O registro fóssil mais antigo de Pseudopsinae da Formação Yixian do Cretáceo Inferior do nordeste da China (Coleoptera: Staphylinidae: Pseudopsinae)
Scientific Reports volume 12 , Número do artigo: 4625 ( 2022 )
Resumo
O Cretáceo testemunhou uma radiação de besouros rove (Staphylinidae), a família de besouros mais rica em espécies. Embora a maioria das subfamílias de estafilinídeos tenham sido documentadas em estratos cretáceos em todo o mundo, não havia registro fóssil da subfamília Pseudopsinae até um fóssil relatado recentemente do âmbar de Mianmar de 99 milhões de anos. Aqui descrevemos um novo fóssil de compressão da Formação Yixian do Cretáceo Inferior do nordeste da China.
É descrito como Cretaceonanobius fossilis. geração et sp. novembro e atribuído à subfamília Pseudopsinae existente, com base nas carinas bem preservadas no pronoto, uma carina nos ventritos II e III, e mesocoxas distintamente separadas. A descoberta de Cretaceonanobius fossilis gen. et sp. novembro data o registro fóssil mais antigo de Pseudopsinae para 125 Ma no Hemisfério Norte e lança novas luzes sobre a história da evolução e paleobiogeografia desta subfamília.
Os besouros rove, Staphylinidae, não são apenas uma das famílias de besouros mais abundantes, mas também a família mais especiosa do reino animal, com mais de 65.000 espécies descritas em 33 subfamílias 1 , 2 . O registro fóssil mais antigo de Staphylinidae remonta ao Jurássico Médio e é representado por fósseis excepcionais conhecidos da Formação Haifanggou no nordeste da China (~ 165 Ma) 3 , 4 , 5 . Durante o Jurássico, foram relatados táxons pertencentes a sete subfamílias, principalmente de latitudes baixas e médias do Hemisfério Norte 6 , 7 , 8 , 9 , 10 , bem como da Austrália 10 . Embora os estudos do relógio molecular apoiem uma radiação jurássica de besouros rove 12 , não é até o Cretáceo que os Staphylinidae se diversificaram no registro fóssil, especialmente graças a espécimes bem preservados de depósitos excepcionais no leste da Ásia, como o âmbar do norte de Mianmar e os xistos da Formação Yixian na China 13 , 14 , juntamente com outros afloramentos em todo o mundo que aguardam estudo mais detalhado. Atualmente, sete subfamílias de Staphylinidae traçam sua primeira aparição no registro fóssil ao âmbar de Mianmar 8 , 15 , 16 , 17 , 18 , 19 . Cinco subfamílias de besouros, a saber, Piestinae, Oxyporinae, Paederinae, Staphylininae e Tachyporinae, bem como espécies de atribuição subfamiliar incerta, foram relatadas na Formação Yixian, representadas por 34 espécies descritas no total 10 , 20 , 21 , 22 , 23 , 24 .
Pseudopsinae é uma subfamília de estafilinídeos relativamente pequena e moderadamente diversa, pertencente ao grupo de subfamílias Staphylinine 25 . Está representado na fauna Recente por quatro gêneros com 55 espécies. Pseudopsis Newman é de longe o maior gênero que compreende 51 espécies distribuídas nas partes Holártica, Neotropical e norte das regiões orientais, bem como na Nova Zelândia. Os demais gêneros, Zalobius LeConte, Asemobius Horn e Nanobius Herman estão restritos à região Neártica ocidental 26 . Até agora, o único fóssil inequívoco do escaravelho pseudopsino foi Cretopseudopsis maweii Liu, Tihelka, Tian, Huang & Cai descrito do Cretáceo ( ca. 99 Ma) âmbar do norte de Mianmar 19 . Aqui descrevemos um novo fóssil de compressão da Formação Yixian, que retrocede ainda mais o registro fóssil mais antigo de Pseudopsinae ao Cretáceo Inferior.
Resultados
Paleontologia Sistemática
Ordem Coleoptera Linnaeus, 1758
Família Staphylinidae Latreille, 1802
Subfamília Pseudopsinae Ganglbauer, 1895
Gênero Cretaceonanobius Liu, Tihelka, Cai et Tian, Gen. nov.
Espécie tipo. Cretaceonanobius fossilis sp. novembro
Diagnóstico . Corpo de tamanho médio. Cabeça pequena, olhos grandes. Antenas curtas, localizadas na frente da cabeça. Região do pescoço distintamente constrita presente. Pronoto suborbicular, com margem lisa, carenas longitudinais do pronoto, mais largas na metade anterior. Élitros curtos, juntos um pouco mais longos que largos, cobrindo apenas parte do tergito II, expondo o restante do abdome.
Etymology. The generic name is a combination of the Latin ‘Cretaceo-’, after the age of the fossil, and the genus Nanobius.
Type material. Holotype, NIGP177043a, b. Part and counterpart with dorsal and ventral structures visible.
Locality and horizon. Huangbanjigou, Beipiao City of Liaoning Province, northeast China; Lower Cretaceous Yixian Formation (Fig. 1A,B).
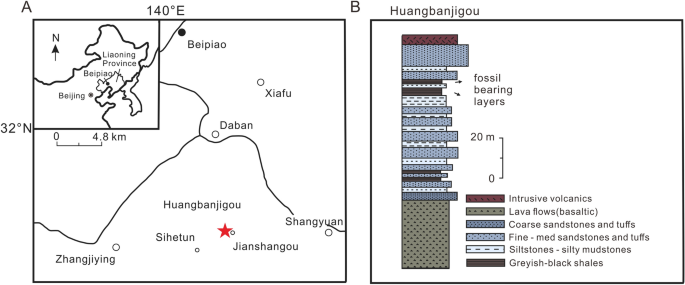
Contexto geográfico e estratigráfico da Formação Yixian, após Liu et al. 54 . ( A ) Mapa da Província de Liaoning ocidental, 56 com a estrela vermelha indicando a localidade fóssil em Huangbanjigou (modificado de Cai et al. 55 ). ( B ) Estratos fósseis expostos em Huangbanjigou (modificado de Wang et al. 56 ).
Etimologia . O epíteto específico refere-se à natureza fóssil do táxon.
Diagnóstico . Quanto ao gênero (vide supra) e uma combinação de pontuação grosseira da cabeça e pronoto, pronoto com carinas longitudinais.
Descrição . Corpo de tamanho médio e alongado, com 5,75 mm de comprimento do clípeo ao ápice abdominal (Fig. 2 ).

Habitus de Cretaceonanobius fossilis gen et sp. novembro (holótipo, NIGP 177043), parte ( a ) (dorsal) e contraparte ( b ) (ventral) umedecidas com álcool 75% sob iluminação de baixo ângulo; ( c ) corpo da linha lateral dorsal no desenho e ( d ) corpo da face ventral no desenho. Barras de escala = 2 mm em ( a ), ( b ) e 1 mm em ( c ), ( d ).
Cabeça, incluindo mandíbulas, 0,97 mm de comprimento, mais larga nos olhos (Fig. 4 a,g). Mandíbulas robustas, curvadas mesalmente (Figs. 3 b,f, 4 b,h). Palpo maxilar alongado, 4 segmentos (Fig. 2 b,d). Antenas filiformes e estreitas, 0,78 mm de comprimento, 11 segmentos, sem maça. Antenômeros 1-3 alongados, cerca de 1,2 vezes mais longos que largos (Fig. 4 c,i). Todos os antenômeros simétricos. Dorso da cabeça densamente pontuado (Fig. 2 a,c). Olhos compostos de tamanho moderado. Ocelos ausentes. Suturas gulares visíveis. Pescoço estreito, contraído (Fig. 4 e,k).

Morphological details of Cretaceonanobius fossilis gen. et sp. nov. (holotype, NIGP 177043). (a) pronotum of the part (arrows, carina on the pronotum); (b) head, ventral; (c) left elytron (arrow, hypomeron); (d) base of abdomen (arrow, intermetacoxal carina); (e) same as (a), line drawing; (f) same as (b), line drawing; (g) same as (c), line drawing; (h) same as (d), line drawing. Scale bars = 200 μm.
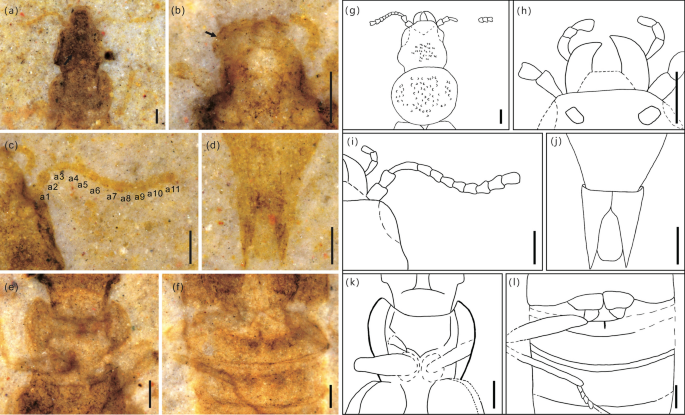
Morphological details of Cretaceonanobius fossilis gen. et sp. nov. (holotype, NIGP 177043). (a) Head, dorsal (arrow, punctation on head); (b) head, ventral (arrow, maxillary palp); (c) antenna; (d) latero-apical parts of segment X apex, ventral; (e) neck and procoxae; (f) metacoxae and metathoracic leg; (g) same as (a), line drawing; (h) same as (b), line drawing; (i) asame as (c), line drawing; (j) same as (d), line drawing; (k) same as (e), line drawing; (l) same as (f), line drawing. Abbreviations: a = antennomere; m = mandible; prc = procoxa. Scale bars = 200 μm.
Pronoto 0,73 mm de comprimento, 1,28 vezes mais comprido que largo, cerca de 0,52 vezes mais comprido que os élitros. Ângulos pronotais anteriores arredondados. Pronoto aproximadamente suborbicular, alargando-se anteriormente, com cinco carenas distribuídas sobre ele (Fig. 3 a,e). Ângulos pronotais posteriores arredondados. Prosterno curto, procoxas contíguas, suborbicular (Fig. 2 b,d). Mesocoxas ovadas, contíguas, suborbiculares. Protrochantina oculta. Processos mesoventral posterior e metaventral anterior presentes. Metacoxas estreitamente separadas.
Élitro curto, 1,38 mm de comprimento, 1,14 vezes mais largo que longo. Superfície elitral pontilhada grosseira e carenas longitudinais, hipomero visível em vista ventral (Fig. 3 c,g).
Pernas longas, delgadas (Fig. 4 f,l). Abdome com 3,65 mm de comprimento, com seis esternitos visíveis. Seis tergitos visíveis dorsalmente. Processo intercoxal do esternito II triangular (Fig. 3 d,h). Tergitos III–VII subiguais em comprimento e cada um com um par de laterosternitos, segmentos VII e VIII gradualmente estreitados, tergito VIII subtriangular, tergito X com grande extensão posterior (Fig. 4 d,j). Abdômen com fileiras de esculturas subtriangulares. Dois espinhos da cauda presentes.
Discussão
Colocação sistemática
The new fossil can be excluded from all extant staphylinid families, with the exception of Pseudopsinae. Its elongate body and antennae located on the front of the head, differentiate the new taxon, from the known members of Mesozoic Staphylininae and Aleocharinae27,28. Based on the lack of a pair of ocelli and a narrower body shape, the fossil is excluded from Omaliinae29. The lack of large eyes and distinctly clubbed antennae, distinguish Cretaceonanobius gen. nov. from Megalopsidiinae and Steninae30,31. Clubbed antennae, enlarged maxillary palpomere 3, and 3-segmented tarsi separate Protopselaphinae, to which Cretaceonanobius fossilis gen. et sp. nov. cannot be assigned27. Robust mandibles and a distinct labial palpus differentiate Oxyporinae and Euaesthetinae from Cretaceonanobius fossilis gen. et sp. nov.29,32. The pronotum of the Cretaceonanobius fossilis gen. et sp. nov. is suborbicular and widest medially, showing significant differences with the known Mesozoic members of Trigonurinae and Tachyporinae, which are characterized by the pronotum broadest at the base29 , 33 . Comparado com Proteininae, o Cretaceonanobius fossilis gen. et sp. novembro possui corpo mais estreito, com seis tergitos expostos 29 , 30 . A antena sem um clube apical distinto ou configuração densa mostra que não é membro de Trichophyinae ou Habrocerinae 34 , 35 . As antenas de Olisthaerinae estão localizadas na face lateral da testa 29 . Diferente do pronoto suborbicular de Cretaceonanobius fossilis gen. et sp. nov., o pronoto em Piestinae é trapezoidal invertida e a cabeça grande 27 . Com base na ausência de paratergitos em Osoriinae e dois pares de paratergitos em Olisthaerinae, Cretaceonanobius fossilis gen. et sp. novembro com apenas um par de paratergitos podem ser distinguidos de ambas as subfamílias 27 , 29 , 36 , 37 . Habrocerinae distingue-se do presente fóssil por mandíbulas assimétricas Enquanto isso, a subfamília existente Habrocerinae, cujas antenas 3-11 são delgadas e cobertas de cerdas, é diferente do Cretaceonanobius fossilis gen. et sp. novembro
Cretaceonanobius fossilis gen. et sp. nov. possesses derived characters such as the punctation on the temple and the spines on the pronotum which unite it with Pseudopsinae to the exclusion of the superficially similar Solieriinae. Based on this combination of characters, we assign C. fossilis gen. et sp. nov. to Pseudopsinae.
The general habitus of Cretaceonanobius gen. nov. resembles Pseudopsis and Nanobius36,38,39. Like many species of the genus Pseudopsis, the fossil has a single pair of laterosternites (although in Nanobius the second outside pair is very narrow and easily overlooked). Similar to Nanobius, it has a very distinct neck, pronotum with distinctly carinae, dense deep punctation on the head and pronotum and has the indication of a carina on sternites II-III. Unlike both genera, which have distributed carinae on the elytra, Cretaceonanobius gen. nov. lacks carinae on the elytra. Some modern Pseudopsis such as Pseudopsis obliterata LeConte, 1879 also lack carinae, although they have rounded elevations and uneven surfaces on the pronotum and elytra.
Cretaceonanobius fossilis gen. et sp. nov. is differentiated from Asemobius, which possesses a triangular head with long maxillary palps, neck concealed dorsally, and pronotum broadest in the anterior third. Furthermore, Zalobius is differentiated by its shape of the pronotum39. C. fossilis gen. et sp. nov. further differs from the Cretaceous Cretopseudopsis maweii from Myanmar amber in that the protrochantins of the latter are barely visible, elytra are carinate, and abdominal terga lack basolateral ridges19. Both genera however share a deeply punctate head surface, very distinct neck. Predating Cretopseudopsis from Myanmar amber, Cretaceonanobius fossilis gen. et sp. nov. represents the earliest fossil record of Pseudopsinae so far.
Geografia e história da evolução do habitat da Pseudopsina
Os Staphylinidae existentes estão amplamente distribuídos em todos os continentes, exceto na Antártida, e contribuem com uma grande parcela da biodiversidade animal em microhabitats, como serrapilheira e solo 40 . Embora o surgimento de angiospermas durante o Cretáceo, conhecido como Revolução Terrestre Cretácea ou Revolução Terrestre das Angiospermas, tenha sido associado à diversificação de algumas linhagens de besouros, ainda não está claro até que ponto isso impactou a dinâmica macroevolutiva dos besouros 41 , 42 . Embora uma origem Triássico-Jurássica de coroa-Staphylinidae apoiada por estudos de relógio molecular seja congruente com o registro fóssil 11 , no momento da origem de subfamílias individuais é menos certo 42 . Como tal, as descobertas de besouros rove dos depósitos do Cretáceo e Jurássico iniciais que antecedem o aparecimento generalizado de plantas com flores são importantes para calibrar a escala de tempo da evolução dos estafilinídeos e testar os impactos da Revolução Terrestre Cretácea na diversificação dos besouros. Os quatro gêneros de Pseudopsine são amplamente distribuídos. Zalobius é conhecido desde o sul da Colúmbia Britânica até o centro da Califórnia (Fig. 5 , círculo preto). Nanobius e Asemobius ocorrem desde o sul da Colúmbia Britânica até o sul da Califórnia e sudoeste da Califórnia (Fig. 5 , pontos vermelhos; pontos roxos). A Pseudopsis está distribuída nas regiões Neártica, Paleártica, Neotropical, Norte Oriental e Australásia 43 , 44 . Todos os quatro gêneros existentes vivem principalmente em latitudes médias e baixas e parcialmente em altas latitudes. Todas as Pseudopsinae mesozóicas conhecidas do âmbar de Myanmar e da Formação Yixian ocorreram nas regiões equatorial e médias de latitude, respectivamente 45 , 46 .
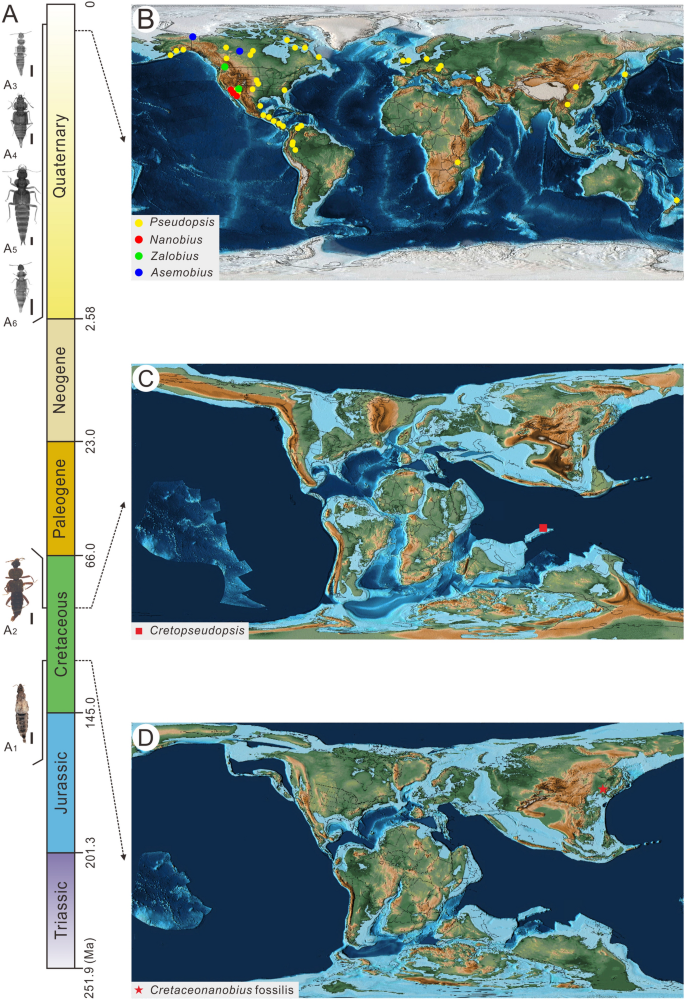
Distribution of extinct and extant Pseudopsinae taxa. (A1) Cretopseudopsis maweii, (A2) Cretaceonanobius fossilis gen. et sp. nov., (A3) Pseudopsis, (A4) Asemobius, (A5) Nanobius, (A6) Zalobius. Scale bars = 500 μm. (B) Recent geographic map after Scotese57: yellow, red, green and blue dots show the distribution of Pseudopsis, Nanobius, Zalobius and Asemobius, respectively; (C) Palaeogeographic map of the Early Cretaceous after Scotese57, with the red square for Cretopseudopsis maweii; (D) Palaeogeographic map of the Early Cretaceous after Scotese57, with the red star indicating the distribution of the Cretaceonanobius fossilis gen. et sp. nov.
Pseudopsinas existentes habitam serrapilheira, bem como detritos de enchentes e gramíneas que crescem perto de córregos, de áreas costeiras a regiões montanhosas 38 , 39 . O paleoambiente âmbar de Myanmar foi reconstruído como uma floresta tropical situada no litoral 47 sugerindo que Cretopseudopsis maweii vivia em ambientes costeiros (Fig. 5 B). Os fósseis de esporos da Formação Yixian pertencem predominantemente a gimnospermas, o tipo de vegetação dominante foi uma floresta úmida de coníferas 48 . As condições climáticas úmidas também são refletidas pelo woodstone 48 , 49 . Ao mesmo tempo, os fósseis de conchostraca foram encontrados na Formação Yixian, que vivia em águas rasas e refletia um clima semiárido a semiúmido 13 . Os insetos encontrados na Formação Yixian incluíam espécies terrestres, aquáticas e semi-aquáticas, o que indicava que havia terra, água parada e condições de vida úmida no Beipiao naquela época 50 . Os hábitos alimentares de Pseudopsinae mostram o ambiente de vida de Cretaceonanobius fossilis geração et sp. novembro pode na área pantanosa que cercada por uma antiga bacia lacustre e árvores cresciam nas altas montanhas próximas, e é consistente com o paleoambiente refletido pela assembléia fóssil da Formação Yixian 13 . Os diversos paleoambientes ocupados por pseudopsinas fósseis refletem seu amplo nicho ecológico durante o Cretáceo.
material e métodos
O espécime estudado é originário da Formação Yixian na vila de Huangbanjigou, cidade de Beipiao na província de Liaoning da China 13 . A Formação Yixian é bem conhecida pelas descobertas de numerosos dinossauros, mamíferos, pássaros, angiospermas e fósseis de insetos bem preservados 46 , 51 , 52 (Jehol Biota). Sua idade foi limitada ao Cretáceo médio, cerca de 125 Ma, por 40 Ar/ 39 Ar datando 53 .
The specimen is compressed in shales, both ventral and dorsal sides have been well preserved (Fig. 1), including morphological details such as the punctation of the head and a single pair of paratergites. Photographs were taken with a Zeiss Discovery V20 microscope equipped with a digital camera after the specimen has been moistened with 75% ethanol. The type specimen is deposited in the Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China. New nomenclatural acts established have been registered in ZooBank under the publication LSID urn:lsid: sid:zoobank.org:act:B4882FCA-1EF8-4A43-8A54-5867FB05C060.
References
Grebennikov, V. V. & Newton, A. F. Good-bye Scydmaenidae, or why the ant-like stone beetles should become megadiverse Staphylinidae sensu latissimo (Coleoptera). Eur. J. Entomol. 106, 275–301 (2009).
Yamamoto, S. & Takahashi, Y. First and oldest Leptochirini rove beetles illuminate diverse cephalic structures in the Cretaceous (Coleoptera: Staphylinidae: Osoriinae). Syst. Entomol. 44, 588–611 (2019).
Cai, C. et al. Integrated phylogenomics and fossil data illuminate the evolution of beetles. R. Soc. Open Sci. 9, 211771 (2022).
Fikáček, M. et al. Reliable placement of beetle fossils via phylogenetic analyses – Triassic Leehermania as a case study (Staphylinidae or Myxophaga?). Syst. Entomol. 45, 175–187 (2020).
Cai, C. et al. Early origin of parental care in Mesozoic carrion beetles. Proc. Natl. Acad. Sci. 111, 14170–14174 (2014).
Ryvkin, A. B. Beetles of the family Staphylinidae from the Jurassic of Transbaikalia. Trudy Paleontologicheskogo Instituta, Akademia nauk SSSR 211, 88–91 (1985).
Tikhomirova, AL Besouros estafilinídeos do Jurássico do Karatau (Coleoptera, Staphylinidae). Em Jurassic Insects of the Karatau (ed. Rohdendorf, BB) 139–154 (Akademiya Nauk SSSR, 1968) ( em russo ).
Cai, C. & Huang, D. O mais velho besouro de rove osoriine do âmbar birmanês do Cretáceo (Coleoptera: Staphylinidae). Res. Cretáceo. 52 , 495-500 (2015).
Weyenbergh, H. Sobre os insetos fósseis do calcário litográfico da Baviera, que estão no Museu Teylers. Extrato dos Arquivos do Museu Teylers 2 , 247–294 (1869).
Cai, C.-Y. & Huang, D.-Y. Diversos besouros oxiporinos do Cretáceo Inferior da China (Coleoptera: Staphylinidae). Sistema Entomol. 39 , 500-505 (2014).
Cai, C.-Y., Yan, EV, Beattie, R., Wang, B. & Huang, D.-Y. Primeiros besouros do Jurassic Talbragar Fish Bed da Austrália (Coleoptera, Staphylinidae). J. Paleontol. 87 , 650-656 (2013).
Lü, L. et al. Linking evolutionary mode to palaeoclimate change reveals rapid radiations of staphylinoid beetles in low-energy conditions. Curr. Zool. 66, 435–444 (2020).
Jiang, B. & Sha, J. Preliminary analysis of the depositional environments of the Lower Cretaceous Yixian Formation in the Sihetun area, western Liaoning, China. Cretaceous Res. 28, 183–193 (2007).
Cruickshank, R. D. & Ko, K. Geology of an amber locality in the Hukawng Valley, Northern Myanmar. J. Asian Earth Sci. 21, 441–455 (2003).
Clarke, D. J. & Chatzimanolis, S. Antiquity and long-term morphological stasis in a group of rove beetles (Coleoptera: Staphylinidae): Description of the oldest Octavius species from Cretaceous Burmese amber and a review of the “Euaesthetine subgroup” fossil record. Cretaceous Res. 30, 1426–1434 (2009).
Cai, C.-Y. & Huang, D.-Y. The oldest micropepline beetle from Cretaceous Burmese amber and its phylogenetic implications (Coleoptera: Staphylinidae). Naturwissenschaften 101, 813–817 (2014).
Chatzimanolis, S., Engel, M. S., Newton, A. F. & Grimaldi, D. A. New ant-like stone beetles in mid-Cretaceous amber from Myanmar (Coleoptera: Staphylinidae: Scydmaeninae). Cretaceous Res. 31, 77–84 (2010).
Thayer, M. K., Newton, A. F. & Chatzimanolis, S. Prosolierius, a new mid-Cretaceous genus of Solieriinae (Coleoptera: Staphylinidae) with three new species from Burmese amber. Cretaceous Res. 34, 124–134 (2012).
Liu, Y., Tihelka, E., Tian, L., Huang, D. & Cai, C. First fossil pseudopsine rove beetle from mid-Cretaceous Burmese amber (Coleoptera: Staphylinidae: Pseudopsinae). Zootaxa 4885, 76–82 (2020).
Yue, Y., Ren, D. & Solodovnikov, A. The oldest fossil species of the rove beetle subfamily Oxyporinae (Coleoptera: Staphylinidae) from the Early Cretaceous (Yixian Formation, China) and its phylogenetic significance. J. Syst. Palaeontol. 9, 467–471 (2011).
Yue, YL, Ren, D. & Solodovnikov, A. Megolisthaerus chinensis gen. et sp. n (Coleoptera: Staphylinidae incertae sedis): Uma enigmática linhagem de besouros do Cretáceo Inferior. Sistema de insetos Evoluir 41 , 317-327 (2010).
Yue, Y., Makranczy, G. & Ren, D. Uma espécie mesozóica de Anotylus (Coleoptera, Staphylinidae, Oxytelinae) de Liaoning, China, com as primeiras evidências de dimorfismo sexual em besouros rove. J. Paleontol. 86 , 508-512 (2012).
Solodovnikov, A., Yue, Y., Tarasov, S. & Ren, D. Besouros rove extintos e existentes se encontram na matriz: Fósseis do Cretáceo Inferior lançam luz sobre a evolução de uma linhagem de insetos hiperdiversa (Coleoptera: Staphylinidae: Staphylininae). Cladística 29 , 360-403 (2013).
Yue, Y., Gu, J.-J., Yang, Q., Wang, J. & Ren, D. A primeira espécie fóssil da subfamília Piestinae (Coleoptera: Staphylinidae) do Cretáceo Inferior da China. Res. Cretáceo. 63 , 63-67 (2016).
Thayer, MK Staphylinidae. No Manual de Zoologia. Vol. 4. Parte 38. 2ª Edição. Coleópteros. Vol. 1. Morfologia e Sistemática (Archostemata, Adephaga, Myxophaga, Staphyliniformia, Scarabaeiformia, Elateriformia) (eds Beutel, RG & Leschen, RAB) 394–442 (De Gruyter, Berlim, 2016).
Newton, AF & Thayer, MK Catálogo de Taxa Superior de Staphyliniformia e Gêneros e Subgêneros de Staphylinoidea (Museu de Campo de História Natural, 2003).
Thayer, M. Staphylinoidea. [chapter] 11.7. Staphylinidae Latreille, 1802. Morphol. Syst. Archostemata Adephaga Myxophaga Polyphaga Partim 296–344 (2005).
Ashe, J. S. Phylogeny of the tachyporine group subfamilies and ‘basal’ lineages of the Aleocharinae (Coleoptera: Staphylinidae) based on larval and adult characteristics: Phylogeny of tachyporine group staphylinids. Syst. Entomol. 30, 3–37 (2005).
Newton, A. F., Thayer, M. K., Ashe, J. S. & Chandler, D. S. Staphylinidae Latreille, 1802. In American Beetles (eds Arnett, R. H. & Thomas, M. C.) 272–418 (CRC Press, 2001).
Newton, A. F. & Thayer, M. K. Protopselaphinae new subfamily for Protopselaphus new genus from Malaysia, with a phylogenetic analysis and review of the Omaliine Group of Staphylinidae including Pselaphidae. In Biology, Phylogeny, and Classification of Coleoptera: Papers Celebrating the 80th Birthday of Roy A (eds Pakaluk, J. & Slipinski, S. A.) 219–320 (Crowson; Muzeum i Instytut Zoologii PAN, Warszawa, 1995).
Eichelbaum, F. Verbesserungen und Zusatze zu meinem Katalog der Staphylinidengattungen aus dem Jahre 1909. Archiv für Natur. (A) 81(5), 98–121 (1915).
Hanley, RS & Goodrich, MA Revisão de Micofagia, Relações de Hospedeiro e Comportamento no Novo Mundo Oxyporinae (Coleoptera: Staphylinidae). Coleopt. Touro. 49 , 267-280 (1995).
Herman, LH Catálogo dos Staphylinidae (Insecta: Coleoptera). 1758 até o final do segundo milênio. VI. Grupo Staphylinine (Parte 3) Staphylininae: Staphylinini (Quediina, Staphylinina, Tanygnathinina, Xanthopygina), XantholininiStaphylinidae Incertae SedisFossils, Protactinae†. Touro. Sou. Mús. Nat. Hist. 2001 , 3021 (2001).
Ashe, JS & Newton, AF Jr. Larvas de Trichophya e filogenia do Grupo Tachyporine de subfamílias (Coleoptera: Staphylinidae) com revisão, novas espécies e caracterização de Trichophyinae. Sistema. Ent. 18 , 267-286 (1993).
Muir, F. Os segmentos abdominais masculinos e edeagos de Habrocerus capillaricornis Grav (Coleoptera, Staphylinidae). Trans. Ent. Soc. Londres. 67 , 398-403 (1919).
Newton, AF Redefinição, filogenia revisada e relações de Pseudopsinae (Coleoptera, Staphylinidae). Sou. Mús. Nov. 2743 , 1-13 (1982).
Moore, I. & Legner, E. F. The genera of the subfamilies Phloeocharinae and Olisthaerinae of America north of Mexico with description of a new genus and new species from Washington (Coleoptera: Staphylinidae). Can. Entomol. 105, 35–41 (1973).
Herman, L. H. Revision and phylogeny of the monogeneric subfamily Pseudopsinae for the world (Staphylinidae, Coleoptera). Bull. Am. Mus. Nat. Hist. 155, 241–317 (1975).
Herman, L. H. Revision and phylogeny of Zalobius, Asemobius, and Nanobius, new genus (Coleoptera, Staphylinidae, Piestinae). Bull. Am. Mus. Nat. Hist. 159, 45–86 (1977).
Parker, J. Staphylinids. Curr. Biol. 27(2), R49–R51 (2017).
Lloyd, G. T. et al. Dinosaurs and the cretaceous terrestrial revolution. Proc. R. Soc. B Biol. Sci. 275, 2483–2490 (2008).
Benton, M. J., Wilf, P. & Sauquet, H. The Angiosperm Terrestrial Revolution and the origins of modern biodiversity. New Phytol. https://doi.org/10.1111/nph.17822 (2021).
Newton, A. F. StaphBase: Staphyliniformia world catalog database (version, Nov 2018). In Species 2000 & ITIS Catalogue of Life (eds Roskov, Y. et al.) (University of Reading, 2020).
Yin, Z.-W. Two new species and additional records of Pseudopsis Newman from China (Coleoptera: Staphylinidae: Pseudopsinae). J. Nat. Hist. 55, 933–951 (2021).
Westerweel, J. et al. Burma Terrane part of the Trans-Tethyan arc during collision with India according to palaeomagnetic data. Nat. Geosci. 12, 863–868 (2019).
Ren, D. et ai. (eds) Rhythms of Insect Evolution: Evidence from the Jurassic and Cretáceo in Northern China (Wiley, 2019).
Grimaldi, D. & Ross, AJ Extraordinário Lagerstätten em âmbar, com referência particular ao Cretáceo da Birmânia. Em Conservação Terrestre Lagerstätten: Janelas para a Evolução da Vida na Terra (eds Fraser, NC & Sues, H.-D.) 287–342 (Dunedin Academic Press, Edimburgo, 2017).
Yu, JX, Pu, RG, Wu, HZ, conjuntos de pólen esporo da parte superior do grupo Rehe, província de Liaoning. Acta Geo. Pecado. 2 (1989).
Zheng, SL, Zheng, YJ & Xing, DH Características idade e clima da Flora Yixian do Jurássico Superior de Liaoning Ocidental. J. Estratigr. 27 (3), 233-241 (2003).
Zhang, H., Wang, B. & Fang, Y. Evolução da diversidade de insetos no Jehol Biota. Sci. China Earth Sci. 53 , 1908-1917 (2010).
Pan, Y., Sha, J., Zhou, Z. & Fürsich, FT A Biota Jehol: Definição e distribuição de relíquias excepcionalmente preservadas de um ecossistema continental do Cretáceo Inferior. Cretáceo. Res. 44 , 30-38 (2013).
Xu, X., Zhou, Z., Wang, Y. & Wang, M. Study on the Jehol Biota: Recent advances and future prospects. Sci. China Earth Sci. 63, 757–773 (2020).
Swisher, C. C., Wang, Y. Q., Wang, X. L., Xu, X. & Wang, Y. Cretaceous age for the feathered dinosaurs of Liaoning, China. Nature 400, 58–66 (1999).
Liu, Y. et al. A transitional fossil sheds light on the early evolution of the Staphylinine group of rove beetles (Coleoptera: Staphylinidae). J. Syst. Palaeontol. 19, 321–332 (2021).
Cai, C., Huang, D. & Solodovnikov, A. A new species of Hesterniasca (Coleoptera: Staphylinidae: Tachyporinae) from the Early Cretaceous of China with discussion of its systematic position. Insect Syst. Evol. 42, 213–220 (2011).
Wang, X. et al. Vertebrate biostratigraphy of the Lower Cretaceous Yixian formation in Lingyuan, western Liaoning and its neighboring southern Nei Mongol (inner Mongolia). China. Vert. PalAs. 38(2), 95–101 (2000).
Scotese, C.R., Dreher, C. GlobalGeology (2012). http:/www.GlobalGeology.com.
Acknowledgements
This work was supported by the Strategic Priority Research Program of the Chinese Academy of Sciences (Grant No. XDB26000000), the National Natural Science Foundation of China (Grant Nos. 41688103 and 42072022), and the Youth Innovation Promotion Association of the CAS (Grant No. 2018347).
Author information
Affiliations
Contribuições
CC desenhou o estudo; YL reuniu os dados; ET e LT contribuíram com a discussão. Todos os autores escreveram o manuscrito.
Autores correspondentes
Declarações de ética
Interesses competitivos
Os autores declaram não haver interesses conflitantes.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.