia permaneceu proeminente na literatura 4 , 42 ,
em parte porque o proóstraco fragmoteutídeo tripartido parece uma
reminiscência de proóstraca vampirópode. Isso é explicável se os campos
laterais forem uma simplesiomorfia neocoleóide - o que significa que os
campos laterais foram perdidos exclusivamente por Syllipsimopodi ou não foram bem preservados no Syllipsimopodi (não
pudemos determinar conclusivamente a presença/ausência).
Alternativamente, os campos laterais de vampiropodes podem ter se
desenvolvido independentemente de estruturas comparáveis em
belemnoides e decabraquios (Fig. 6 ). Análises cladísticas anteriores 1 , 2 não
testaram os argumentos estratigráficos para as origens dos
fragmoteutídeos. Phragmoteuthids originou-se no último Permiano
(Changhsingian) 42 , e os vampiropodes mais antigos já eram conhecidos desde o Triássico Médio (ladiniano) 7 ;
um cronograma confortável para a divergência. No entanto, nossa
filogenia FBD incorpora explicitamente a estratigrafia e ainda rejeita
uma origem de fragmoteutídeos para Vampyropoda. Isso não é
surpreendente, pois Syllipsimopodi antecede
todos os fragmoteutídeos conhecidos em quase 70 milhões de anos, e já
possuía um gládio dorsal sem fragmocone ou rostro primordial.
Spirulida é aqui recuperada como a ordem mais basal decabraquiana (Fig. 6 ),
mas isso pode ser influenciado por sua condição de concha
plesiomórfica. No entanto, os resultados parecem substancialmente mais
plausíveis do que os cladogramas anteriores incluindo fósseis, que
reconstruíram os espirulídeos profundamente aninhados dentro de
Decabrachia 1 , 2 . Filogenias moleculares produziram resultados conflitantes para a posição de Spirula , sugerindo colocação dentro de Bathyteuthida 47 ou como irmã de Myopsida 48 , Oegopsida 9 , 49 , Bathyteuthida + Oegopsida 50 ou Sepiida 51 .
Recuperamos Sepiida como a próxima ordem divergente mais antiga, e os
teutídeos (Myopsida, Bathyteuthida e Oegopsida) são recuperados em um
clado irmão de Idiosepiida + Sepiolida (Fig. 6 ).
No entanto, os decabrachianos não foram o foco desta análise, e nossa
amostragem limitada não pretende ou espera resolver significativamente
as inter-relações dentro desse grupo.
Discussão
A
análise filogenética colocou a origem de Neocoleoidea entre 397,8 e
334,8 Ma, com mediana de 363,4 Ma. Isso coincide aproximadamente com os
limites Frasniano-Famenniano (371,1 Ma) e Devoniano-Carbonífero (359,3
Ma) 8 :
um intervalo dinâmico notável para a extinção em massa do Devoniano
Superior (Frasniano-Famenniano), o início da Idade do Gelo Paleozóica
Superior e a extinção em massa do final do Devoniano 8 , 52 , 53 . Coletivamente, esses eventos transformaram fundamentalmente a biosfera marinha.
A estufa do Paleozóico 54 fomentou as primeiras florestas 55 , um dos sistemas de recifes mais extensos da história da Terra 56 , o surgimento de vertebrados (ou seja, peixes) como um componente importante dos ecossistemas marinhos 57 , 58 , 59 , 60 , e a origem dos amonóides 57 , 58 , 59 , 61 , 62 .
Embora ainda não existam evidências fósseis conclusivas, é lógico que
esse ambiente também cultivou os primeiros coleóides; a divergência
entre Gordoniconus e Neocoleoidea está prevista para 431,6-355,5 Ma, com uma mediana de 389,0 Ma (Devoniano Médio, Eifeliano 8 ).
O evento Big Five final do Frasnian marcaria o desaparecimento dos
recifes de corais tabulados e estromatoporóides do meio do Paleozóico
(populações de corais igualmente extensas não se desenvolveriam por ~
200 milhões de anos) 56 e foi catastrófico para a biodiversidade global 63 , que continuou a diminuir ao longo do Famênia 64 . No entanto, com exceção das extinções entre os restantes vertebrados 'ostracodermes' sem mandíbula 57 , os táxons nadadores parecem ter sido relativamente inalterados 57 , 62 , 65 .
No rescaldo imediato da crise, a disparidade de amonóides em relação à
diversidade caiu para seus níveis mais baixos do Paleozóico, mas no
Famenniano médio a mesma métrica de disparidade atingiu seu valor mais
alto do Paleozóico. 62 .
O final do Frasnian poderia ter liberado o ecoespaço para os
neocoleóides nascentes, e eles podem ter se beneficiado das mesmas
condições que promoveram a alta disparidade de amonóides no Famennian.
O início da Idade do Gelo do Paleozóico Superior 52 , a primeira grande glaciação desde o final do Ordoviciano 54 , coincide com a extinção em massa do final do Devoniano 53 . Este evento parece ter tido menos impacto direto nas comunidades bentônicas 56 , 64 , mas precipitou uma grande reorganização da coluna d'água 57 , 65 .
Vertebrados 'Placodermes', amonóides clymeniids e tentaculitoids
homoctenid foram extintos; a diversidade de nautilóides discosorídeos e
oncocerídeos, bem como de vertebrados 'acantodianos' e sarcopterígios,
foi bastante diminuída 57 , 65 .
Esses grupos impactados negativamente são geralmente caracterizados
como mais fortemente blindados, menos hidrodinâmicos e mais lentos; em
contraste, os táxons que proliferaram nos oceanos do Mississippian (por
exemplo, vertebrados eucondrichthyan e actinopterygian, amonóides
prolecanitídeos) eram geralmente menos sobrecarregados e mais ágeis 57 , 65 . Nossa origem prevista de Neocoleoidea se encaixaria perfeitamente nesse padrão de reorganização marinha e nektonização.
O Bear Gulch lagerstätte preserva uma fatia do Carbonífero após outra crise, a extinção em massa de meados de Serpukhovian 53 , 57 .
Este evento pode ser mais conhecido por concluir a era Mississipiana
dos crinóides, mas os táxons pelágicos também foram impactados.
Amonóides, conodontes e condrictios, em particular, enfrentaram taxas de
extinção elevadas 53 . A disparidade amonóide se contraiu precipitadamente em relação à diversidade e não se recuperaria até o Moscovo 66 . Talvez as origens dos vampiropodes estivessem ligadas a esses nichos vagos; no entanto, a divergência de Syllipsimopodi é
estimada para o Tournaisian/Visean (354,4–330,3 Ma, mediana 337,5 Ma),
tornando este cenário improvável. De maneira menos controversa, pode-se
argumentar que as características exibidas por Syllipsimopodi contribuíram
para a sobrevivência de vampiropodes em várias extinções em massa
(meados de Serpukhovian, Guadalupian, final-Permiano e Triássico
Superior), até sua proliferação no Jurássico 7 .
Ao
internalizar o fragmocone, os coleóides poderiam adicionar nadadeiras
para complementar a natação a jato energeticamente cara. Além disso, os
coleóides escaparam das restrições inerentes impostas pela câmara do
corpo sobre o tamanho máximo da cabeça, aparelho bucal, funil e
apêndices, e o novo proóstraco fornece uma placa grande e leve para
ancorar esse anterior expandido. A expansão do anterior aumenta o peso
anterior, mas o fragmocone posterior permanece cheio de gás flutuante,
de modo que o animal pode ser forçado a uma orientação vertical
descendente em vez da presumivelmente preferível horizontal (embora
observações de Spirula possam desafiar essa sabedoria convencional 67 ) . Os primeiros coleóides, como Gordoniconus ,
parecem resolver esse problema encapsulando o fragmocone posteriormente
em um rostro primordial mineralizado que pode funcionar como um
contrapeso 13 (com
o benefício adicional de aumentar o momento de deslizamento após cada
pulso do jato). Belemnoids elaboraram sobre isso com rostra e epirostra
mais pesadas, e decabrachians extintos, como Belopterina 68 e Anomalosaepia 69 ,
encapsularam seus fragmocones em bainhas elaboradas e mineralizadas. Os
vampiropodes, no entanto, adotaram uma abordagem diferente – a
flutuabilidade neutra foi mantida através da composição iônica e do
conteúdo de água do mar de seus tecidos 6 ,
em vez do conteúdo gasoso de um fragmocone posterior. Assim, evitando o
problema de orientação inteiramente. Os vampiropodes parecem ter
enfrentado pressão seletiva para eliminar o fragmocone não funcional,
apesar dos benefícios potenciais do momento de deslizamento. Isso teria
diminuído as despesas metabólicas, reduzido o peso e liberado espaço
adicional para os tecidos moles abaixo da porção posterior da camada
proostracal da concha, que agora é um gládio.
O gládio é considerado um derivado do proóstraco, uma projeção anterodorsal da concha interna do coleóide 14 . Uma hipótese amplamente aceita sustenta que o proóstraco é um resquício da superfície dorsal da câmara do corpo da concha 14 .
No entanto, foi demonstrado que o proóstraco belemnóide é uma camada
lamelo-orgânica que se estende e encapsula o ápice do fragmocone; esta
camada está situada entre a conoteca interna mineralizada e o rostro
primordial externo mineralizado 70 .
Portanto, (1) o proóstraco provavelmente nunca foi mineralizado e
sempre de composição semelhante ao gladii derivado, (2) o proóstraco
evidentemente não é composto de tecidos conotecais (ou seja, camada
prismática externa, camada nacarada, camada prismática interna) e,
portanto, não pode ser formado através da perda parcial da conoteca, e
(3) o proóstraco não está restrito à câmara do corpo, então é improvável
que seja um remanescente dele. Essa reavaliação da homologia
proostracal também implica que o cone não é homólogo ou remanescente do
fragmocone. Em vez disso, o cone é a porção do ápice da concha composta
por tecidos prostracais; o cone cobriria o ápice do fragmocone se um
fragmocone estivesse presente (Fig. 5 ).
Isso explicaria a ausência de um sifúnculo e septos no cone dos
vampiropodes – os vampiropodes perderam completamente um fragmocone e
retêm apenas a camada proostracal sobrejacente. Esse tecido de cobertura
nunca revestiu o sifúnculo ou septo interno, portanto, nenhum
remanescente dessas estruturas seria esperado. Em contraste, muitos
decabráquios retêm um fragmocone não mineralizado como uma camada
distinta dentro do cone (e abaixo do gládio), completa com sifúnculo e
septos 71 .
Portanto, parece improvável que o proóstraco seja um vestígio da câmara
do corpo. Em vez disso, o proóstraco parece mais provavelmente ser uma
extensão anterior da camada proostracal, que encapsulava ancestralmente a
concha coleóide. Essa camada proostracal pode ser homóloga ao
perióstraco náutilo 14 .
Syllipsimopodi é o mais antigo vampiropode conhecido e gládio com cefalópodes por ~81,9 milhões de anos 7 , 8 , como tal, fornece uma visão incomparável sobre as origens do vampiropode bauplan. O campo mediano de Syllipsimopodi assemelha-se
a um triângulo agudo que se abre anteriormente a partir do ápice e
termina em uma borda anterior plana; nenhuma zona hiperbolar está
presente e os campos laterais estão ausentes ou pequenos. O cone é
pequeno e restrito posteriormente. Este simples gládio lembra o
proóstraco do caule coleóide Gordoniconus , que parece ser uma simples continuação anterior da concha passada pelo fragmocone. O gládio de Syllipsimopodi também
se assemelha aos prototeutídeos, exceto que as bordas laterais do campo
mediano do prototeutídeo são reforçadas (uma sinapomorfia da subordem),
e os prototeutídeos têm campos laterais inequívocos (embora simples)
próximos ao ápice da concha. Loligosepiídeos e vampiromorfos adicionam
uma borda anterior curva simples ao campo mediano, campos laterais mais
alongados e zonas hiperbolares proeminentes; entre esses táxons, o cone
em forma de funil que lembra um ápice de fragmocone também é substituído
pelo cone em forma de taça. Os teudopseídeos elaboram ainda mais essa
estrutura com uma borda anterior distintamente pontiaguda do campo
mediano e campos laterais e zonas hiperbolares mais complexos.
Uma trajetória evolutiva pode ser observada entre os vampiropodes. (1) Coleoides do caule ( Gordoniconus )
alongaram a camada proostracal anterior à câmara do corpo, formando o
proóstraco. (2) No final do Mississipiano, os vampiropodes ( Syllipsimopodi )
formaram um gládio através da eliminação do fragmocone subjacente
mineralizado e do rostro primordial sobrejacente. Dois apêndices (par de
braço II) podem ter sido alongados, convergindo para os tentáculos de
decabraquios posteriores (par de braço IV), mas sem dúvida todos os dez
braços permaneceram robustos. (3) No início do Triássico, campos
laterais notáveis foram adicionados ao gládio ( Idahoteuthis ).
(4) O par de braços II, que pode já ter sido isolado no desenvolvimento
dos outros pares de braços, foi reduzido a um filamento vestigial em
octobráquios. (5) Octopodiformes expandiram a zona hiperbolar entre os
campos mediano e lateral; o cone assumiu a forma de taça. Os
teudopseídeos muensterelóides abriram totalmente o cone, formando uma
patela. (6) Os octopods eliminaram o campo mediano, formando suportes de
barbatanas (cirrates) ou estiletes (incirrates); o par de braços II foi
perdido.
Excluindo
os octopods, o padrão de evolução vampyropod após a origem parece ser
de adição e não de subtração. Estruturas são adicionadas ao proóstracum
que são então expandidas e elaboradas por meio de controles de
desenvolvimento cada vez mais complexos; não há evidência de uma
redução evolutiva gradual de extensos campos laterais semelhantes a
fragmoteutídeos ou de um cone alongado anteriormente. Se o gládio é um
remanescente da câmara do corpo, então as reduções ventrolaterais devem
ter ocorrido rapidamente. Alternativamente, o vampyropod gladius se
formou através de uma simples mutação que desativou os processos de
desenvolvimento responsáveis pela formação das camadas de conchas
biomineralizadas. A estrutura restante teria sido uma simples placa
dorsal que se enrolava posteriormente ao redor do ápice (isto é, o
proóstraco); nenhuma sequência complexa de reduções graduais é
necessária. Sugerimos que este último cenário seja um modelo muito mais
plausível para a formação do gládio. Mesmo que a evidência fosse
equívoca, o cenário que propomos requer menos alterações genéticas (1
alteração = perda de todos os tecidos mineralizados) do que o modelo
convencional (7 alterações = perda da câmara ventral do corpo, perda da
câmara lateral do corpo, perda do rostro primordial, perda dos septos,
perda do sifúnculo, encurtamento do cone, desmineralização). No
entanto, a evidência não é ambígua: nenhuma filogenia sugeriu
vampiropodes derivados de um ancestral semelhante a fragmoteutídeos.
Além disso, Syllipsimopodi é anterior a todos os neocoleóides conhecidos e já possui um gládio totalmente desenvolvido sem fragmocone. Syllipsimopodi e
nossa filogenia demonstram que os vampiropodes provavelmente se
originaram logo após a origem dos neocoleóides. A rápida formação do
gládio pode ser mais facilmente explicada pela modificação de um
processo fundamental de desenvolvimento do que pela perda gradual de uma
série de estruturas complexas.
Syllipsimopodi bideni gen.
et sp. novembro é um vampyropod carbonífero com gládio com dez braços
robustos com ventosas. É o mais antigo vampyropod e neocoleoid
conhecido. Os neocoleóides originaram-se antes do Serpukhoviano;
provavelmente no final do Devoniano, potencialmente como um componente
das reorganizações pelágicas mais amplas do Famenniano ao Tournaisiano.
Nossa filogenia Bayesiana FBD reconstrói Syllipsimopodi como o vampiropódio mais basal, Idahoteuthis como um vampirópode, Prototeuthidina como o clado octobrachiano mais basal, Proteroctopus como
o vampiromorfo do caule mais basal, Loligosepiina como um clado de
vampiromorfos do caule, Teudopseina como um grau parafilético de
octopods do caule, Phragmoteuthis como
um belemnoid, e uma Belemnoidea monofilética como irmã de Decabrachia.
Pelo Serpukhovian, os vampiropodes já possuíam um gládio totalmente
formado sem um fragmocone. Isso, juntamente com a rejeição reafirmada da
hipótese das origens dos fragmoteutídeos, sugere que é improvável que o
proóstraco/gládio seja um remanescente da câmara do corpo do
fragmocone.
O
modelo convencional para a formação do gládio poderia ser plausível se
os vampiropodes se originaram muito depois das origens devonianas de
Neocoleoidea, se tivéssemos uma sequência de fósseis mostrando reduções
graduais da câmara do corpo e do fragmocone, se os primeiros proóstracos
fossem mineralizados, se os vampiropodes exibissem uma tendência geral
para a redução do gládio, ou se os vampiropodes descenderam dos
fragmoteutídeos. No entanto, cada um deles carece ou é contrariado por
evidências. Syllipsimopodi demonstra
que os vampiropodes existiam no Mississipiano e já possuíam um gládio
desmineralizado sem qualquer remanescente discernível do fragmocone. Syllipsimopodi mostra
que os primeiros gladii eram estruturas simples que foram elaboradas
por linhagens sucessivas; não conchas complexas a serem reduzidas
evolutivamente a um mínimo essencial. Por fim, nenhum fóssil foi
descoberto que possa indicar uma redução gradual da câmara do corpo,
exceto para fragmoteutídeos, e nossa análise filogenética reafirma o
crescente consenso de que os fragmoteutídeos não são
vampiropodes-tronco.
Métodos
Este
estudo está em conformidade com todos os regulamentos éticos
relevantes; nenhuma aprovação foi necessária de qualquer
conselho/comitê superior.
Análise filogenética
Redesenhamos a matriz morfológica de caracteres-táxons de Sutton et al. 2 (expandido por Kruta et al. 1 )
sob uma estrutura de codificação de contingência mais extensa e
explícita; adicionando caracteres e excluindo caracteres sobrepostos
conforme necessário. Todos os caracteres são detalhados nos Métodos Suplementares . Adicionamos Syllipsimopodi bideni (ROMIP 64897), Gordoniconus beargulchensis 13 (AMNH 43264, 50267), Idahoteuthis parisiana 39 e Acanthoteuthis speciosus 72 . Removemos os teudopseídeos do Cretáceo Superior Marekites vinarensis e Eoteuthoides caudata porque
ambos são preservados apenas como um cone fragmentário, fornecendo
muito poucos caracteres filogeneticamente informativos 7 . Também removemos as espécies carboníferas Jeletzkya douglassae e Pohlsepia mazonensis . Todos os táxons incluídos estão detalhados nos Métodos Suplementares .
Jeletzkya foi
removido porque quase nenhum caractere podia ser codificado com
confiança. Ele só foi descrito com base em uma radiografia mostrando uma
estrutura vagamente espatulada e apêndices com ganchos anexados 73 . O fóssil não foi preparado para fora de sua concreção 73 ou
escaneado por micro-CT, e ainda nunca foi claramente observado ou
redecrito. A estrutura espatulada foi interpretada como um gládio, mas
isso não foi baseado em nenhum caráter claramente diagnóstico e uma
identificação alternativa como um fragmocone, rostro ou alguma outra
estrutura não pode ser descartada. Jeletzkya é
descrito como tendo dez braços, mas os autores não indicaram onde cada
braço está na radiografia; só podemos distinguir 4 manchas vagamente
retangulares pouco distintas da estática 73 .
Assim, os únicos caracteres que poderiam ser codificados de forma
confiável seriam 'presença de uma concha' e 'presença de ganchos de
braço'. Os ganchos podem ser diagnósticos para belemnoids, mas o fóssil é
muito mal compreendido para atribuição a qualquer táxon de cefalópode
em particular, sem falar na inclusão em uma análise filogenética.
Exclusão de Jeletzkya tem um impacto mínimo nesta investigação, pois os belemnoides não são o foco principal.
O fóssil Pohlsepia foi inicialmente descrito como um octopod cirrato 10 , no entanto, é improvável que seja um cirrato, vampiropode, cefalópode ou molusco. Pohlsepia é muito anterior aos mais antigos octopods conhecidos (Cretáceo Superior) 7 , estimativas de tempo de divergência molecular para a divisão cirrato-incirrato (Jurássico) 9 e todos os outros octobrachians aparentemente ancestrais, exceto Syllipsimopodi . Pohlsepia carece de um gládio (ou vestígio de gládio), fragmocone ou rostro primordial 10 – uma combinação altamente improvável em um cefalópode carbonífero. Os apêndices propostos 10 carecem
de ganchos, ventosas, cirros, uma teia de braço e a característica
contagem de braços de 8/10; não há bico, saco de tinta inequívoco, nem
rádula. As aletas sugeridas 10 poderiam
alternativamente ser interpretadas como dobras de tecido
tafonomicamente deformadas. Os únicos caracteres conhecidos com
confiança, o contorno do corpo bulboso e a presença de apêndices, não
são diagnósticos de cefalópodes; uma afinidade cnidária pode ser mais
provável. Por carecer de quase todas as sinapomorfias de coleóides e
cefalópodes, filogenias passadas o recuperaram na base da árvore. 1 , 2 .
Isso cria efetivamente um segundo grupo externo enganoso que influencia
a polaridade do caráter em toda a filogenia. Até que uma redescrição de
Pohlsepia possa determinar com segurança sua afinidade, o gênero não deve ser incluído em nenhum estudo de inter-relações de cefalópodes.
A
matriz final é composta por 79 táxons e 153 caracteres morfológicos. As
datas de ponta referem-se à primeira aparição (FAD) de cada gênero,
agrupadas por idade e datadas usando Gradstein et al. 8 ,
com duas exceções. (1) Os FADs de espécies foram usados se várias
espécies do mesmo gênero foram codificadas na análise. (2) A
ordem/família FAD foi usada para cada ordem/família com apenas uma
espécie incluída, ou seja, Phragmoteuthida (FAD de Permoteuthis ) 42 , Belemnitida (FAD de Sichuanobelus ) 74 , Spirulida (FAD de Kostromateuthis ) 75 , Sepiida (FAD de Ceratisepia ) 41 , e Argonautidae ( FAD de Obinautilus ) 7 .
O grupo externo, Nautilus , foi datado do Darriwilian (Ordoviciano Médio) baseado em Centrocyrtoceras 76 .
Tem sido argumentado com base em estimativas de relógio molecular que
nautilids divergiram de coleóides perto do limite Siluriano-Devoniano 11 ;
no entanto, nenhum fóssil de cefalópodes foi usado para calibrar este
relógio, e a margem de erro para a divisão nautilídeo-coleóide é de ± 60
milhões de anos 11 . Os nautilídeos mais antigos são os lechritrochoceratids Silurianos (Wenlock) 76 , 77 . do Ordoviciano Médio Centrocyrtoceras 76 . Essa narrativa, de que os tarphycerids são nautilids do caule 76 , foi amplamente suplantada pelo argumento de que os nautilids se originaram de um ancestral ortoceratoid desconhecido 11 , 33 .
No entanto, a hipótese tarphycerid foi recentemente revivida e
expandida como parte de um esforço maior para reclassificar as várias
ordens de nautilóides fósseis 78 .
Neste cenário revivido, nautilídeos descendem de tarphycerids,
tarphycerids descendem de oncocerids e oncocerids descendem de
ellesmerocerids 78 — uma modernização de várias propostas muito mais antigas de inter-relações nautilóides 79 , 80 , 81 , 82 .
Nenhuma hipótese de origens nautilid ainda foi testada
filogeneticamente. Preferimos a hipótese do tarphycerid porque ela
delineia uma sequência fóssil testável específica 76 . Em contraste, nenhum fóssil ortoceratoide foi proposto como possível nautilídeo do caule. Usamos conservadoramente o FAD de Centrocyrtoceras (em oposição ao FAD de Ellesmerocerida) até o momento Nautilus na análise FBD, porque nenhuma sequência específica foi proposta ligando tarphycerids a oncocerids 76 , 78 . Centrocyrtoceras permanece dentro da margem de erro do relógio molecular para a divisão nautilídeo-coleóide 11 , 76 .
A
matriz caracter-táxon foi inicialmente composta como um arquivo NEXUS
no Mesquite 3.61. A análise Bayesiana com data de ponta foi realizada em
MrBayes 3.2.7 sob o processo de nascimento-morte fossilizada (FBD).
Usamos o modelo de clock de taxas de gama independentes (IGR) e uma taxa
de clock de distribuição normal anterior com uma média de 0,0025 e um
desvio padrão de 0,1 alterações por milhão de anos. A idade da árvore
foi fixada em um mínimo de 486,9 Ma - a base do Ordoviciano 8 e a primeira aparição de Bactroceras 83 , que pode ser o mais antigo coleóide de caule conhecido 78 . A média da árvore foi fixada em 529,0 Ma, a base do estágio cambriano 2 8 , porque o Terreneuviano superior marca o cefalópode fóssil mais antigo conhecido 84 . A análise do MCMC executou 40.000.000 simulações de 4 cadeias com um burn in de 25.000 gerações.
Atos de nomenclatura
Este
trabalho publicado e os atos nomenclaturais nele contidos foram
registrados no ZooBank, o sistema de registro online proposto para o
Código Internacional de Nomenclatura Zoológica (ICZN). Os LSIDs
(Identificadores de Ciências da Vida) do ZooBank podem ser resolvidos e
as informações associadas visualizadas por meio de qualquer navegador da
Web padrão anexando o LSID ao prefixo ' http://zoobank.org/ '. O LSID para esta publicação é: urn:lsid:zoobank.org:pub:57F16B89-1162-4261-922E-AD3B7FB54765.
Fotografia
As fotografias da Fig. 3b e da Fig. 4a–d foram
tiradas usando uma câmera Canon EOS 60D com uma lente Macro USM
EF-S60mm f/2.8 e um filtro de vidro multi-revestido digital Hoya 52 mm
Circular Polarizing Pro 1; O pacote Cognisys Stackshot 3X Macro Rail e o
Helicon Focus 6.7.1 Pro foram usados para z-stack de imagens.
Fotografias para Fig. 4e–ge Figs Suplementares. 4 – 8 foram tiradas com uma câmera Nikon D300. As imagens compostas foram costuradas usando o Adobe Photoshop 2021.
Todos os dados gerados ou analisados durante este estudo estão incluídos neste artigo publicado (e seus de Informações Complementares arquivos O arquivo de dados NEXUS utilizado neste estudo está disponível no banco de dados MorphoBank sob o código de acesso http://morphobank.org/permalink/?P4160 . A matriz morfológica do táxon de caracteres e as datas das pontas do arquivo NEXUS (Dados Suplementares 1 ) também são fornecidas nos Métodos Suplementares . O dados de origem é
a saída do MrBayes TRE. Correspondências e solicitações de materiais
devem ser endereçadas a Christopher D. Whalen em cwhalen@amnh.org. O
tipo e único espécime de Syllipsimopodi bideni gen.
et sp. novembro está depositado no Royal Ontario Museum (ROMIP 64897).
números de identificação de espécimes do museu para todos os outros
fósseis analisados diretamente estão nos Métodos Suplementares .
Nenhuma aprovação ou orientação ética foi necessária porque este estudo
analisou apenas fósseis de invertebrados em coleções de museus; nenhum
material novo foi coletado e este estudo não inclui quaisquer vestígios
arqueológicos ou espécimes recentes. dados de origem são fornecidos com este artigo.
Referências
Kruta, I. et ai. Proteroctopus ribeti na evolução dos coleóides. Paleontologia 59 , 767-773 (2016).
Fuchs, D. Problemas de homologia na morfologia dos cefalópodes: enganosas (dis)similaridades entre diferentes tipos de 'ceco'. Suíço J. Paleontol. 138 , 49-63 (2019).
Fuchs, D., Keupp, H. & Wiese, F. Protoconch morfologia de Conoteuthis (Diplobelida, Coleoidea) e suas implicações na origem presumida do Sepiida. Cretac. Res. 34 , 200-207 (2012).
Clements,
T., Colleary, C., De Baets, K. & Vinther, J. Os mecanismos de
Bouyancy limitam a preservação de tecidos moles de cefalópodes coleóides
em lagerstätten do Mesozóico. Paleontologia 60 , 1-14 (2017).
Gradstein, FM & Ogg, JG em Geologic Time Scale 2020 (eds. Gradstein, FM, Ogg, JG, Schmitz, MD & Ogg, GM) 21–32 (Elsevier, 2020).
Tanner,
AR et ai. Relógios moleculares indicam rotatividade e diversificação de
cefalópodes coleóides modernos durante a Revolução Marinha Mesozóica. Proc. R. Soc. B 284 , 20162818 (2017).
Kröger,
B., Vinther, J. & Fuchs, D. Origem e evolução dos cefalópodes: uma
imagem congruente emergente de fósseis, desenvolvimento e moléculas. 602-613 33 (2011).
Cuvier,
G. Segunda Memória sobre a organização e as relações dos animais de
sangue branco, na qual tratamos da estrutura dos moluscos e sua divisão
em ordem, lida na Sociedade de História Natural de Paris, 11 Prairial an
third. ou J. des. Sci. a partir de. Lett. a partir de. Artes 2 , 433-449 (1795).
von
Boletzky, S. Aspectos evolutivos do desenvolvimento, estilo de vida e
modo reprodutivo em octopods incirrate (Mollusca, Cephalopoda). Rev. suisse Zool. 99 , 755-770 (1992).
Grogan,
ED & Lund, R. O ambiente geológico e biológico do calcário Bear
Gulch (Mississippian de Montana, EUA) e um modelo para sua deposição. Geodiversitas 24 , 295–315 (2002).
Mickle,
KE, Lund, R. & Grogan, ED Três novos peixes paleoniscoides do
Calcário Bear Gulch (Serpukhovian, Mississippian) de Montana (EUA) e as
relações de actinopterígios inferiores. Geodiversitas 31 , 623-668 (2009).
Lund,
R., Greenfest-Allen, E. & Grogan, ED Habitat e diversidade do peixe
Bear Gulch: a vida em uma baía marinha do Mississippi de 318 milhões de
anos. Paleogeogr. Paleoclimatol. Paleoeco. 342–343 , 1–16 (2012).
Grogan,
ED, Lund, R. & Fath, M. Um novo condrichthyan petalodonte do Bear
Gulch Limestone de Montana, EUA, com reavaliação de Netsepoye hawesi e comentários sobre a morfologia de petalodontes holomórficos. Paleontol. J. 48 , 1003-1014 (2014).
Lund, R. New petalodonts (Chondrichthyes) do Alto Mississippian Bear Gulch Calcário (Namurian E2b) de Montana. J. Vertebr. Paleontol. 9 , 350-368 (1989).
Lund, R. A morfologia de Falcatus falcatus (St. John e Worthen), um chondrichthyan stethacanthid Mississipian do calcário Bear Gulch de Montana. J. Vertebr. Paleontol. 5 , 1-19 (1985).
Landman,
NH & Davis, RA Mandíbula e colheita preservada em um cefalópode
nautilóide ortocônico do calcário Bear Gulch (Mississippian, Montana). N. Mex. Bur. Mineiro de Minas. Recurso. 44 , 103-107 (1988).
Mapes, RH, Weller, EA & Doguzhaeva, LA in Cephalopods - Present and Past (Tanabe, K., Shigeta, Y., Sasaki, T. & Hirano, H. eds.) 155–170 (Tokai University Press, 2010) .
Landman, NH, Mapes, RH & Cruz, C. in Cephalopods—Present and Past (eds. Tanabe, K., Shigeta, Y., Sasaki, T. & Hirano, H.) 147–153 (Tokai University Press, 2010 ).
Grogan, ED & Lund, R. Debeerius ellefseni (fam.
nov., gen. nov., spec. nov.), um condrichthyes autodiastílico do
calcário do Mississippian Bear Gulch de Montana (EUA), as relações dos
condrichthyes e comentários sobre a evolução dos gnatostomados. J. Morphol. 243 , 219-245 (2000).
Pointon,
MA, Chew, DM, Ovtcharova, M., Sevastopulo, GD & Crowley, QG Novas
datas U-Pb de alta precisão de tufos carboníferos da Europa Ocidental;
implicações para a calibração da escala de tempo, a periodicidade dos
ciclos carboníferos tardios e a correlação estratigráfica. J. Geol. Soc. Londres. 169 , 713-721 (2012).
Klug, C., Kröger, B., Vinther, J., Fuchs, D. & De Baets, K. in Ammonoid paleobiology: from macroevolution to paleogeography 3-24 (Springer, 2015).
Sasaki, T., Shigeno, S. & Tanabe, K. in Cephalopods—Present and Past (Tanabe, K., Shigeta, Y., Sasaki, T. & Hirano, H. eds) 35–66 (Tokai University Press, 2010).
Shigeno, S. et ai. em Cephalopods—Present Past , 23–34 (Tokai University Press, 2010).
Fuchs,
D., Keupp, H. & Schweigert, G. Primeiro registro de uma coroa
completa do braço do Loligosepia do Jurássico Inferior (Cephalopoda). Palaontologische Z. 87 , 431–435 (2013).
Jereb, P., Roper, CFE, Norman, MD & Finn, JK Cephalopods
of the World, Annotated and Illustrated Catalogue of Cephalopod Species
Known to Date, Volume 3, Octopods and Vampire Squid . (Organização para Agricultura e Alimentação das Nações Unidas, 2014).
Fuchs, D. & Weis, R. Taxonomia, morfologia e filogenia de coleóides teudopseídeos do Jurássico Inferior (Cephalopoda). Ano novo B. Geol. e Paleontologia - Papers 257 , 351–366 (2010).
Doguzhaeva,
LA et al. Um gladius do Triássico Inferior associado a restos de
tecidos moles de Idaho, EUA - um cefalópode coleóide semelhante a uma
lula no início da Era Mesozóica. Acta Paleontol. Pol. 63 , 341-355 (2018).
Košťák,
M., Jagt, JWM, Speijer, RP, Stassen, P. & Steurbaut, E. New
Paleocene sepiid coleoids (Cephalopoda) do Egito: significado evolutivo e
origem do sepiid 'rostrum'. PLoS ONE 8 , e81180 (2013).
Warnke,
KM, Meyer, A., Ebner, B. & Lieb, B. Avaliando o tempo de
divergência de Spirulida e Sepiida (Cephalopoda) com base em sequências
de hemocianina. Mol. Filogeneto. Evoluir 58 , 390-394 (2011).
Sanchez,
G. et ai. Filogenia em nível de gênero de cefalópodes usando marcadores
moleculares: status atual e áreas problemáticas. PeerJ 6 , e4331 (2018).
Strugnell,
J., Norman, M., Jackson, J., Drummond, AJ & Cooper, A. Filogenia
molecular de cefalópodes coleóides (Mollusca: Cephalopoda) usando uma
abordagem multigênica; o efeito do particionamento de dados na resolução
de filogenias em uma estrutura Bayesiana. Mol. Filogeneto. Evoluir 37 , 426-441 (2005).
Lakin,
JA, Marshall, JEA, Troth, I. & Harding, IC Estufa a casa de gelo:
uma revisão bioestratigráfica das últimas glaciações do
Devoniano-Mississipiano e seus efeitos globais. Geol. Soc. Especificação Publ. 423 , 439-464 (2016).
Stein, WE et ai. do Devoniano médio Archaeopteris sinalizam uma mudança revolucionária nas primeiras florestas fósseis. atual Biol. 30 , 421-431 (2020).
Friedman,
M. & Sallan, LC Quinhentos milhões de anos de extinção e
recuperação: um levantamento fanerozóico de padrões de diversidade em
grande escala em peixes. Paleontologia 55 , 707-742 (2012).
Korn, D. & Klug, C. em Terra e Vida: Biodiversidade Global, Intervalos de Extinção e Perturbações Biogeográficas Através do Tempo (ed. Talent, JA) 491–534 (Springer Science+Business Media, 2012).
Whalen,
CD, Hull, PM & Briggs, DEG Ecomorfometria de amonóides paleozóicos
testam a disponibilidade do ecoespaço como um condutor da diversificação
morfológica. Sci. Av. 6 , eabc2365 (2020).
Fan, J. et ai. Um resumo de alta resolução da biodiversidade de invertebrados marinhos do Cambriano ao Triássico Inferior. Ciência 367 , 272-277 (2020).
Sallan,
LC & Coates, MI Extinção do fim do Devoniano e um gargalo na
evolução inicial dos vertebrados modernos com mandíbulas. Proc. Natl Acad. Sci. EUA 107 , 10131–10135 (2010).
McCoy,
VE et ai. Assinaturas químicas de tecidos moles distinguem entre
vertebrados e invertebrados do Carbonífero Mazon Creek Lagerstätte de
Illinois. Geobiologia 18 , 560-565 (2020).
Lindsay, DJ, Hunt, JC, McNeil, M., Beaman, RJ & Vecchione, M. A primeira observação in situ da lula de chifre de carneiro Spirula spirula vira o 'conhecimento comum' de cabeça para baixo. Diversidade 12 , 449 (2020).
Yancey, TE & Garvie, CL Redescrição de Anomalosaepia ( Cephalopoda: Coleoida): um sepióide com um esqueleto bimineral de calcita e aragonita. J. Paleontol. 85 , 904-915 (2011).
Doguzhaeva,
LA & Summesberger, H. Pro-ostraca de belemnoids Triássicos
(Cephalopoda) dos Alpes Calcários do Norte, com observações sobre seu
modo de preservação em um ambiente do norte de Tétis que permitiu a
carbonização de estruturas não biomineralizadas. Neues Jahrb. pele Geol. und Palaontologie - Abhandlungen 266 , 31–38 (2012).
Arkhipkin,
AI, Bizikov, VA & Fuchs, D. Vestigial phragmocone no gladius aponta
para uma origem em águas profundas de lula (Mollusca: Cephalopoda). Profundo. Res. I 61 , 109-122 (2012).
Klug,
C., Schweigert, G., Fuchs, D., Kruta, I. & Tischlinger, H.
Adaptações à natação de alta velocidade estilo lula em belemnitids
jurássicos. Biol. Lett. 12 , 20150877 (2016).
Turek, V. in Cephalopods - Present and Past , 85-92 (Tokai University Press, 2010).
King,
AH & Evans, DH Classificação de alto nível dos cefalópodes
nautilóides: uma proposta para a revisão do Tratado Parte K. Swiss J. Palaeontol. 138 , 65-85 (2019).
Teichert, C. et ai . Tratado
sobre Paleontologia de Invertebrados, Parte K, Mollusca 3:
Cephalopoda—General Features, Endoceratoidea - Actinoceratoidea -
Nautiloidea, Bactritoidea (Sociedade Geológica da América e University of Kansas Press, 1964).
Aubrechtová, M. Uma revisão do cefalópode Ordoviciano Bactrites sandbergeri Barrande: posição sistemática e paleobiogeografia de Bactroceras . Geobios 48 , 193-211 (2015).
Hildenbrand,
A., Austermann, G., Fuchs, D., Bengtson, P. & Stinnesbeck, W. Um
cefalópode potencial do início do Cambriano do leste da Terra Nova,
Canadá. Comum. Biol. 4 , 388 (2021).
Esta
pesquisa foi apoiada através do NSF Postdoctoral Research Fellowship
Program in Biology Program, Grant No. 2010822, concedido ao CDW; e as
Bolsas de Pesquisa para Estudantes da Sociedade Paleontológica, Prêmio
Elis L. Yochelson, concedido ao CDW Pelo acesso a espécimes e serviços
de museu, agradecemos a JB Caron (Museu Real de Ontário), S. Butts
(Museu Yale Peabody), J. Utrup (Museu Yale Peabody ) e B. Hussaini
(Museu Americano de História Natural). Agradecemos a DEG Briggs
(Universidade de Yale, Yale Peabody Museum), J. Vinther (Universidade de
Bristol) e RE Plotnick (Universidade de Illinois em Chicago) pelas
conversas e comentários úteis. Agradecemos a S. Thurston, MH Chase e A.
Smith (Museu Americano de História Natural) pelos serviços de
microscopia e imagem. Agradecemos a KA Whalen (Detroit Institute of
Arts) pelas reconstruções artísticas.
Informação sobre o autor
Afiliações
Departamento de Paleontologia de Invertebrados, Museu Americano de História Natural, Nova York, NY, 10024, EUA
Christopher D. Whalen e Neil H. Landman
Departamento de Ciências da Terra e Planetárias, Universidade de Yale, New Haven, CT, 06511, EUA
Christopher D. Whalen
Contribuições
A
CDW concebeu o estudo, obteve financiamento e identificou o fóssil. O
CDW editou e codificou os caracteres morfológicos e executou a análise
filogenética. CDW criou todas as figuras científicas, exceto as
reconstruções artísticas. CDW compôs o manuscrito com contribuições da
NHL. Ambos os autores aprovaram o artigo final.
Acesso Aberto Este
artigo está licenciado sob uma Licença Creative Commons Atribuição 4.0
Internacional, que permite o uso, compartilhamento, adaptação,
distribuição e reprodução em qualquer meio ou formato, desde que você dê
os devidos créditos ao(s) autor(es) original(is) e à fonte, fornecer um
link para a licença Creative Commons e indicar se foram feitas
alterações. As imagens ou outros materiais de terceiros neste artigo
estão incluídos na licença Creative Commons do artigo, a menos que
indicado de outra forma em uma linha de crédito para o material. Se o
material não estiver incluído na licença Creative Commons do artigo e
seu uso pretendido não for permitido por regulamentação legal ou exceder
o uso permitido, você precisará obter permissão diretamente do detentor
dos direitos autorais. Para visualizar uma cópia desta licença, visite http://creativecommons.org/licenses/by/4.0/ .
 =
Cambriano (verde escuro), O = Ordoviciano (azul), S = Siluriano (azul
claro), D = Devoniano (marrom), C = Carbonífero (azul), P = Permiano
(laranja vermelho), T R = Triássico (roxo) ), J = Jurássico (ciano), K = Cretáceo (verde), P G =
Paleogeno (laranja), N = Neogene (amarelo), não rotulado = Quaternário
(amarelo pálido). As setas roxas indicam nós nomeados, a barra roxa
indica o grau teudopseid. Representações artísticas criadas por K.
Whalen.
=
Cambriano (verde escuro), O = Ordoviciano (azul), S = Siluriano (azul
claro), D = Devoniano (marrom), C = Carbonífero (azul), P = Permiano
(laranja vermelho), T R = Triássico (roxo) ), J = Jurássico (ciano), K = Cretáceo (verde), P G =
Paleogeno (laranja), N = Neogene (amarelo), não rotulado = Quaternário
(amarelo pálido). As setas roxas indicam nós nomeados, a barra roxa
indica o grau teudopseid. Representações artísticas criadas por K.
Whalen. 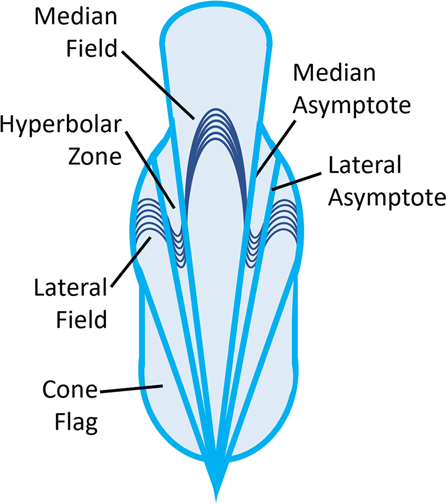


Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.