Sahelanthropus tchadensis – NOSSO ÚLTIMO ANCESTRAL COMUM CHIMPANZÉ-HUMANO, ATÉ O MOMENTO.
Embora a espécie humana tenha sua origem datada em aproximadamente 200 mil anos, o processo de hominização começou a certa de 7 milhões de anos, na África. Hoje vamos tratar de um ponto muito importante na história da humanidade, o hominínio mais antigo, Salehanthropus tchadensis e espécies relacionadas a ele em um ponto específico da árvore filogenética humana (entre 7 e 5,8 milhões de anos). Este ponto é no tempo é crucial porque nos leva ao encontrado de um conceito muito importante, o “último ancestral comum chimpanzé-humano” (CHLCA).
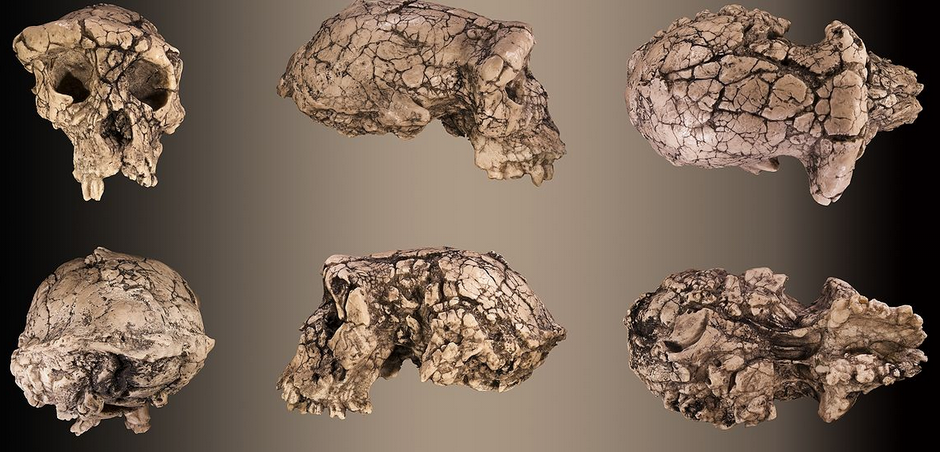
Crânio de Sahelanthropus tchadensis. Clique para ampliar
Sahelanthropus tchadensis é visto como a espécie mais antiga relacionada à evolução humana (Brunet et al, 1995). Infelizmente só há poucos fragmentos desta espécie e novas descobertas são aguardadas. Estudos anatômicos indicam que este hominínio era bípede.
Os fósseis de S. tchadensis foram descobertos no deserto Djurab do Chade por uma equipe de pesquisadores liderados por Michel Brunet. A descoberta foi feita em julho entre 2001 e março de 2002 em três locais (TM 247, TM 266 [que produziu a maioria do material] e TM 292).
Até hoje, este é o mais antigo ancestral humano conhecido depois da separação da linhagem humana e dos chimpanzés. Isto tem uma importância grande, pois destaca um conceito importante na evolução humana, o “Chimpanzee–Human Last Common Ancestor (CHLCA), ou o “último ancestral comum do homem com os chimpanzés”, que até o momento é representado pelo S. tchadensis, mas esta sujeito a mudanças mediante novas descobertas, mais antigas.
Os ossos deste hominídeo foram encontrados longe da maioria dos achados fósseis hominídeos anteriores, que são geralmente da África Oriental e Austral. Uma mandíbula de Australopithecus bahrelghazali foi encontrada em Chade por Beauvilain e Brunet e Moutaye em 1995 (Brunet et al, 1995). Como o dimorfismo sexual conhecido por ter existido nos primeiros hominídeos, alguns autores argumentam que a diferença entre o Ardipithecus e Sahelanthropus pode não ser grande o suficiente para justificar uma espécie separada para o último (Haile-Selassie et al, 2004).
Dimorfismo sexual esta relacionado a estrutura social e interfere diretamente no tamanho relativo dos dentes dos animais. Embora o dimorfismo sexual ocorra por diversos processos dependendo do grupo biológico que ele esta presente, em primatas existem algumas situações chave que representam a sua origem. Relações monogâmicas relativamente estáveis tendem a diminuir o dimorfismo sexual de tal forma que ambos os sexos tenham tamanhos próximos e a diferença no tamanho dos dentes caninos seja semelhante, como ocorre em gibões, siamangos e muitos primatas prossímios. O dimorfismo é mais expressivo quando a estrutura social é baseada em um sistema de macho alpha que tem acesso a diversas fêmeas e por isto é constantemente atacado por outros machos que buscam tomar seu posto. Por essa razão, machos maiores e com caninos mais expressivos tendem a serem selecionados e consolidar um dimorfismo mais intenso como ocorre em símios como os chimpanzés, gorilas e alguns macacos das Américas (Macacos do novo Mundo). Quando as espécies vivem de modo solitário em que o macho e a fêmea só se encontram para a reprodução, a pressão seletiva leva ao ápice do dimorfismo sexual, como ocorre em orangotangos e alguns prossímios (Neves et al, 2015). Se de fato, o S. tchadensis for uma variação de dimorfismo sexual de Ardipithecus teremos uma evidência sobre a estrutura social deste hominínio mais antigo ligado a nossa origem.

Crânio de Sahelanthropus tchadensis. Clique para ampliar e ver os detalhes.
Os achados de Sahel no Chade incluem um pequeno crânio, chamado Toumai (que na língua local significa “esperança de vida”), cinco peças de mandíbula e alguns dentes, tornando-se uma cabeça que tem uma mistura de características derivadas e primitivas. A caixa craniana compreende um volume de apenas 320 cm³ a 380 cm³, bastante semelhante ao dos chimpanzés existentes e é notavelmente menor que o volume humana aproximado, que é de 1.350 cm³ (Neves et al, 2015).
Os dentes, extremidade da testa (com traços grosseiros) e estrutura facial diferem muito daqueles encontrados no Homo sapiens. Características cranianas mostram um rosto mais liso, com uma arcada dentária em forma de “U” (contrária a do Homo sapiens que é paraboloide), caninos pequenos e um forame magno anterior. Não há restos pós-cranianos recuperados (ou seja, ossos abaixo do crânio). O crânio sofreu uma grande distorção durante o tempo de fossilização e está achatado dorso-ventralmente mostrando um lado direito deprimido (Brunet; et al, 2002).
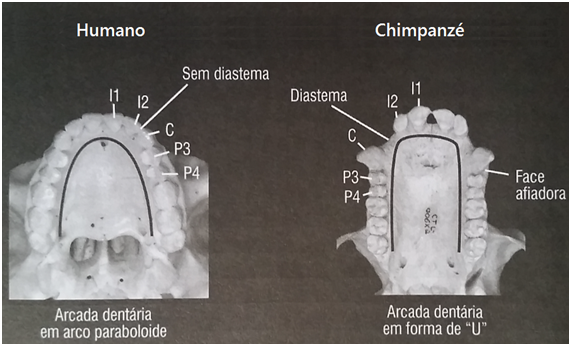
Arcada dentaria em “U” nos humanos e paraboloide em chimpanzés
Como não há restos pós-cranianos “encontrados”, não se sabe definitivamente se Sahelanthropus tchadensis era de fato bípede, apesar de apresentar um forame magno anteriormente colocado sugere que este ter sido o caso. Após uma análise do forame magno no estudo preliminar do achado autor da descoberta concluiu isto com base na morfologia base-cranial semelhante ao hominineos mais recentes (Brunet et al, 2002). Alguns paleontólogos (Wolpoff et al, 2002) contestaram esta interpretação, afirmando que o base-crânio, bem como características da dentição e faciais não representam adaptações únicas para o clado dos hominídeos ou indicativos de bipedalismo. Sendo assim, o desgaste canino seria semelhante a outros primatas do Mioceno (A new hominid from the Upper Miocene of Chad, Central Africa).
Um fêmur de um hominídeo foi descoberto ao lado (cerca de um metro de distância) do crânio de S. tchadensis em 2009, mas nada ainda foi publicado (Hawks 2009). É possivel que o fêmur esteja associada ao crânio. Este osso foi encontrado sem a extremidade distal, onde fica a da cabeça e o colo do fêmur, embora a anatomia da diáfise seja muito característica a de um hominíneo. Pode haver alguma evidência na distribuição na porção cortical no colo do fêmur, mas a ruptura distal ainda não permite fazer uma comparação válida de espessuras corticais superiores e inferiores. A não-publicação de um artigo esta relacionada a precaução em arriscar um estudo precoce a público sobre se o hominineo de fato representa um bípede.

Fêmur de Sahelanthropus tchadensis
Não significa necessariamente que o fêmur e o crânio foram depositados juntos, ao mesmo tempo ou localização exata. A dúvida é se de fato o fêmur pertence ao mesmo especime do crânio identificado. A acumulação de diferentes fósseis em uma pequena área que poderia ter acontecido como um erosão ou com o vento qu depositou o material longe do local original, alterando a topografia local (fenômeno chamado de deflação eólica). Tudo indica ser representante de uma única espécie de primata antigo, ja que o fêmur é proporcional ao tamanho esperado ao indivíduo e ao crânio. Parece improvável que duas espécies de hominíneos de porte semelhante, que viveram na mesma área durante o período de tempo limitado e específico sejam representado na mesma estratigrafia. Os paleontólogos aguardam novas descobertas embora esperava-se que algum parecer prévio e não definitivo já tivesse sido dado sobre este achado (Hawks, 2009).
A localização de S. tchadensis na árvore evolutiva humana sugere um posicionamento ancestral de outros dois hominineos; o Orrorin tugenensis e os Ardipithecus (na qual duas espécies são conhecidas), sendo a espécies A. kadabba o mais próximo de S. tchadensis depois de O. tugenensis.

Local da árvore da vida onde se encontra Sahelanthropus tchadensis, Orrorin tugenensis e Ardipithecus kadabba. Veja a árvore completa aqui.
É possível notar que O. tugenensis (6 milhões de anos) e A. kadabba (5,8 – 5,2 milhões de anos) partem de um mesmo ancestral em períodos próximos e que Sahelanthropus tchadensis antecede ambos. Muitas das características de Ardipithecus kadabba se aproximam de S. tchadensis. Por exemplo: pélvis e membros inferiores sugerem bipedismo, braços longos e polegar opositor nos pés indicam uma habilidade para escalar ambientes arborícolas; a morfologia das mãos e membros superiores sugerem que Ardipithecus kadabba não era capaz de se locomover de forma quadrúpede, ou seja, apoiando-se em nós dos dedos das mãos (nodopedalia). Apresentam também baixo dimorfismo sexual sugerindo uma monogamia.
O nome do gênero, Ardipithecus, é derivado da língua local de Afar, onde “Ardi” significa “solo” e “Pithecus” refere-se a um primata; Kadabba é um epíteto também vindo da língua local e refere-se ao pai de família. Então, o nome completo refere-se a um primata terrestre.
A descoberta de A. kadabba consiste em um fragmento mandibular do lado direito, um molar (M3) e cinco fragmentos de dente, além de outro osso que havia sido encontrado em 1992 e que só foi catalogado em 2001 com a descrição total das peças encontradas em uma Depressão de Afar, na Etiópia. Na primeira descrição feita por Yohannes Haile-Selassie foi reconhecido como uma sub-espécie de Ardipithecus ramidus. Só em 2004, esta designação de outra espécie foi criada (Haile-Selassie, 2004).
Ardipithecus kadabba mostra também semelhança com Orrorin tugenensis.
O O. tugenensis é a única espécie de hominíneo deste gênero, e esta extinto. O nome foi dado pelos autores da descoberta feita próximo a cidade de Tugen, no Quênia. O grupo, liderado por Martin Pickford, encontrou alguns fósseis no ano 2000. Inicialmente, o autor datou-o em 7 milhões de anos, mas a datação do relógio molecular deixou um resultado mais preciso, 6 milhões de anos (Mioceno).
Depois que os fósseis foram encontrados em 2000, eles foram mantidos no museu Kipsaraman, em sua aldeia, mas o museu foi fechado posteriormente. Desde então, de acordo com presidente do Quênia, Eustace Kitonga, os fósseis são armazenadas em um cofre de banco em segredo em Nairobi (Daily Nation)
Os fósseis encontrados até agora são de, no mínimo, 5 indivíduos. Eles incluem um fêmur, sugerindo que o Orrorin andava de forma ereta; um úmero direito, sugerindo habilidades de escalador, mas não de braquiação; e dentes sugerem uma dieta parecida com a dos humanos modernos. Com molares grandes e os pequenos caninos sugere-se que o Orrorin comia frutas, vegetais e, ocasionalmente, carne. Essa espécie tinha, aproximadamente, o mesmo tamanho que o chimpanzé.
Outros fósseis encontrados nessas rochas mostram que o Orrorin viveu em um ambiente arbóreo, mas não na savana como dito por muitas teorias sobre evolução humana e, em particular, sobre as origens do bipedalismo. Se o Orrorin foi um ancestral do homem moderno, o Australopithecus afarensis estaria em um dos lados do ramo da família dos hominíneos (Senut et al, 2001).
A cabeça do fêmur é esférica, o colo é alongado e oval e o trocânter é menor, se projetando medialmente. Isso sugere que o Orrorin era bípede, e o resto pós-craniano indica que ele subia em árvores. Enquanto a falange proximal é curva, a falange distal é de proporções humanas e, apesar de ter sido sugerido estar associado a fabricação de ferramentas, provavelmente estar mais associado a se prender em árvores (Henke, 2007).
Bernard Wood e Nicholas Lonerga (2008) argumentam que S. tchadensis, A. ramides e A. kadabba são variedades de uma mesma espécie que apresentam características diferentes, mesmo na arcada dentária. Isso pode ser visualizado em outros tipos de Homininae que são substancialmente diferentes em suas características de dentição (Wood & Lonergan, 2008).
O potencial de Sahelanthropus tchadensis
O último ancestral comum chimpanzé-humano, (CHLCA), refere-se á última espécie ancestral comum compartilhada entre de humanos e chimpanzés; ela representa o ponto do nó em que a linha do gênero Homo se separa da linha que deu origem ao gênero Pan. O último ancestral comum de humanos e chimpanzés é estimado em ter vivido durante o final do Mioceno, isto é, a mais de 7 ou 8 milhões de anos atrás.

Reconstrução de Sahelanthropus tchadensis
A especiação de Pan para Homo parece ter sido um processo longo e arrastado. Inicialmente acreditava-se que após a divergência, houve períodos de hibridação entre grupos populacionais e um processo de divergência que durou alguns milhares ou milhões de anos (Patterson et al, 2006). Em algum momento durante o final do Mioceno ou no início do Plioceno os primeiros membros do clado humano completaram uma separação definitiva da linhagem do Pan, algumas estimativas por especialistas variam entre 13 milhões (Arnason et al, 1998) com uma margem de 4 milhões de anos para mais ou para menos (ou seja, 17 ou 9 milhões de anos) (Patterson et al, 2006). A última estimativa, de 9 milhões de anos é a mais aceita. E eventos de hibridação são rejeitados por Wakeley (2008) e as estimativas mais recentes.
Richard Wrangham (2001) argumentou que a espécie CHLCA era muito parecida com o chimpanzé comum (Pan troglodytes), então é assim que ele deve ser classificado, como um membro do gênero Pan e ser dado o nome taxonômico Pan prior (De Waal, 2001). No entanto, até o momento, nenhum fóssil foi identificado como candidato mais provável para a vaga de CHLCA, ou como o táxon Pan prior.
Em estudos genéticos humanos, o CHLCA é útil como um ponto de ancoragem para o cálculo genético de Polimorfismo de nucleotídeo único (SNP, sigla de single nucleotide polymorphism) em populações humanas em que os chimpanzés são utilizados como um grupo externo, que é, a espécie existente geneticamente mais semelhante ao Homo sapiens. O SNP refere-se a uma variação na sequência de DNA que afeta somente uma das 4 bases (adenina, timina, citosina ou guanina) na sequência do genoma. Alguns autores consideram a troca de poucos nucleotídeos, assim como pequenas inserções ou deleções (chamadas de “indel”) como SNPs. Nestes casos, o termo polimorfismo de nucleotídeo simples é mais adequado. Estas variações devem ocorrer em no mínimo 1% de uma determinada população para ser considerada como um SNP. Se, por outro lado, a frequência de uma variação for inferior a 1%, a mesma será considerada simplesmente uma mutação. Os SNPs constituem 90% de todas as variações genômicas humanas e aparecem, em média, uma vez a cada 1.300 bases, ao longo do genoma humano. Dois terços dos SNP correspondem a substituições de uma citosina por uma timina. Além de poder acarretar mudanças morfológicas, essas variações na sequência do DNA podem influenciar a resposta dos organismos a doenças, bactérias, vírus, produtos químicos, fármacos, etc.
A tribo “Hominini” foi proposta com base na ideia de que, ao construir uma filogenia, quando surge uma tricotomia (três espécies surgindo em um mesmo ponto, o que é improvável), a espécie menos semelhante deve estar separada dos outros dois. Originalmente, isto produziu o gênero Homo que, previsivelmente, foi considerado “mais diferente” entre os três gêneros que incluem Pan e Gorilla. No entanto, mais tarde descobertas e análises revelaram que Pan e Homo estão mais próximas geneticamente do que Pan e Gorilla; assim, o gênero dos chimpanzés foi encaminhado para a tribo Hominini com o Homo. Por esta razão podemos considerar humanos, chimpanzés, gorilas e orangotangos como “hominídeos”, mas quando nos referirmos aos fósseis ligados á evolução humana chamamos de “hominínios”, e se incluirmos os chimpanzés a eles, então chamamos de “hominíneos”.

Classificação humana
Gorilla tornou-se um gênero separado e foi encaminhado para uma nova “Gorillini”. Mann e Weiss (1996) propuseram que a tribo Hominini deve abranger Pan, bem como Homo, mas agrupados em subtribos separados (Mann et al, 1996). Richard Wrangham (2001) argumentou que a espécie CHLCA era muito semelhante ao dos chimpanzés, então ele deve ser classificado como um membro do gênero Pan com o nome de Pan prior (De Waal, 2001). Até o presente momento, Sahelantrhopus tchadensis é o candidato potencial para a vaga de CHLCA ou ao táxon Pan prior.
Os descendentes do “lado humano” de CHLCA devem ser especificados como membros da tribo Hominini, isto é, para a inclusão do gênero Homo e seus gêneros estreitamente relacionados (Australopithecus), mas com a exclusão de gênero Pan. Esse agrupamento representa o ”clado humano” e seus membros que são chamados de hominínios (Bradley, 2006). Os descendentes do “lado chimpanzé” refere-se á tribo “Panini”, que foi idealizado a partir da família Hominidae sendo composto por um de trifurcação subfamílias (Wood & Richmond, 2000).
Sahelanthropus tchadensis é a espécie extinta com uma morfologia esperada do CHLCA; e que viveu cerca de 7 milhões de anos, que é muito próximo ao tempo da divergência chimpanzé-humano. Mas não está claro se ele deve ser classificado como um membro da tribo Hominini, ou seja, um hominídeo, ou como um ancestral direto do Homo e Pan e um candidato potencial para a própria espécie CHLCA. Alguns espécimes fósseis no “lado chimpanzé” foram encontrados; o primeiro chimpanzé fóssil data entre 545 e 284 mil anos e foi descoberto em Leste Africano Rift Valley, no Quênia (McBrearty, 2005) (McBrearty et al, 2005).
Outros gêneros extintos listados foram ligados ao ancestral de Homo, ou são desdobramentos paralelos. Apenas Orrorin e Sahelanthropus, existiam na época da divergência entre chimpanzés e nossa linhagem, e assim, um ou ambos podem ser ancestral tanto do gênero Homo quanto de Pan.
Uma reconstrução virtual detalhado do crânio TM 266 de S. tchadensis corrigiu as distorções causadas pela fossilização. Uma tomografia computadorizada de alta resolução foi usada para criar uma representação digital do crânio ao longo das principais fissuras e foi avaliada em diversos testes independentes. Em primeiro lugar, a face e o complexo neuro-base-cranial foram reconstituídas em separados, em um plano ao longo das margens superior e laterais da região pós-orbital. Em segundo lugar, a morfologia foi avaliada e reconstruída e comparada contra uma restrição anatômica não considerada durante a reconstrução virtual.
A reconstrução confirma que S. tchadensis é um hominídeo e não está intimamente relacionada com os grandes primatas africanos (Wolpoff et al, 2002). A análise da base do crânio indica que S. tchadensis eram um bípede ereto, o que sugere que o bipedalismo estava presente no mais antigo membro conhecido dos hominídeos e provavelmente surgiu logo após a divergência da linhagem chimpanzé e humana.

Reconstrução do crânio de Sahelanthropus tchadensis
Victor Rossetti
Palavras chave: NetNature, Rossetti, Sahelanthropus tchadensis, Orrorin tugenensis, Ardipithecus kadabba, evolução humana, Chimpanzé, Humanos, Último ancestral comum, Fósseis, Crânio, Fêmur
.
Referências
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.