O mais antigo fóssil de mina de folhas do leste da Ásia fornece informações sobre o antigo fluxo nutricional em uma interação planta-herbívoro
Scientific Reports volume 12 , Número do artigo: 5254 ( 2022 )
Resumo
O final do Triássico viu um florescimento de interações planta-artrópodes. No final do Triássico, os insetos desenvolveram todas as estratégias distintas de herbivoria, incluindo algumas das primeiras ocorrências de mineração de folhas. Aqui, descrevemos fósseis de minas de folha excepcionalmente bem preservados em uma Cladophlebis Brongniart da Formação Momonoki, Grupo de Minas, Japão (Carniano Médio), representando as minas de folhas inequívocas mais antigas do leste da Ásia. Todas as minas exibem um rastro de excremento distinto - uma linha sinuosa contínua, que mais tarde se torna uma faixa larga contendo partículas esferoidais - demonstrando o desenvolvimento larval.
Embora as formas das trilhas de excremento sejam geralmente comparáveis às de Lepidoptera ou Coleoptera, elas não podem ser inequivocamente atribuídas a um táxon específico de mineração de folhas existente. Além disso, análises elementares por fluorescência de raios X (XRF) revelam que a trilha de excremento compreende coprólitos de fosfato. As variações quantitativas de P, S e Si entre coprólitos e nervuras foliares podem refletir processos fisiológicos (por exemplo, consumo, absorção e excreção) mediados por produtos químicos vegetais. Nossas descobertas reforçam a ideia de que a mineração de folhas se tornou uma estratégia de alimentação generalizada de insetos herbívoros no final do Triássico.
Introdução
A mineração de folhas é um meio de herbivoria pelo qual os insetos consomem folhagem viva enquanto habitam dentro do tecido da planta hospedeira 1 . A biologia da mineração foliar tem historicamente atraído muita atenção e é amplamente estudada por ecologistas 2 . Os mineiros existentes podem ser identificados entre espécies e nível de subfamília com base na forma da mina e taxonomia da planta hospedeira. Assim, as associações planta-inseto podem ser reconstruídas analisando as folhas minadas. As folhas extraídas também fornecem informações sobre grande parte da história de vida geral de um mineiro individual: ou seja, o processo de desenvolvimento, desde a oviposição, passando pelo crescimento larval e o estágio de pupa, até a emergência do adulto, é rastreável externamente, o que o torna um sistema ideal para a estudo de demografia e dinâmica populacional ao modelar as taxas e causas de mortalidade (por exemplo, parasitismo, competição) em populações naturais 3 , 4 . Como os insetos mineiros fornecem uma riqueza de informações ecológicas, eles têm sido frequentemente usados para estudar a população, a comunidade e a ecologia evolutiva das interações planta-inseto. 5 , 6 , 7 .
A evolução da mineração de folhas é um aspecto convincente do desenvolvimento de nichos alimentares de insetos. Grupos de insetos com hábitos de mineração evoluíram várias vezes em várias ordens de insetos holometábolos (por exemplo, Lepidoptera, Coleoptera, Diptera, Hymenoptera) 1 , 8 . Os mineiros são mais suscetíveis às defesas químicas das plantas hospedeiras e são mais severamente atacados por parasitoides do que os alimentadores externos 9 . Assim, eles tendem a apresentar alta especificidade de hospedeiro, com muitas espécies sendo herbívoros monófagos ou oligófagos 2 , 10 , e estão inseridos em uma rede multitrófica complexa 11 , 12 , 13 . Alguns grupos de insetos mineiros têm sido utilizados para avaliar padrões e processos de macroevolução dentro de uma estrutura filogenética 14 , 15 , 16 .
A mineração de folhas também foi examinada em estudos paleobiológicos porque trilhas de minas podem ser encontradas a partir de fósseis de impressão/compressão de folhas de plantas. Em alguns casos, o bicho-mineiro responsável pela mina é atribuído a um grupo específico de insetos comparando a forma da mina fóssil e a taxonomia da planta hospedeira com análogas existentes 17 , 18 , 19 , 20 , 21 ; esses registros podem ajudar a calibrar a filogenia de insetos no tempo. Além disso, o dano especializado observado em minas de folhas fósseis nos permite responder a várias questões relacionadas a mudanças dinâmicas ou persistência de interações planta-inseto antes e depois de perturbações ecológicas 22 , 23 , 24 , 25 .
No que diz respeito à origem evolutiva da mineração de folhas, no entanto, as informações disponíveis são escassas. A primeira ocorrência credível de mineração de folhas até agora remonta ao Triássico Médio-Final, que é muito posterior a outros grupos de alimentação funcional: a saber, esporangivoria, perfuração, alimentação de folhagem externa, predação de sementes, perfuração e sucção e galhas. 26 , 27 . Evidências substanciais de mineração de folhas são, no entanto, muito escassas nos registros anteriores ao Triássico Superior.
Aqui descrevemos um novo tipo de fóssil de mina de folhas da Formação Momonoki, sudoeste do Japão. Abaixo estão descritas as minas primorosamente preservadas em uma Cladophlebis , e os vestígios preservam seus restos orgânicos. O espécime já está exposto há anos em um armário no Museu de História e Folclore da Cidade de Minas, Yamaguchi, Japão. A localidade fóssil é datada como sendo o Carniano Médio (ca. 220 Ma) 28 e, portanto, as minas representam um dos mais antigos fósseis de minas-folha credíveis em todo o mundo. Além disso, análises elementares usando um espectrômetro de fluorescência de raios X (XRF) são realizadas nas minas fósseis, lançando uma nova luz sobre essa antiga associação planta-herbívoro da perspectiva da estequiometria ecológica.
Materiais
Cenário geológico
O Grupo de Minas, que enfrenta a China continental, está localizado no lado norte da Linha Tectônica Mediana, que divide o extremo sudoeste da ilha de Honshu, no Japão (Fig. 1 ). O cenário estratigráfico é subdividido em três estratos: as Formações Hirabara, Momonoki e Aso, em ordem crescente 29 , 30 . O material de estudo foi obtido da Formação Momonoki, que produz os fósseis de plantas e insetos mais abundantes 31 , 32 . A Formação Momonoki é datada do Carniano Médio (ca. 220 Ma) com base em dados de idade U-Pb de zircão detrítico 28 , o que é consistente com a bioestratigrafia de invertebrados marinhos e terrestres 29 , 33 . A Formação Momonoki excede 1000 m de espessura e está em conformidade com a Formação Hirabara subjacente 28 , 32 . Os arenitos e arenitos lamacentos da Formação Momonoki hospedam impressões de macrofósseis de plantas e insetos. O espécime foi coletado pelo HT em um corte de estrada na Estrada Nacional 435 enquanto a estrada estava em construção; a localidade também continha muitos fósseis de insetos 32 , 34 , 35 , 36 . A Formação Momonoki é um depósito não marinho caracterizado por depósito lacustre, deltaico, sem fósseis de invertebrados marinhos, ao contrário das Formações Hirabara e Aso 31 , 37 .
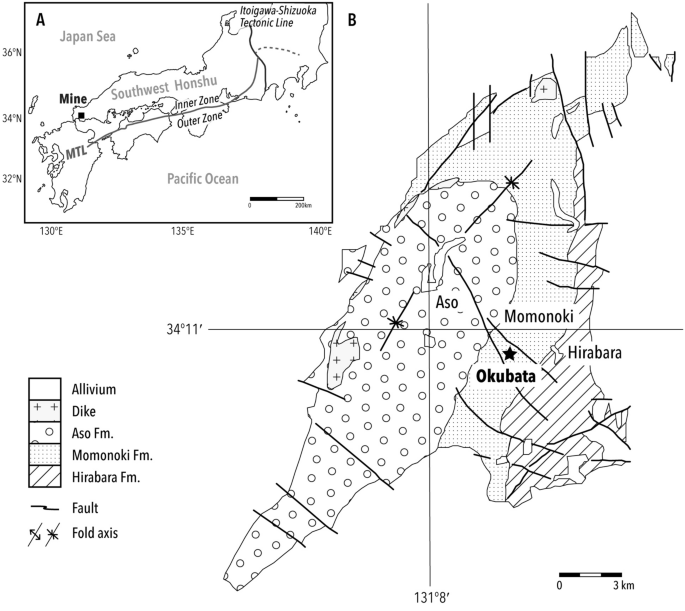
Estude a localização do local e o mapa geológico. ( A ) Mapa mostrando o sítio fóssil, Mine, Yamaguchi, no sudoeste de Honshu, Japão. A mina está localizada na Zona Interna, região formada por falhas complexas ao longo da Linha Tectônica Mediana (MTL) e separada da Zona Externa (lado sul). ( B ) Mapa geológico do Grupo Mina, ligeiramente modificado de estudos anteriores 38 , 39 . A estrela denota a localidade fóssil, Okubata.
Cenário paleobiológico
Os restos vegetais desta localidade são representados principalmente por impressões de folhagens, fragmentos de caule com folhas e, ocasionalmente, frutificações. A assembléia vegetal da Formação Momonoki é composta por samambaias, esfenófitas, cicadófitas, ginkgophytes e coníferas 40 . Existem três características da Formação Momonoki que são distintas de algumas outras assembléias de plantas fósseis coevas 41 : Sphenopsida (por exemplo, Neocalamites Halle e Equisetites Sternberg), samambaias (por exemplo, Camptopteris Presl, Clathropteris Brongniart, Dictyophyllum Lindley et Hutton, Cladophlebis ) e as coníferas (por exemplo, Podozamites Braun, Cycadocarpidium Nathorst) estão bem representadas; cicadófitas e ginkgophytes 42 são comuns; Danaeopsis Schimper e Symopteris Xu (Marattiaceae). Hepáticas ( Pallaviciniites Schuster) também estão presentes 43 . A composição floral da Formação Momonoki faz parte da Região Florística do Sul da Ásia Oriental 41 , 44 ; a região florística se estende por uma ampla faixa geográfica, desde o sul da Indonésia e norte até uma pequena área do nordeste da China, e é caracterizada pela ocorrência de Dictyophyllum e Clathropteris 41 .
Uma diversa fauna fóssil de insetos foi encontrada na Formação Momonoki 36 , 45 . Esses fósseis são oriundos de camadas estratigráficas específicas de siltitos escuros e de granulação fina, desprovidos de detritos grosseiros 31 . A maioria dos fósseis de insetos são asas isoladas, mas corpos quase completos também são encontrados 36 . Os insetos registrados abrangem pelo menos 12 ordens: Odonata, Ephemeroptera, Blattodea, Plecoptera, Paraplecoptera, Reculida, Neuroptera, Hymenoptera, Coleoptera, Hemiptera, Mecoptera, Diptera 32 . Em particular, há ricos rendimentos de asas de Coleoptera (Taldycupedidae) e Blattodea (Mancusoblattidae, Mesoblattinidae) entre a fauna de insetos deste depósito 32 , 34 . Notavelmente, a Formação Momonoki produz moscas verdadeiras 36 , embora não descritas, que foram registradas apenas em alguns depósitos do Triássico Médio-Final 46 , 47 , 48 , 49 , 50 , 51 . Embora mais de 6.000 espécimes de insetos fósseis sejam registrados nesta formação 30 , 36 , a diversidade taxonômica é amplamente inexplorada, e numerosos espécimes de insetos terrestres e aquáticos permanecem não descritos. Atualmente, 18 espécies em dez famílias pertencentes a sete ordens são descritas 32 , 33 , 34 , 35 , 45 . Acredita-se que a fauna de insetos do Grupo Mine tenha afinidade com quatro localidades remotas 32 , 33 : a Formação Mount Crosby na Austrália, a Formação Madygen no Quirguistão, a Formação Djam Djun no Vietnã e a Formação Yan-Chen na China.
Resultados
Descrição do vestígio fóssil
Três minas de folhas com estruturas semelhantes são encontradas em cinco pínulas consecutivas da pina basecopia (pontas de seta brancas na Fig. 2 A,B). Os tecidos vegetais, incluindo a epiderme e as nervuras, destacam-se nitidamente contra a matriz profundamente intemperizada da laje. As minas cobrem amplamente a lâmina, embora as margens das áreas minadas e remanescentes não sejam claramente distinguidas como tecido epidérmico rompido, indicando que os mineiros deixaram algumas camadas de tecido não consumidas. As minas se originam da extremidade proximal ou distal dos lobos. Os pontos iniciais (isto é, local de oviposição) são obscuros ou invisíveis (pontas de seta laranja na Fig. 2 C,D), situados em uma nervura foliar secundária. As trilhas de frass aparecem como fios senoidais estreitos com uma largura de 0,5 a 1,3 mm (1,1 mm em média), acompanhando muitas curvas fechadas com loops ocasionais. As trilhas de excremento expandem-se posteriormente em bandas largas de 0,5 a 0,8 mm de largura, com o acúmulo denso de material fecal; as pelotas de frass individuais são dificilmente discerníveis. As trilhas de frass são linhas únicas e contínuas, mas acompanhadas de algumas lacunas; ao todo, cada trilha de excremento cobre quase toda a parte distal de uma pínula. Embora as trajetórias das trilhas de excremento se estendam livremente através das nervuras secundárias, elas não atravessam as primeiras nervuras (isto é, nervuras centrais), exceto aquelas próximas ao ápice da folha; tais características comportamentais dão assim a toda a estrutura de mineração em uma pínula uma forma de U. Essas minas se conectam à pínula vizinha através da base alargada da pínula (Fig. 2 D-E). O fato de as nervuras foliares dentro da área minerada não estarem distorcidas determina que os feixes vasculares não sejam consumidos. O tecido escurecido no final da mina indica uma possível câmara pupal (pc, Fig. 2 D).

Fóssil de mina de folhas do Triássico tardio em Cladophlebis denticulata da Formação Momonoki, Yamaguchi, Japão. ( A ) Espécime (MMHF11-00001a), no qual minas em pínulas e marcas de mastigação são marcadas com pontas de seta brancas e asteriscos amarelos, respectivamente, e ( B ) sua contraparte (MMHF11-00001b), aproximadamente na mesma escala. ( C ) Ampliação da pina com três minas; pontas de seta laranja significam ponto de partida putativo (ou seja, local de oviposição) de minas-folha; no entanto, o ponto de partida da mina à direita é invisível. ( D ) O alargamento de duas pínulas mostra a transição da trilha de frass, sugerindo desenvolvimento larval durante a mineração, e ( E ) sua contraparte na mesma escala. As pontas de seta em laranja e azul denotam um local de oviposição (os) e uma possível câmara pupal (pc), respectivamente.
Localidade
Okubata, Omine, Mine, Yamaguchi.
Idade
Carnian, Triássico (ca. 220 Ma).
Estratigrafia
Formação Momonoki, Mine Group, prefeitura de Yamaguchi, Japão.
Material
Um único espécime de peça e contraparte (MMHF11-00001a, MMHF11-00001b; Fig. 2 A,B, respectivamente) coletado por HT da Rota Nacional 435 enquanto a estrada estava em construção (Fig. 1 ).
Planta hospedeira
A planta hospedeira é considerada Cladophlebis nebbensis (Brongniart) Nathorst com base nas seguintes características. Raque com 3 mm de largura, sulcada adaxialmente e arredondada abaxialmente. Pinna suboposta, fixada por uma base larga à raque em um ângulo de 70-75 graus, com entrenós de 34 mm; ápice desconhecido. Folhas estéreis e bipinadas. Pínulas sésseis, subopostas, surgindo em um ângulo de 71-81 graus em relação ao pavilhão craniano, catádromos em ordem; pínulas acroscópicas ligeiramente mais longas e estreitas que as basescopia; próximos, ocasionalmente sobrepostos. Pínula com cerca de 11–14 mm de comprimento e 7–8 mm de largura (área superficial ca. 91,9 mm 2 ), com base alargada e ápices obtusos; nervuras catádromas, com nervura primária distinta atingindo a margem apical e 11-13 nervuras secundárias bifurcando-se uma vez. Esta espécie pode alternativamente ser Todites fukutomii Kimura et Ohana 52 , mas esta possibilidade não é considerada aqui porque alguns caracteres distintivos (por exemplo, pínulas férteis e nervuras secundárias bifurcadas) estão ausentes em nosso espécime.
Remarks
A folhagem focal de Cladophlebis é alvo de alimentadores de folhagem externos, além de mineiros. Excisões delimitadas por bordas de reação evidentes (a área definida por dois asteriscos amarelos na Fig. 2 A) indicam sinais de alimentação externa por um folívoro mandibular.
Análises de fluorescência de raios X (XRF)
Os elementos foram quantificados em quatro pontos amostrais: (A) trilha de frass, (B) nervura foliar, (C) lâmina foliar, (D) matriz rochosa (Fig. 3 ). O espectro de XRF detectou picos de dez elementos (Al, Si, P, S, K, Ca, Ti, Fe, Sr, Zr); os outros picos foram do gás Rh utilizado para as análises. Os valores para quatro elementos (Si, P, S, Ca) foram particularmente notáveis porque esses elementos podem ser pelo menos parcialmente biogênicos e envolvidos em processos fisiológicos. A lâmina da folha era composta apenas por uma fina película de tecido vegetal e, portanto, a composição elementar pode incluir a de rocha sob a lâmina. Os resultados para a lâmina foliar geralmente mostraram uma tendência semelhante à da rocha; no entanto, P e S, presumivelmente de origem biogênica, foram detectados, confirmando que o tecido foliar estava preservado na rocha. Relatamos os resultados abaixo como a quantidade total de raios-X por segundo (cps). Variações marcadas foram encontradas em Si; foi maior na lâmina foliar (376.457 ± 5.394 cps), ligeiramente maior que na rocha (366.144 ± 5.323 cps) e menor na nervura foliar (114.689 ± 3.029 cps). O maior P foi registrado na nervura foliar (11.680 ± 1.149 cps), seguido pelo P no capim (7.923 ± 0.992 cps), enquanto os picos de P não foram detectados na lâmina ou rocha. S mostrou uma tendência semelhante a P, mas com picos no excremento (29,034 ± 1,614 cps) e na veia (11,680 ± 1,149 cps) sendo maiores do que os de P. Ca foi consistentemente detectado em todos os pontos de amostra medidos e mostrou apenas pequenas diferenças entre eles; foi maior na nervura foliar (19.380 ± 1.381 cps), seguida por frass (17.519 ± 1.286 cps), rocha (16.851 ± 1.251 cps) e lâmina foliar (13.895 ± 1.158 cps).
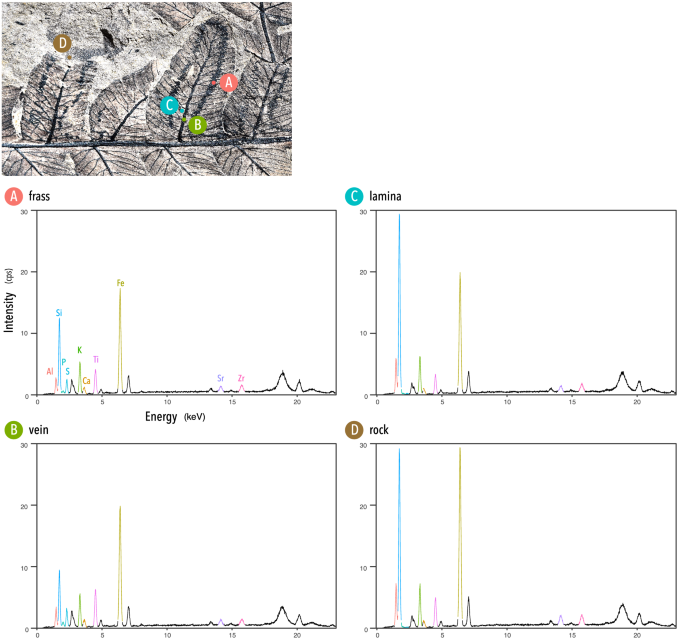
Results of X-ray fluorescence (XRF) analyses conducted with an EA6000VX High Sensitivity XRF Analyzer (Hitachi High-Tech Science Corporation). (Upper panel) Positions of the four sample points are specified on the specimen. (Lower panels) XRF spectra in the range of 0–23 keV illustrated for each sample point (A–D). Vertical axes signify cps (X-ray counts per sec).
Discussion
Significant features of the mines and the possible culprit
As Cladophlebis da Formação Momonoki são minas de folhas por insetos holometábolos e podem ser distinguidas de outros métodos de alimentação (por exemplo, alimentação de superfície) e também de características de origem tafonômica porque satisfazem mais ou menos os seguintes critérios para formas de folhas de insetos -minas 53 : um local de oviposição em uma extremidade da estrutura de mineração; uma área de oviposição alargada como uma mancha ou traço curvilíneo; um sinal do tecido foliar evacuado; a largura do tecido vegetal evacuado e da trilha de excrementos aumenta de uma extremidade à outra; a presença de um rastro de excremento, particulado ou fluidizado; tecido de resposta juntamente com o tecido marginal; um terminal distinto, como uma região expandida (câmara). Neste caso, as características das trilhas de frass são compartilhadas pelas três minas no mesmo pinna (Fig. 2 A,B), que são mais prováveis de serem produzidas pelo mesmo táxon de insetos que possui estratégias estereotipadas de consumo e excreção. Cada mina é composta por uma trilha de escória contínua e única, cuja largura se expande posteriormente no final. A mudança abrupta nas formas das trilhas fecais pode ser interpretada como desenvolvimento larval durante a mineração. Nas pínulas mineradas, os limites entre áreas mineradas e não mineradas são obscuros porque as trilhas de excremento freqüentemente cortam as nervuras das folhas secundárias sem distorcê-las. Isso pode ser interpretado de duas maneiras: as larvas podem minerar apenas células epidérmicas e não consumir mesófilos, como no caso de Phyllocnistis Zeller (Lepidoptera: Gracillariidae) 54 , 55 ; caso contrário, as larvas poderiam minerar o mesofilo evitando tecidos vasculares (por exemplo, algumas moscas agromyzid 56 ). A última estratégia é possível porque o mesofilo das samambaias é anatomicamente bem diferenciado em tecidos paliçada e esponjoso 57 .
A morfologia da mina e a variedade de plantas hospedeiras muitas vezes nos fornecem chaves para identificar insetos minadores de folhas, porque insetos minadores de folhas tendem a estar associados a uma faixa relativamente estreita de plantas e mostram padrões comportamentais estereotipados e característicos de táxons na oviposição e alimentação 2 . No entanto, em geral, a convergência entre diferentes ordens/famílias de insetos e a variação entre espécies intimamente relacionadas dificultam a diferenciação de mineradores com base em suas formas de mina 53 .
A afinidade taxonômica das minas da Formação Momonoki é aqui examinada à luz da forma da mina, antecedentes sistemáticos e evolutivos de clados de mineração de folhas, os grupos existentes de mineiros samambaia-pinnule e as origens cronológicas de possíveis culpados. A forma geral da mina não é comparável àquelas de estruturas de mineração conhecidas por mineradores de samambaias existentes 58 . Os insetos mineradores de samambaias existentes são encontrados em quatro ordens: Diptera, Coleoptera, Lepidoptera e Hymenoptera 59 .
Leaf-mining flies are diverse, encompassing three infraorders, Culicomorpha (e.g., Chironomidae), Bibinomorpha (e.g., Sciaridae), and Muscomorpha (Empidoidea–Muscoidea). Among them, Agromyzidae contain an overwhelming number of leaf-mining taxa. No evidence for the presence of dipteran leaf-mining taxa is available for the Late Triassic, although nematocerans and some of the earliest groups of brachycerans are markedly diverse46,47. Typical leaf mines of Agromyzidae, to which all fossil dipteran leaf mines have been assigned60, make linear-blotch mines61, and mine through mesophylls leaving major veins62; they tend to contain fluidized frass, which is often deposited as two discontinuous rows of pellets19. However, the forms of leaf mines by Agromyzidae greatly vary among taxa63. Flies are relatively diverse as miners of fern pinnules (and stems), represented by Chirosia Rondani (Anthomyiidae)64, Agromyzidae (e.g., Chromatomyia Hardy, Phytoliriomyza Hendel), and Cecidomyiidae59. Among them, Chirosia é um componente predominante que usa frondes ou caules de uma variedade de taxa de samambaias 58 , 65 , 66 , 67 , 68 , 69 , 70 . Em minas de algumas espécies de Phytoliriomyza, a trilha de frass apresenta uma mudança de uma estreita linha sinuosa para uma faixa mais ampla de pellets fecais, que, em certa medida, se assemelham às das minas da Formação Momonoki.
Leaf-mining taxa of Hymenoptera are mainly composed of sawflies (Tenthredinidae). This order was already diverse by the end of the Triassic71, with the oldest fossils dating back to the Middle Triassic72. Leaf-mining sawflies produce large conspicuous blotch mines53,73 and the larvae deposit cylindrical frass pellets which are scattered irregularly about the mine cavities58,74. The extant fern-feeders of Hymenoptera do not include pinnule-miners, and only Blasticotomidae and Tenthredinidae, as petiole-borers and internal fern-feeders, respectively, are known59.
Os coleópteros mineiros são conhecidos de Buprestoidea, Chysomeloidea, Curculionoidea e alguns outros clados 60 ; taxa de mineração samambaia-pinnule são relatados a partir dessas superfamílias 59 . Os besouros tornaram-se difundidos em todo o mundo no Triássico Médio e Superior 75 , 76 , 77 . O grupo mais antigo de besouros, Protocoleoptera, é encontrado na Formação Momonoki 34 , 45 , embora se acredite que sejam saproxilicos (isto é, brocas de madeira em decomposição), com base em algumas evidências circunstanciais 78 . Minas lineares contendo pellets fecais granulares do Triássico são frequentemente atribuídas a besouros, por exemplo, Polyphaga 79 . Notavelmente, as minas de buprestida têm algumas características distintivas que são comparáveis às das minas descritas. As minas de buprestida são tipicamente manchadas; muitas vezes são minas de profundidade total, evitando tecidos epidérmicos e vasculares; os rastros de excremento são geralmente longos e fibrosos quando depositados de fresco, podendo posteriormente fragmentar-se e tornar-se granular 58 , 80 , 81 , 82 , 83 .
Os minadores de folhas de lepidópteros consistem principalmente em grupos de microlepidópteros, incluindo Nepticuloidea, Gracillaroidea e parte de Yponomeutoidea. Com base em uma filogenia molecular recente calibrada por fósseis 84 , o aparecimento de clados de mariposas-mineiras, representados pela divisão entre Nepticulidae e Opostegidae, remonta ao Jurássico Superior; para calibrar esta filogenia, foram usados fósseis em escala de asa de Coelolepida da fronteira Triássico-Jurássica da Alemanha 85 . Os fósseis de minas de folhas de nepticulídeos mais antigos são conhecidos da Formação Dakota do Cretáceo Inferior (102 Ma) 18 , 86 , 87 . As minas de lepidópteros apresentam variação considerável na forma da mina, consumo de tecido e conteúdo (por exemplo, pellets fecais). Notavelmente, as minas foliares de Ectoedemia (Nepticulidae) normalmente começam como galerias finas e fortemente sinuosas que posteriormente se tornam manchas largas 87 . Além disso, nepticulídeos típicos geralmente deixam pelotas granulares, com mudanças abruptas no padrão de acumulação em algumas espécies. É importante ressaltar que essas características da mina são vistas na Cladophlebis da Formação Momonoki. As mariposas Gracillariidae produzem minas serpentinas ou blotch e o modo de mineração de folhas é conservado no nível da subfamília 88 . Muitos táxons de gracillarídeos (Acrocercopinae, alguns Gracillariinae e Ornixolinae e Lithocollectinae) formam uma mina linear estreita durante a fase de alimentação de seiva dos ínstares iniciais, que mais tarde se torna uma mina de manchas simples. No que diz respeito à evidência fóssil e molecular, é, portanto, improvável que um membro Nepticulidae tenha causado a mina do Triássico Superior, embora o Nepticuloidea ou outro grupo de mariposas mineiras não possa ser descartado.
Ao todo, a forma das Cladophlebis não se conforma às minas típicas de Diptera (Agromyzidae) e Hymenoptera; em vez disso, ele compartilha mais recursos com os de Coleoptera ou Lepidoptera em termos de formas de mina. A atribuição de uma mina fóssil a um grupo taxonômico particular de insetos está sujeita a incertezas e limitações 19 . Os relatos biológicos disponíveis de insetos mineiros são limitados e as minas frequentemente não são fotografadas ou ilustradas. Particularmente, neste caso, uma série de características diagnósticas – hábito de oviposição (ex. não facilmente reconhecível. Além disso, a mina aqui registrada é anterior ao tempo estimado de divergência dos principais clados de insetos mineiros da era moderna, conforme examinado acima. No geral, o potencial minador pode ser Coleoptera ou Lepidoptera, em termos de semelhança geral da forma da mina, tempo de aparecimento e presença de registros em nível de ordem, embora a possibilidade de Hymenoptera e Diptera não esteja completamente excluída , devido à falta de recursos diagnósticos conclusivos. Futuros estudos taxonômicos sobre os fósseis de insetos do mesmo depósito forneceriam suporte para determinar o suspeito minador.
The stoichiometric footprint of the studied plant–insect interaction
As análises elementares indicam variabilidade quantitativa em alguns elementos (Si, P, S) que podem ser parcialmente responsáveis por processos fisiológicos nos ciclos nutricionais (Fig. 3 ); os tecidos vegetais são depositados e parcialmente removidos por um inseto e então o inseto metaboliza e excreta as substâncias não digeridas. O conteúdo do excremento é, portanto, o produto da absorção, metabolismo e excreção. Em comparação com a nervura foliar, os excrementos fossilizados (coprólitos) mostram-se altamente fosfáticos, o que é consistente com o fato de os coprólitos serem em grande parte compostos de fosfato de cálcio.
Outro ponto notável é a variação da intensidade de Si entre os pontos amostrais. Para a lâmina foliar, Si biogênico e litológico podem ser conjugados e, portanto, é necessário cautela na interpretação de nosso resultado de que a maior intensidade de Si foi encontrada na lâmina foliar. No entanto, a diferença incremental de Si entre o excremento e a nervura foliar pode ser resultado da sílica biogênica contida no excremento. A biomineralização da sílica, especialmente na forma de fitólitos (SiO 2 , nH 2 O), é encontrada em muitos clados de pteridófitas 89 , 90 , e esses fitólitos podem aumentar a resistência das plantas à alimentação de herbívoros 91 , 92 . Este teor relativamente mais alto de Si nos coprólitos de frass pode, portanto, refletir compostos defensivos não digeridos.
Este estudo ilustra que a estequiometria ecológica, um método que traça o fluxo de energia e elementos nos ecossistemas, pode ser aplicada a interações entre plantas e herbívoros endofíticos que ocorreram há 220 milhões de anos. Da mesma forma, as análises de fluorescência de raios X síncrotron (SRS-XRF) para fósseis de folhas excepcionalmente preservadas de Acer pseudoplatanus L. da Formação Green River (Eoceno, ca. 50 Ma) 93 revelaram uma alta concentração de alguns elementos metálicos (por exemplo, Cu, S, Zn) nas plantas e nos materiais fecais; no entanto, o fósforo nas folhas estava abaixo dos limites de detecção. Investigações futuras de paisagens químicas de plantas (nutrição e defesa) em várias configurações temporais e espaciais forneceriam novos insights sobre os padrões macroevolutivos de combate entre plantas e herbívoros.
Origin and early history of leaf-mining
As minas de folhas credíveis estão ausentes ou muito raras antes do Triássico Superior 27 . A partir do Paleozóico, pelo menos dois tipos de vestígios fósseis foram anteriormente atribuídos como possíveis minas de folhas, embora isso não seja suportado atualmente. Um tipo diz respeito a estruturas em forma de U ou V em pínulas de medulares do Carbonífero Superior ou Permiano Inferior 94 , 95 , que foram posteriormente atribuídas a infecção fúngica ou bacteriana 27 , 96 , 97 . Os outros tipos representam uma série de pequenas e extensas estruturas serpentinas 94 , 98 , 99 , cujos nomes de icnotaxon são Asteronomus mendriformis Müller e A. divergens Müller; atualmente, são reconhecidas como estruturas de origem tafonômica 27 . Um exemplo notável do início do Permiano é uma possível mina em um megafilo de Glossopteris cf. indica da Formação Rio Bonito, Mina Morro do Papaléo, Brasil; essa estrutura continha o possível excremento do mineiro e terminava com uma expansão terminal, que possivelmente era uma câmara larval/pupal 100 . Outro exemplo é um contorno em forma de U na folhagem de Vjaznikopteris rigida Naugolnykh da Bacia do Rio Volga (o limite P–Tr), Rússia Europeia 101 . No entanto, a identidade desses vestígios como minas de folhas é discutível porque os critérios acima mencionados para minas de insetos não são atendidos.
A partir do Triássico Médio-Final, várias formas distintas de estruturas de mineração são conhecidas (Fig. 4 ), e algumas delas são descritas como tipos de danos distintos (DT) 102 . Vários sítios da Formação Molteno (Carnian) são uma rica fonte de fósseis de herbivoria e oviposição 79 , 103 . Dois tipos de minas foliares, um de cada folhagem de Heidiphyllum elongatum (Morris) (DT41, DT71) e um de Sphenobaiera schenckii (Feistmantel) Florin (DT139), foram registrados nesta localidade 79 (Fig. 4 A); além disso, esta localidade possui uma mina não descrita e bem preservada sobre Cladophlebis (Labandeira, CC, com . pess. ), embora sua forma difere consideravelmente da da Formação Momonoki. Outro registro da flora Gondwanan é uma mina serpentina na Heidiphyllum Retallack, Triassohyponomus dinmorensis Rozefelds et Sobbe , da Formação Blackstone (Carnian) do Ipswich Coal Measures Group, Queensland, Austrália 104 , 105 (Fig. 4 B). Alguns fósseis de traços de várias outras localidades do Triássico Médio-Final também foram atribuídos a minas de folhas; por exemplo, uma pequena estrutura de mineração semilinear, carregada de frass (DT40) na folhagem da pteridosperma Scytophyllum bergeri Bornemann, de Monte Agnello, N. Itália (ladiniano tardio) 106 (Fig. 4 C). Algumas estruturas de mineração também são conhecidas da folhagem de Nilssonia sturii Krasser da Formação Lunz (Carnian), na parte oriental dos Alpes Calcários do Norte, Áustria 107 (Fig. 4 D); também, dois tipos de minas em algumas folhagens de gimnospermas (? Glossophyllum ) são relatados em Dzhayloucho (Ladinian-Carnian), perto de Madygen, Quirguistão 108 , 109 (Fig. 4 E).
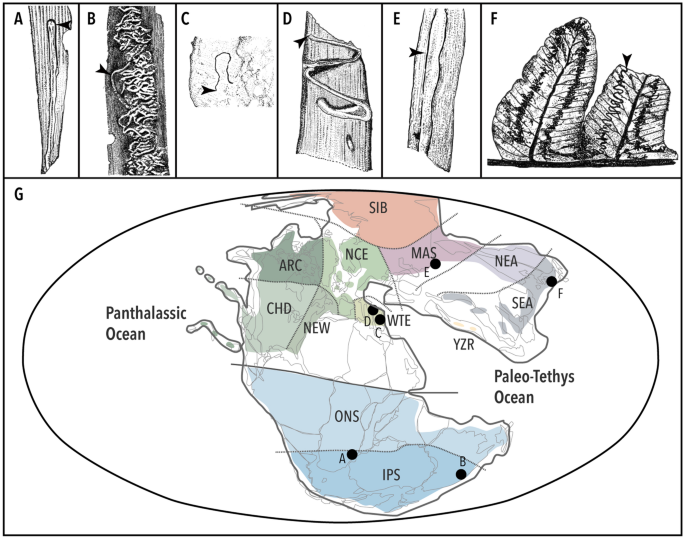
Estruturas de mineração conhecidas até agora desde o Triássico Médio-Final. ( A ) Heidiphyllum (DT71) da Formação Molteno (Carnian) 79 ; duas outras minas registradas (DT41, DT139) desta formação não estão ilustradas. ( B ) Heidiphyllum foliage mine, Triassohyponomus dinmorensis , da Blackstone Formation (Carnian) do Ipswich Coal Measures Group, Queensland, Austrália 105 . ( C ) Mina de folhagem de pteridospermas de Monte Agnello, N. Itália (Ladino tardio) 106 . ( D ) Nilssonia da Formação Lunz (Carnian), parte oriental dos Alpes Calcários do Norte, Áustria 107 . ( E ) Mina de folhagem de gimnospermas de Dzhayloucho (Ladinian-Carnian), perto de Madygen, Quirguistão 108 . ( F ) pinnule Cladophlebis da Formação Momonoki (nosso estudo). ( G ) Mapa paleogeográfico do Triássico Superior (Carniano) e as localizações aproximadas das localidades das minas fósseis são mostradas; a zonação floral é baseada em estudo anterior 44 . Cores e siglas de três letras representam zonas florais como segue: SIB Siberian Subprovince, MAS Subprovíncia da Ásia Central, NEA Subprovíncia do Norte da Ásia, Subprovíncia do Sudeste Asiático SEA Subprovíncia ARC Arctic Canada, Subprovíncia NCE Atlântico Norte/Europa Central, NOVA Newark, Subprovíncia CHD Chinle/Dockum, WTE Western Tethydean, YZR Yarlung-Zangbo- Subprovíncia River, Subprovíncia ONS Onslow, Subprovíncia IPS Ipswich. Os desenhos de linha foram feitos pela YI com o Adobe Illustrator® 2021.
As de Cladophlebis (Fig. 4 F) descritas aqui representam um novo tipo de dano que serve como a mais antiga mina fóssil credível da Região Florística do Sul da Ásia Oriental, cuja assembléia paleobotânica está geograficamente e taxonomicamente distante de qualquer um dos mencionados acima floras (Fig. 4 G). Nossa descoberta, portanto, reforça a visão de que a mineração de folhas se tornou um método de alimentação difundido para insetos que se alimentam de plantas no final do Triássico. A essa altura, eles já haviam colonizado uma ampla gama de grupos de plantas: coníferas, pteridospermas, cicadófitas, ginkgophytes e samambaias.
Methods
As interações planta-artrópodes foram recenseadas para ca. 200 caixas de armazenamento cheias (ca. 536 × 336 cm) de espécimes fósseis. Assim, a área de superfície examinada de fósseis chegou a nada menos que 3600 m 2 ; os espécimes foram coletados em várias localidades da Formação Momonoki. Embora muitas interações planta-inseto tenham sido encontradas e Cladophlebis fossem um componente dominante da assembléia floral fóssil, as minas de folhas descritas neste artigo foram encontradas em apenas um espécime. Três minas de folhas foram encontradas em um xisto, como uma parte e um espécime de contrapartida. O espécime tipo fóssil foi identificado por HY O xisto foi coletado na Rota Nacional 435 por HT com a obtenção de permissão (Fig. 1 ). O espécime do tipo fóssil está depositado e disponível ao público no Museu de História e Folclore de Mine City (MMHF), prefeitura de Yamaguchi, Japão; os códigos de catálogo são MMHF11-00001a e MMHF11-00001b. O mapa geológico (Fig. 1 ) foi feito por HY com base em estudos relevantes 38 , 39 , e posteriormente levemente modificado por YI, usando o Adobe Illustrator® 2021. O estudo de campo em plantas fósseis atende às diretrizes institucionais, nacionais e internacionais relevantes e legislação.
Fotografias de materiais foram tiradas por NO com uma Nikon D850 usando três tipos de lentes Macro-NIKKOR (120 mm F 46,3, 55 mm f 2,8 e 65 mm f 4,5). As fotos foram posteriormente editadas pela YI com o Adobe Photoshop® 2021 para aumentar o contraste e apagar o fundo. Áreas e comprimentos foram medidos por YI com Fiji (Fiji Is Just Image J) 110 . Para a preparação da Fig. 4 A–F, as ilustrações foram feitas por YI traçando fotos da literatura relevante com canetas de desenho à base de água, digitalizadas com um CanoScan LiDE 400 (Canon) com resolução de 300 dpi e editadas com Adobe Photoshop® 2021.
Para examinar a pegada estequiométrica dessa antiga interação planta-inseto, os elementos do traço da mina foram analisados por HY via fluorescência de raios-X de dispersão de energia (XRF) usando um analisador de XRF de alta sensibilidade EA6000VX (Hitachi High-Tech Science Corporation). Um espectrômetro XRF mede elementos entre Na e U em ordem crescente de número atômico. As composições elementares foram medidas quantitativamente a partir de três pontos do espécime onde permaneceram compostos orgânicos derivados de folhas fósseis (um ponto foi colocado em uma trilha de excremento de uma mina de folhas onde os coprólitos do inseto eram visíveis; cada ponto de amostra foi fixado na nervura da folha e lâmina de mesma pínula) e, como ponto de controle, da rocha sedimentar hospedeira. A duração da medição foi definida em 120 s usando um colimador de 0,2 × 0,2 mm sob uma tensão de excitação de 50 kV e uma corrente de tubo de 1000 μA. Os dados de saída foram visualizados pelo YI com ggplot2 111 e formatados com Adobe Illustrator® 2021.
Referências
Connor, EF & Taverner, MP A evolução e significado adaptativo do hábito de mineração de folhas. Oikos 79 , 6-25. https://doi.org/10.2307/3546085 (1997).
Hespenheide, HA Bionomia de insetos mineiros. Anu. Rev. Entomol. 36 , 535-560. https://doi.org/10.1146/annurev.en.36.010191.002535 (1991).
Kato, M. Estrutura, organização e resposta de uma comunidade parasitóide rica em espécies para hospedar a dinâmica populacional de bicho-mineiro. Oecologia 97 , 17-25 (1994).
López, R., Carmona, D., Vincini, A. M., Monterubbianesi, G. & Caldiz, D. Population dynamics and damage caused by the leafminer Liriomyza huidobrensis Blanchard (Diptera: Agromyzidae), on seven potato processing varieties grown in temperate environment. Neotrop. Entomol. 39, 108–114. https://doi.org/10.1590/S1519-566X2010000100015 (2010).
Lopez-Vaamonde, C., Godfray, HCJ & Cook, JM Dinâmica evolutiva do uso de plantas hospedeiras em um gênero de mariposas mineiras. Evolução 57 , 1804-1821. https://doi.org/10.1111/j.0014-3820.2003.tb00588.x (2003).
Lopez-Vaamonde, C. et ai. As filogenias moleculares calibradas por fósseis revelam que as mariposas mineradoras de folhas irradiaram milhões de anos depois de suas plantas hospedeiras. J. Evol. Biol. 19 , 1314-1326. https://doi.org/10.1111/j.1420-9101.2005.01070.x (2006).
Scheffer, SJ, Lewis, ML, Hébert, JB & Jacobsen, F. Diversidade e uso de plantas hospedeiras na América do Norte Phytomyza Holly Leafminers (Diptera: Agromyzidae): Colonização, divergência e especificidade em uma radiação associada ao hospedeiro. Ana Entomol. Soc. Sou. 114 , 59-69. https://doi.org/10.1093/aesa/saaa034 (2021).
Tooker, JF & Giron, D. A evolução da endofagia em insetos herbívoros. Frente. Planta Sci. 11 , 581816. https://doi.org/10.3389/fpls.2020.581816 (2020).
Hawkins, BA Pattern and Process in Host-Parasitoid Interactions (Cambridge University Press, 1994).
Novotny, V. & Basset, Y. Especificidade do hospedeiro de insetos herbívoros em florestas tropicais. Proc. R. Soc. B Biol. Sci. 272 , 1083-1090. https://doi.org/10.1098/rspb.2004.3023 (2005).
Lewis, OT et ai. Estrutura de uma comunidade diversificada de insetos-parasitóides de floresta tropical. J. Anim. Eco 71 , 855-873. https://doi.org/10.1046/j.1365-2656.2002.00651.x (2002).
Hirao, T. & Murakami, M. Quantitative food webs of lepidopteran leafminers and their parasitoids in a Japanese deciduous forest. Ecol. Res. 23, 159–168. https://doi.org/10.1007/s11284-007-0351-6 (2008).
Pocock, MJO, Evans, DM & Memmott, J. A robustez e restauração de uma rede de redes ecológicas. Ciência 335 , 973-977. https://doi.org/10.1126/science.1214915 (2012).
Leppänen, SA, Altenhofer, E., Liston, AD & Nyman, T. Filogenética e evolução do uso de plantas hospedeiras em flebotomíneos (Hymenoptera: Tenthredinidae: Heterarthrinae). Mol. Filogeneto. Evoluir 64 , 331-341. https://doi.org/10.1016/j.ympev.2012.04.005 (2012).
Doorenweerd, C., Van Nieukerken, EJ & Menken, SBJ Uma filogenia global de mariposas Ectoedemia (Lepidoptera: Nepticulidae): Explorando mudanças na família de plantas hospedeiras e alopatria como condutores de especiação. PLoS ONE 10 , 1–20. https://doi.org/10.1371/journal.pone.0119586 (2015).
Nakadai, R. & Kawakita, A. Phylogenetic test of speciation by host shift in leaf cone moths (Caloptilia) feeding on maples (Acer). Ecol. Evol. 6, 4958–4970. https://doi.org/10.1002/ece3.2266 (2016).
Opler, P. A. Fossil lepidopterous leaf mines demonstrate the age of some insect-plant relationships. Science 179, 1321–1323. https://doi.org/10.1126/science.179.4080.1321 (1973).
Labandeira, C. C., Dilcher, D. L., Davis, D. R. & Wagner, D. L. Ninety-seven million years of angiosperm-insect association: Paleobiological insights into the meaning of coevolution. Proc. Natl. Acad. Sci. U. S. A. 91, 12278–12282. https://doi.org/10.1073/pnas.91.25.12278 (1994).
Winkler, IS, Labandeira, CC, Wappler, T. & Wilf, P. Distinguindo as minas foliares de Agromyzidae (Diptera) no registro fóssil: Novos táxons do Paleógeno da América do Norte e Alemanha e suas implicações evolutivas. J. Paleontol. 84 , 935-954. https://doi.org/10.1666/09-163.1 (2010).
van Nieukerken, EJ, Doorenweerd, C., Hoare, RJB & Davis, DR Classificação e catálogo revisados de Nepticulidae e Opostegidae globais (Lepidoptera, Nepticuloidea). Zookeys 2016 , 65–246. https://doi.org/10.3897/zookeys.628.9799 (2016).
Maccracken, SA, Sohn, J.-C., Miller, IM & Labandeira, CC Uma nova mina de folhas do Cretáceo Superior Leucopteropsa spiralae gen. et sp. novembro (Lepidoptera: Lyonetiidae) representa a primeira evidência fóssil confirmada de Cemiostominae. J. Syst. Paleontol. 19 , 131-144. https://doi.org/10.1080/14772019.2021.1881177 (2021).
Wilf, P., Labandeira, C. C., Johnson, K. R. & Ellis, B. Decoupled plant and insect diversity after the end-Cretaceous extinction. Science 313, 1112–1115. https://doi.org/10.1126/science.1129569 (2006)
Donovan, M. P., Wilf, P., Labandeira, C. C., Johnson, K. R. & Peppe, D. J. Novel insect leaf-mining after the end-Cretaceous extinction and the demise of Cretaceous leaf miners, Great Plains, USA. PLoS ONE 9, e103542. https://doi.org/10.1371/journal.pone.0103542 (2014).
Donovan, MP, Iglesias, A., Wilf, P., Labandeira, CC & Cúneo, NR Recuperação rápida de associações planta-inseto da Patagônia após a extinção do final do Cretáceo. Nat. Eco Evoluir 1 , 0012. https://doi.org/10.1038/s41559-016-0012 (2017).
Donovan, MP, Wilf, P., Iglesias, A., Cúneo, NR & Labandeira, CC Interações bióticas persistentes de uma conífera Gondwanan da Patagônia Cretácea à Malesia moderna. Comum. Biol. 3 , 708. https://doi.org/10.1038/s42003-020-01428-9 (2020).
Labandeira, CC As quatro fases das associações planta-artrópodes no tempo profundo. Geol. 409-438 4 . https://doi.org/10.1344/105.000000344 (2006).
Labandeira, C. C. Silurian to Triassic plant and hexapod clades and their associations: new data, a review, and interpretations. Arthropod Syst. Phylogen. 64, 53–94 (2006).
Wakita, K., Nakagawa, T., Sakata, M., Tanaka, N. & Oyama, N. Phanerozoic accretionary history of Japan and the western Pacific margin. Geol. Mag. https://doi.org/10.1017/s0016756818000742 (2018).
Katayama, M. Stratigraphical study on the Mine Series. J. Geol. Soc. Jpn. 46, 127–141. https://doi.org/10.5575/geosoc.46.127 (1939).
Maeda, H. & Oyama, N. Stratigraphy and fossil assemblages of the Triassic Mine Group and Jurassic Toyora Group in western Yamaguchi Prefecture. J. Geol. Soc. Japan 125, 585–594. https://doi.org/10.5575/geosoc.2019.0020 (2019).
Aizawa, J. Fossil insect-bearing strata of the Triassic Mine Group, Yamaguchi Prefecture. Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist. Ser. A 10, 91–98 (1991).
Oyama, N. & Maeda, H. Madygella humioi sp. nov. from the Upper Triassic Mine Group, Southwest Japan: The oldest record of a sawfly (Hymenoptera: Symphyta) in East Asia. Paleontol. Res. 24, 64–71 (2020).
Fujiyama, I. Mesozoic insect fauna of East Asia part 1. Introduction and upper Triassic faunas. Bull. Natl. Sci. Mus. 16, 331–386 (1973).
Fujiyama, I. Late Triassic insects from Mine, Yamaguchi, Japan, Part 1. Odonata. Bull. Natl. Sci. Mus. Tokyo Ser. C 17, 49–56 (1991).
Ueda, K. A Triassic fossil of scorpion fly from Mine, Japan. Bull. Kitakyushu Mus. Nat. Hist. Hum. Hist. Ser. Ser. A 10, 99–103 (1991).
Takahashi, F., Ishida, H., Nohara, M., Doi, E. & Taniguchi, S. Ocorrência de fósseis de insetos do Late Triassic Mine Group. Touro. Cidade Mina Mus. prefeito de Yamaguchi. Jpn. 13 , 1-27 (1997).
Kametaka, M. Proveniência do grupo de minas do Triássico Superior Sudoeste do Japão. J. Geol. Soc. Jpn. 105 , 651-667 (1999).
Takahashi, E. & Mikami, T. Triassic. Em Geologia da Prefeitura de Yamaguchi (ed. Museu Yamaguchi) 93–108 (Museu Yamaguchi, 1975).
Kiminami, Grupo K. Atsu e Grupo Mina. Em Monografia sobre Geologia do Japão 6, Região de Chugoku (ed. Sociedade Geológica do Japão) 85–88 (Asakura Publishing Co., Ltd., 2009).
Naito, G. Fósseis de Plantas do Grupo Mina (Comitê de Educação da Cidade de Minas, 2000).
Kimura, T. Distribuição geográfica de plantas paleozóicas e mesozóicas no leste e sudeste da Ásia. Hist. Biogeografia Placa Tecton. Evoluir Jpn. Ásia Oriental 1982 , 135-200 (1987).
Kimura, T., Naito, G. & Ohana, T. Baiera cf. furcata (Lindley and Hutton) Braun from the Carnic Momonoki Formation, Japan. Bull. Natl. Sci. Mus. 9, 91–114 (1983).
Katagiri, T. Pallaviciniites oishii (comb. Nov.), a thalloid liverwort from the Late Triassic of Japan. Bryologist 118, 245–251. https://doi.org/10.1639/0007-2745-118.3.245 (2015).
Kustatscher, E. et al. Flora of the Late Triassic. In The Late Triassic World, Topics in Geobiology, Vol. 46 (ed. Tanner, L. H.) 545–622 (Springer, 2018). https://doi.org/10.1007/978-3-319-68009-5_13.
Oyama, N., Yukawa, H. & Maeda, H. Mesozoic insect fossils of Japan: Significance of the Upper Triassic insect fauna of the Mine Group, Yamaguchi Pref. Bull. Mine City Mus. Yamaguchi Prefect. Jpn. 33, 1–13 (2020).
Shcherbakov, DE, Lukashevich, ED & Blagoderov, V. Triássico Diptera e radiação inicial da ordem. Int. J. Dipterol. Res. 6 , 75-115 (1995).
Krzemiński, W. & Krzemińska, E. Triássico Diptera: Descrições, revisões e relações filogenéticas. Acta Zool. Cracóv. 46 , 153-184 (2003).
Blagoderov, V., Grimaldi, DA & Fraser, NC Como o tempo voa para as moscas: Diptera diversos do Triássico da Virgínia e radiação precoce da ordem. Sou. Mús. Nov. 3572 , 1-39. https://doi.org/10.1206/0003-0082(2007)509[1:HTFFFD]2.0.CO;2 (2007).
Lukashevich, E. D., Przhiboro, A. A., Marchal-Papier, F. & Grauvogel-Stamm, L. The oldest occurrence of immature Diptera (Insecta), Middle Triassic France. Ann. la Société Entomol. Fr. 46, 4–22. https://doi.org/10.1080/00379271.2010.10697636 (2010).
Schmidt, A. R. et al. Arthropods in amber from the Triassic Period. Proc. Natl. Acad. Sci. 109, 14796–14801. https://doi.org/10.1073/pnas.1208464109 (2012).
Lara, MB & Lukashevich, ED O primeiro díptero do Triássico (Insecta) da América do Sul, com revisão de Hennigmatidae. Zootaxa 3710 , 81-92. https://doi.org/10.11646/zootaxa.3710.1.6 (2013).
Kimura, T. & Ohana, T. Algumas samambaias fósseis da Formação Middle Carnic Momonoki, prefeitura de Yamaguchi, Japão. Touro. Nacional Sci. Mús. Ser. C Geol. Paleontol. 6 , 73-92 (1980).
Hering, EM Biology of the Leaf Miners https://doi.org/10.1007/978-94-015-7196-8 . (Primeiro, 1951).
Kirichenko, N. et al. Systematics of Phyllocnistis leaf-mining moths (Lepidoptera, Gracillariidae) feeding on dogwood (Cornus spp.) in Northeast Asia, with the description of three new species. Zookeys 2018, 79–118. https://doi.org/10.3897/zookeys.736.20739 (2018).
Cerdeña, J. et al. Phyllocnistis furcata sp. nov.: A new species of leaf-miner associated with Baccharis (Asteraceae) from Southern Peru (Lepidoptera, Gracillariidae). Zookeys 2020, 121–145. https://doi.org/10.3897/zookeys.996.53958 (2020).
Elb, P. M., Melo-de-Pinna, G. F. & de Menezes, N. L. Morphology and anatomy of leaf miners in two species of Commelinaceae (Commelina diffusa Burm. F. and Floscopa glabrata (Kunth) Hassk). Acta Bot. Brasilica 24, 283–287. https://doi.org/10.1590/S0102-33062010000100030 (2010).
Vasco, A., Moran, RC & Ambrose, BA A evolução, morfologia e desenvolvimento de folhas de samambaia. Frente. Planta Sci. 4 , 1-16. https://doi.org/10.3389/fpls.2013.00345 (2013).
Eiseman, C. Leafminers da América do Norte. (Charley Eiseman, 2019).
Yang, J., Wang, X., Duffy, K. & Dai, X. Uma lista preliminar mundial de insetos mineradores de samambaias. Biodivers. Dados J. 9 , e62839. https://doi.org/10.3897/BDJ.9.e62839 (2021).
Ding, Q., Labandeira, CC & Ren, D. Biologia de um minador de folhas (Coleoptera) em Liaoningocladus boii (Coniferales) do Cretáceo Inferior do nordeste da China e a biologia de mineração de folhas de possíveis clados de insetos culpados. Artrópode Syst. Filogenia. 72 , 281-308 (2014).
Boucher, S. Revision of the Canadian species of Amauromyza Hendel (Diptera: Agromyzidae). Can. Entomol. 144, 733–757. https://doi.org/10.4039/tce.2012.80 (2012).
Scheirs, J., Vandevyvere, I. & De Bruyn, L. Influence of monocotyl leaf anatomy on the feeding pattern of a grass-mining agromyzid (Diptera). Ann. Entomol. Soc. Am. 90, 646–654 (1997).
Boucher, S. Moscas mineiras (Diptera: Agromyzidae). Na Enciclopédia de Entomologia (ed. Capinera JL) 2163–2169 (Springer, 2008). https://doi.org/10.1007/978-1-4020-6359-6 .
Eiseman, CS Novos registros de criação para minadora muscoide (Diptera: Anthomyiidae, Scathophagidae) nos Estados Unidos. Proc. Entomol. Soc. Lavar. 120 , 25-50. https://doi.org/10.4289/0013-8797.120.1.25 (2018).
Meikle, AA Os insetos associados à samambaia. Agrícola. Prog. 14 , 58-61 (1937).
Lawton, JH A estrutura da comunidade de artrópodes na samambaia. Robô. J. Lin. Soc. 73 , 187-216. https://doi.org/10.1111/j.1095-8339.1976.tb02022.x (1976).
Lawton, J. H., MacGarvin, M. & Heads, P. A. Effects of altitude on the abundance and species richness of insect herbivores on bracken. J. Anim. Ecol. 56, 147–160. https://doi.org/10.2307/4805 (1987).
Cooper-Driver, Gi. A. Insect-fern associations. Entomol. Exp. Appl. 24, 310–316. https://doi.org/10.1111/j.1570-7458.1978.tb02787.x (1978).
Eiseman, C. S. Further Nearctic rearing records for phytophagous muscoid flies (Diptera: Anthomyiidae, Scathophagidae). Proc. Entomol. Soc. Washingt. 122, 595–603. https://doi.org/10.4289/0013-8797.122.3.595 (2020).
Santos, MG & Maia, VC Uma sinopse de galhas de samambaia no Brasil. Biota Neotrop. 18 , e20180513. https://doi.org/10.1590/1676-0611-BN-2018-0513 (2018).
Peters, RS et ai. História evolutiva dos himenópteros. atual Biol. 27 , 1013-1018. https://doi.org/10.1016/j.cub.2017.01.027 (2017).
Ronquist, F. et ai. Uma abordagem de evidência total para datação com fósseis, aplicada à radiação inicial dos himenópteros. Sistema Biol. 61 , 973-999. https://doi.org/10.1093/sysbio/sys058 (2012).
Needham, JG, Frost, SW & Tothill, BH Leaf-Mining Insects (Waverly Press, 1928).
Smith, DR, Eiseman, CS, Charney, ND & Record, S. Um novo Neártico Scolioneura (Hymenoptera, Tenthredinidae) folhas de mineração de Vaccinium (Ericaceae). J. Hymenopt. Res. 43 , 1-8. https://doi.org/10.3897/JHR.43.4546 (2015).
Zheng, D. et ai. Radiação de insetos do Triássico Médio-Final revelada por diversos fósseis e idades isotópicas da China. Sci. Av. 4 , eat1380. https://doi.org/10.1126/sciadv.aat1380 (2018).
Zhang, SQ et ai. História evolutiva de Coleoptera revelada por extensa amostragem de genes e espécies. Nat. Comum. 9 , 1-11. https://doi.org/10.1038/s41467-017-02644-4 (2018).
McKenna, DD et ai. A evolução e a base genômica da diversidade de besouros. Proc. Nacional Acad. Sci. 116 , 24729-24737. https://doi.org/10.1073/pnas.1909655116 (2019).
Gimmel, ML & Ferro, ML Visão geral dos coleópteros saproxilicos. Em Saproxylic Insects, Zoological Monographs , Vol. 1 (ed. Ulyshen, MD) 51–128 (Springer, 2018). https://doi.org/10.1007/978-3-319-75937-1_2 .
Labandeira, C. C., Anderson, J. M. & Anderson, H. M. Expansion of arthropod herbivory in Late Triassic South Africa: The Molteno Biota, Aasvoëlberg 411 site and developmental biology of a gall. In The Late Triassic World, Topics in Geobiology Vol. 46 (ed. Tanner, L. H.) 623–719 (Springer International Publishing AG, 2018).
Fiebrig, K. Eine Schaum bildende Käferlarve Pachyschelus spec. (Bupr. Sap.) Die Ausscheidung von Kautschuk aus der Nahrung und dessen Verwertung zu Schutzzwecken (auch bei Rhynchoten). Z. f. Wiss. Insektenbiol. 4, 333–339 (1908).
Bruch, C. Metamórfosis de Pachyschelus undularius (Burm.). Physis 3, 30–36 (1917).
Herring, Minas Neotropicais de Buprestide EM. Arb. Fisiol. Angew. Entomol. 9 , 241-249 (1942).
Kogan, M. Contribuição ao conhecimento da sistemática e biologia de buprestídeos minadores do gênero Pachyschelus Solier, 1833: (Coleoptera, Buprestidae). Mem. Inst. Oswaldo Cruz 61 , 429–457 (1963).
Kawahara, AY et ai. A filogenia revela o tempo evolutivo e o padrão de borboletas e mariposas. Proc. Nacional Acad. Sci. 116 , 22657-22663. https://doi.org/10.1073/pnas.1907847116 (2019).
Van Eldijk, TJB et ai. Uma janela Triássico-Jurássica para a evolução dos lepidópteros. Sci. Av. 4 , e1701568. https://doi.org/10.1126/sciadv.1701568 (2018).
Sohn, J. C., Labandeira, C. C., Davis, D. & Mitter, C. An annotated catalog of fossil and subfossil Lepidoptera (Insecta: Holometabola) of the world. Zootaxa. https://doi.org/10.11646/zootaxa.3286.1.1 (2012).
Doorenweerd, C., Van Nieukerken, E. J., Sohn, J. C. & Labandeira, C. C. A revised checklist of Nepticulidae fossils (Lepidoptera) indicates an Early Cretaceous origin. Zootaxa 3963, 295–334. https://doi.org/10.11646/zootaxa.3963.3.2 (2015).
Kawahara, A. Y. et al. A molecular phylogeny and revised higher-level classification for the leaf-mining moth family Gracillariidae and its implications for larval host-use evolution. Syst. Entomol. 42, 60–81. https://doi.org/10.1111/syen.12210 (2017).
Mazumdar, J. Phytoliths of pteridophytes. S. Afr. J. Bot. 77, 10–19. https://doi.org/10.1016/j.sajb.2010.07.020 (2011).
Trembath-Reichert, E., Wilson, J. P., McGlynn, S. E. & Fischer, W. W. Four hundred million years of silica biomineralization in land plants. Proc. Natl. Acad. Sci. U. S. A. 112, 5449–5454 https://doi.org/10.1073/pnas.1500289112 (2015).
Hunt, JW, Dean, AP, Webster, RE, Johnson, GN & Ennos, AR Um novo mecanismo pelo qual a sílica defende as gramíneas contra a herbivoria. Ana Robô. 102 , 653-656. https://doi.org/10.1093/aob/mcn130 (2008).
Reynolds, OL, Keeping, MG & Meyer, JH Resistência aumentada de silício de plantas a insetos herbívoros: Uma revisão. Ana Aplic. Biol. 155 , 171-186. https://doi.org/10.1111/j.1744-7348.2009.00348.x (2009).
Edwards, NP et ai. Metalome da folha preservado por mais de 50 milhões de anos. Metalômica 6 , 774-782. https://doi.org/10.1039/C3MT00242J (2014).
Müller, A. H. Über Hyponome fossiler und rezenter Insekten, erster Beitrag. Freib. Forschungsh. C 366, 7–27 (1982).
Beck, A. L. & Labandeira, C. C. Early Permian insect folivory on a gigantopterid-dominated riparian flora from north-central Texas. Palaeogeogr. Palaeoclimatol. Palaeoecol. 142, 139–173. https://doi.org/10.1016/S0031-0182(98)00060-1 (1998).
Jarzembowski, E. A. The oldest plant-insect interaction in Croatia: Carboniferous evidence. Geol. Croat. 65(3), 387–392. https://doi.org/10.4154/GC.2012.28 (2002).
Donovan, M. P. & Lucas, S. G. Insect herbivory on the Late Pennsylvanian Kinney Brick Quarry Flora, New Mexico, USA. Kinney Brick Quarry Lagerstätte. N. M. Mus. Nat. Hist. Sci. Bull. 84, 193–207 (2021).
Potonié, R. Ueber das Rothliegende des Thüringer Waldes. Theil II: Die Flora des Rothliegenden von Thüringen. Abh. Preuss. Geol. Landesanst. 9, 1–298 (1893).
Potonié, R. Mitteilungen über mazerierte kohlige Pflanzenfossilien. Z. Bot. 13, 79–88 (1921).
Adami-Rodrigues, KA, Iannuzzi, R. & Pinto, ID Interações planta-inseto do Permiano de uma flora Gondwana do sul do Brasil. Foss. Strat. 51 , 106-126 (2004).
Krassilov, VA & Karasev, E. Primeira evidência de interação planta-artrópode na fronteira do Permiano-Triássico na Bacia do Volga na Rússia Europeia. Alavésia 2 , 247–252 (2008).
Labandeira, CC, Wilf, P., Johnson, K. & Marsh, F. Guia para insetos (e outros) tipos de danos em fósseis de plantas comprimidos. Versão 3.0. Smithson. Instituição, Washington, DC 25 (2007).
Scott, AC, Anderson, JM & Anderson, HM Evidência de interações planta-inseto na formação Molteno do Triássico Superior da África do Sul. J. Geol. Soc. Londres. 161 , 401-410. https://doi.org/10.1144/0016-764903-118 (2004).
Tillyard, R. J. Mesozoic Insects of Queensland No. 9. Orthoptera, and Additions to the Protorthoptera, Odonata, Hemiptera, and Planipennia. Proc. Linn. Soc. N. S. W. 47, 447–470 (1922).
Rozefelds, A. C. & Sobbe, I. Problematic insect leaf mines from the Upper Triassic Ipswich Coal Measures of Southeastern Queensland Australia. Alcheringa 11, 51–57 (1987).
Wappler, T., Kustatscher, E. & Dellantonio, E. Interações planta-inseto do Triássico Médio (final do Ladiniano) de Monte Agnello (Dolomitas, N-Itália)-padrão inicial e resposta a perturbações ambientais abióticas. PeerJ 2015 , e921. https://doi.org/10.7717/peerj.921 (2015).
Meller, B., Ponomarenko, AG, Vasilenko, DV, Fischer, TC & Aschauer, B. Primeiro élitro de besouro, abdômen (Coleoptera) e um vestígio de mina de Lunz (Carnian, Late Triassic, Lunz-am-See, Áustria) e seus aspectos tafonômicos e evolutivos. Paleontologia 54 , 97-110. https://doi.org/10.1111/j.1475-4983.2010.01009.x (2011).
Vassilenko, DV Traços de interações planta-artrópodes de Madygen (Triássico, Quirguistão): Dados preliminares. Sovremennaya paleontologia: klassicheskie i noveishie metody 9–16 (2009).
Zherikhin, V. V. Insect Trace Fossils. In History of Insects (ed. Rasnitsyn A. P., Quicke, D. L.) 303–324 (Kluwer Academic Publishers, 2010).
Schindelin, J. et al. Fiji: An open-source platform for biological-image analysis. Nat. Methods 9, 676–682. https://doi.org/10.1038/nmeth.2019 (2012).
Wickham, H. ggplot2: Gráficos elegantes para análise de dados (Springer-Verlag, 2016).
Reconhecimentos
Agradecemos aos membros do Museu de História e Folclore da Cidade de Minas e do Museu de Fósseis de Minas; Hiroyuki Makara (ITCFP) e Soichiro Kawabe (FPDM/FPU) por auxiliar nas análises de XRF; Makoto Kato por nos fornecer informações valiosas sobre insetos mineiros existentes; Conrad C. Labandeira pela discussão e compartilhamento de dados inéditos; Robert B. Davis pela revisão da versão anterior do manuscrito. Este estudo foi apoiado por uma bolsa de pesquisa para Pesquisa de Campo Ambiental pela Asahi Glass Foundation (Asahi Glass Co., Ltd.), Grant-in-Aid for Scientific Research (KAKENHI) números de concessão JP20K15852 (YI) e JP19J20625 (NO) de a Sociedade Japonesa para a Promoção da Ciência (JSPS).
Informação sobre o autor
Afiliações
Contribuições
YI concebeu o projeto. YI, NO e HY contribuíram para a coleta de dados de traços fósseis. YI analisou os dados e escreveu o artigo com a contribuição de todos os coautores. YI fez as ilustrações, visualizações e desenhos, e interpretou os dados com a entrada de HY, NO; HT e HY identificaram a planta hospedeira, realizaram análises de XRF e forneceram informações paleobotânicas e geológicas. NO tirou fotos dos espécimes na Fig. 2 e forneceu informações paleoentomológicas. HT recolheu o espécime. KS contribuiu para a curadoria do acervo do museu. Todos os coautores comentaram os rascunhos do manuscrito e aprovaram o manuscrito final para publicação.
autor correspondente
Declarações de ética
Interesses competitivos
Os autores declaram não haver interesses conflitantes.
Informação adicional
Nota do editor
A Springer Nature permanece neutra em relação a reivindicações jurisdicionais em mapas publicados e afiliações institucionais.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.