
Equinodermos
Capítulo por:
Jaleigh Q. Pier e Jansen A. Smith, Instituto de Pesquisa Paleontológica, Ithaca, Nova York.
Uma versão inicial deste capítulo foi compartilhada publicamente pela primeira vez em 20 de dezembro de 2019; o capítulo completo foi compartilhado pela primeira vez em 24 de julho de 2020.
Este capítulo foi atualizado pela última vez em 24 de julho de 2020.
Citação do capítulo:
JQ Pier e JA Smith. 2019. Equinodermos. In: A Enciclopédia Digital da Vida Antiga. https://digitalatlasofancientlife.org/learn/echinodermata
Conteúdo do capítulo:
Echinodermata ←
–– 1. Taxa Exclusivamente Fóssil
–– 2. Crinoidea
–– 3. Asteroidea
–– 4. Ophiuroidea
–– 5. Echinoidea
–– 6. Holothuroidea
Materiais associados:
Coleção de ensino virtual de modelos de fotogrametria 3D de fósseis de Echinodermata disponíveis aqui!
Acima da imagem: Espécimes de Echinodermata preservados recentes. Imagem de Jonathan R. Hendricks de espécimes de equinodermos modernos exibidos na Academia de Ciências Naturais da Universidade de Drexel, licenciados sob uma Licença Creative Commons Atribuição-Compartilhamento pela mesma Licença 4.0 Internacional.
Instantâneo do Filo Echinodermata
- Classes vivas: Crinoidea, Asteroidea, Ophiuroidea, Echinoidea, Holothuroidea
- Classes extintas exemplares: Blastoidea, Ctenocystoidea, Diploporita, Edioasteroidea, Eocriniodea (até 28 classes exclusivamente fósseis foram identificadas)
- Diversidade: ~ 10.995 espécies existentes, 30.325+ espécies extintas
- Ecologia: exclusivamente marinho (estenohalina), herbívoro, carnívoro, detritívoro, filtrador
- Principais características do grupo: simetria em 5 partes, estereomo, endoesqueleto de ossículos ou placas de CaCO3, sistema vascular de água com pés tubulares
- Registro Fóssil: Cambriano a Recente
Visão geral
“Entre os equinodermos encontramos algumas das mais belas de todas as criaturas marinhas.” (Nichols, 1967, pg. 15)
O filo Echinodermata representa o maior filo animal de animais exclusivamente marinhos e inclui animais tão morfologicamente diversos como estrelas do mar (Asteroidea), estrelas quebradiças (Ophiuroidea), pepinos do mar (Holothuroidea), ouriços do mar e dólares de areia (Echinoidea) e lírios do mar ( Crinoidea). O nome do filo é derivado do grego "echinos", que significa espinhoso, e "derma", que significa pele. Os equinodermos são deuterostômios e, portanto, estão mais relacionados aos cordados (que incluem nós mesmos) do que a quase todos os outros filos de invertebrados.
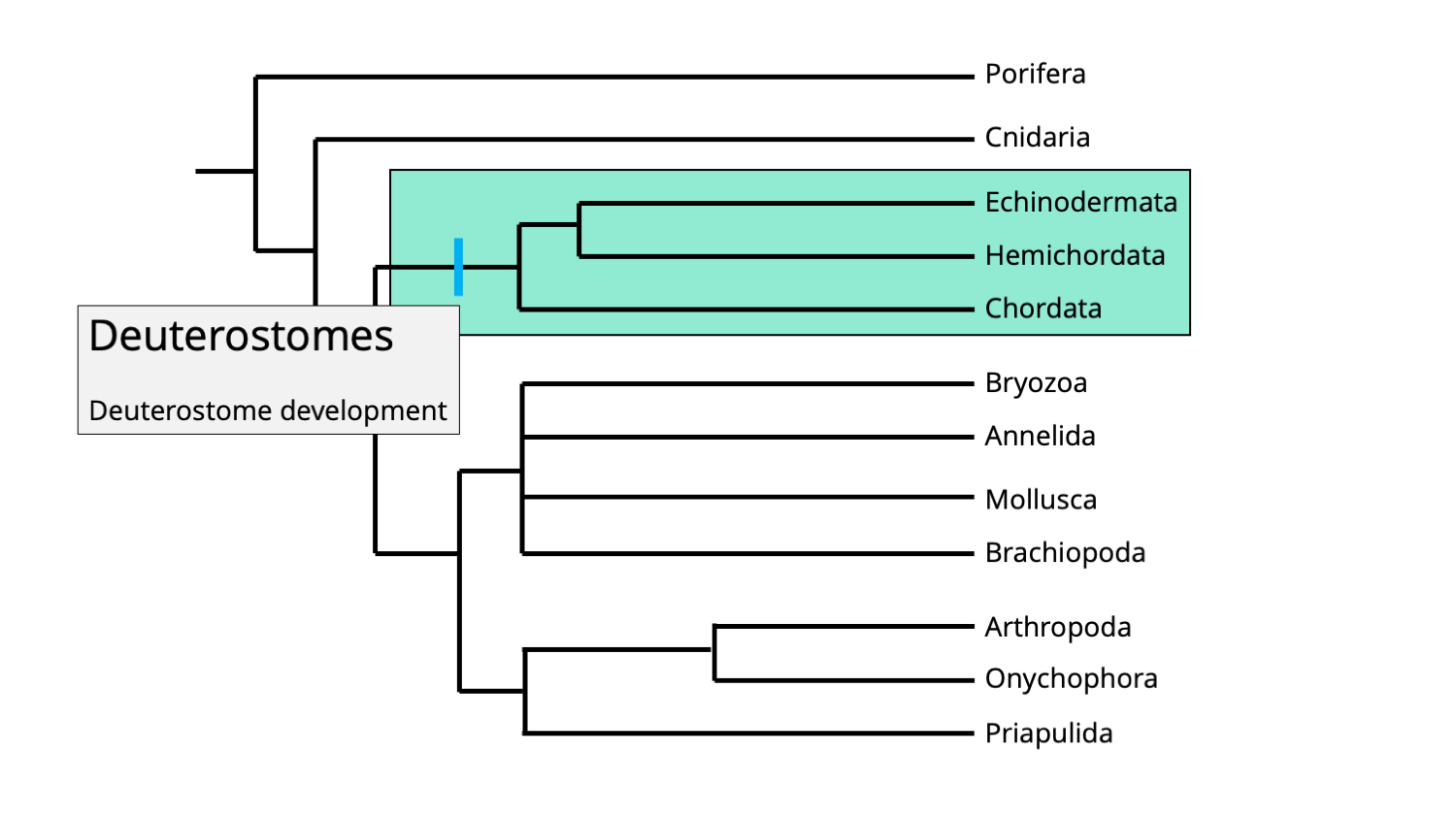
Posições filogenéticas do clado bilateriano Deuterostomia, caracterizada pelo desenvolvimento do ânus e depois da boca durante o desenvolvimento embrionário. Imagem de Jonathan R. Hendricks, licenciada sob uma Licença Creative Commons Atribuição-Compartilhamento pela mesma Licença 4.0 Internacional.
Equinodermos adultos carecem de uma região sensorial da cabeça; no entanto, eles têm um anel nervoso e possuem várias formas de percepção sensorial. Alguns grupos têm um sistema digestivo simples e unidirecional, mas isso não os impediu de ocupar uma grande variedade de modos de alimentação. Eles são um dos grupos de invertebrados mais bem sucedidos e algumas classes (especialmente crinóides e equinóides) têm registros fósseis substanciais.
'Echinoderms de uma história de sucesso' por: Shape of Life ( YouTube ).
Filogenia
Os equinodermos são um grupo diversificado com uma variedade de formas corporais, dificultando a compreensão de como as várias classes estão relacionadas. Os primeiros equinodermos surgiram no Cambriano, com muitas outras classes evoluindo até o final do Ordoviciano, incluindo todas as classes existentes (saiba mais abaixo na seção sobre equinodermos fósseis).
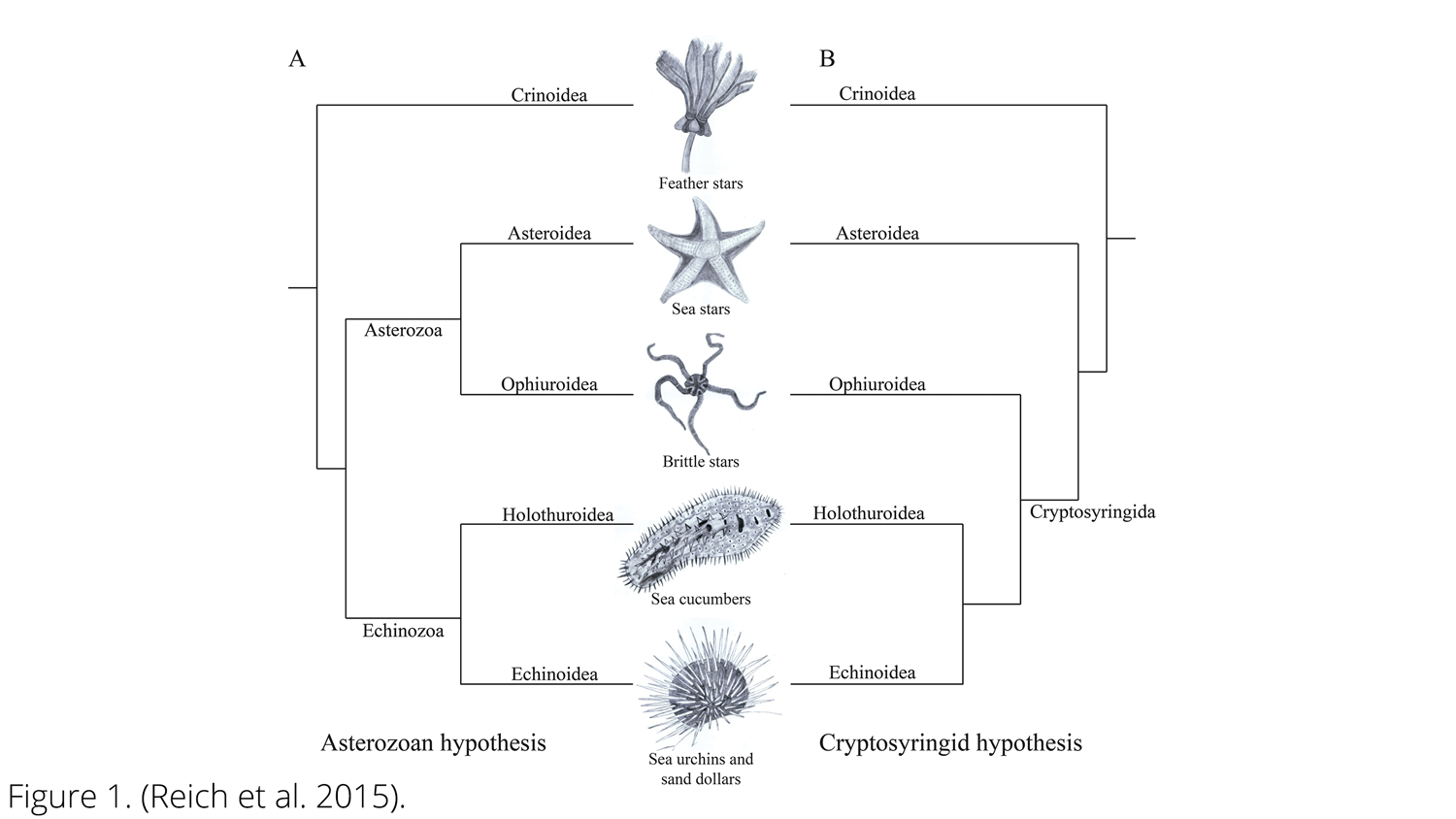
Imagem de hipóteses filogenéticas de equinodermos concorrentes. A: Hipótese de asterozoários B: Hipótese de criptosiringídeos. ( Reich et al., 2015 ; Creative Commons Attribution-ShareAlike 4.0 International License ).
Várias hipóteses concorrentes para explicar a filogenia dos equinodermos foram desenvolvidas ao longo dos anos, incluindo duas hipóteses contrastantes para as relações entre as cinco classes vivas. Em uma delas, chamada de hipótese Asterozoa, Ophiuroidea e Asteroidea são táxons irmãos e formam um clado chamado Asterozoa, enquanto Holothuroidea e Echinoidea formam um clado chamado Echinozoa (com Crinoidea irmã de Asterozoa + Echinozoa). Na segunda, chamada de hipótese Cryptosyringid, Ophiuroidea é colocado como o táxon irmão de Holothuroidea + Echinoidea. Análises filogenéticas moleculares recentes ( Reich et al ., 2015 ; Telford et al ., 2014 ) apoiaram a hipótese de Asterozoários, que também favorecemos aqui.
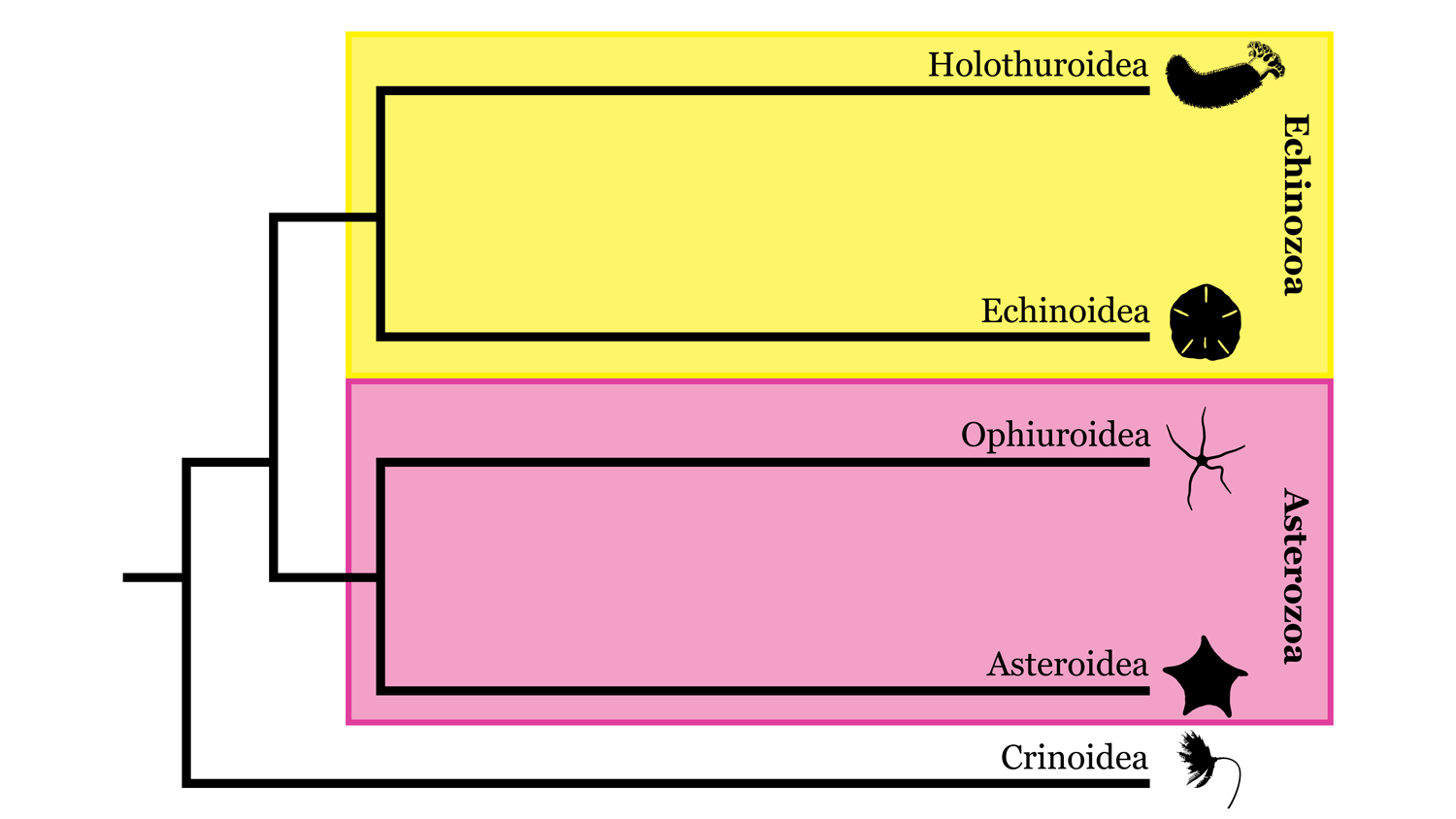
Visão geral altamente simplificada da filogenia de Echinodermata baseada em parte na hipótese de relacionamentos apresentada por Reich et al. (2015). Imagem por: Jaleigh Q. Pier, licenciado sob uma Licença Creative Commons Atribuição-Compartilhamento pela mesma Licença 4.0 Internacional.
Características gerais
Simetria Pentaradial
Todas as formas adultas de equinodermos modernos têm 5 pontos, ou simetria pentarradial, o que significa que eles têm características morfológicas que são divisíveis por cinco. Por exemplo, as estrelas do mar têm cinco braços (ou alguns múltiplos de cinco braços) e os endosqueletos calcários (ou, testes) de ouriços-do-mar podem ser divididos de forma semelhante em quintos (veja a imagem abaixo). Da mesma forma, os dólares de areia têm cinco "pétalas" em seu lado dorsal. E os estranhos pepinos do mar? Se você pensar em um pepino do mar como um "teste" equinóide alongado, eles também possuem simetria pentarradial. É importante notar, no entanto, que os equinodermos larvais geralmente têm simetria bilateral (uma sinapomorfia para animais bilaterais; veja aqui). Além disso, embora todas as classes existentes de equinodermos tenham simetria pentarradial, algumas formas fósseis extintas do início do Paleozóico não tinham.
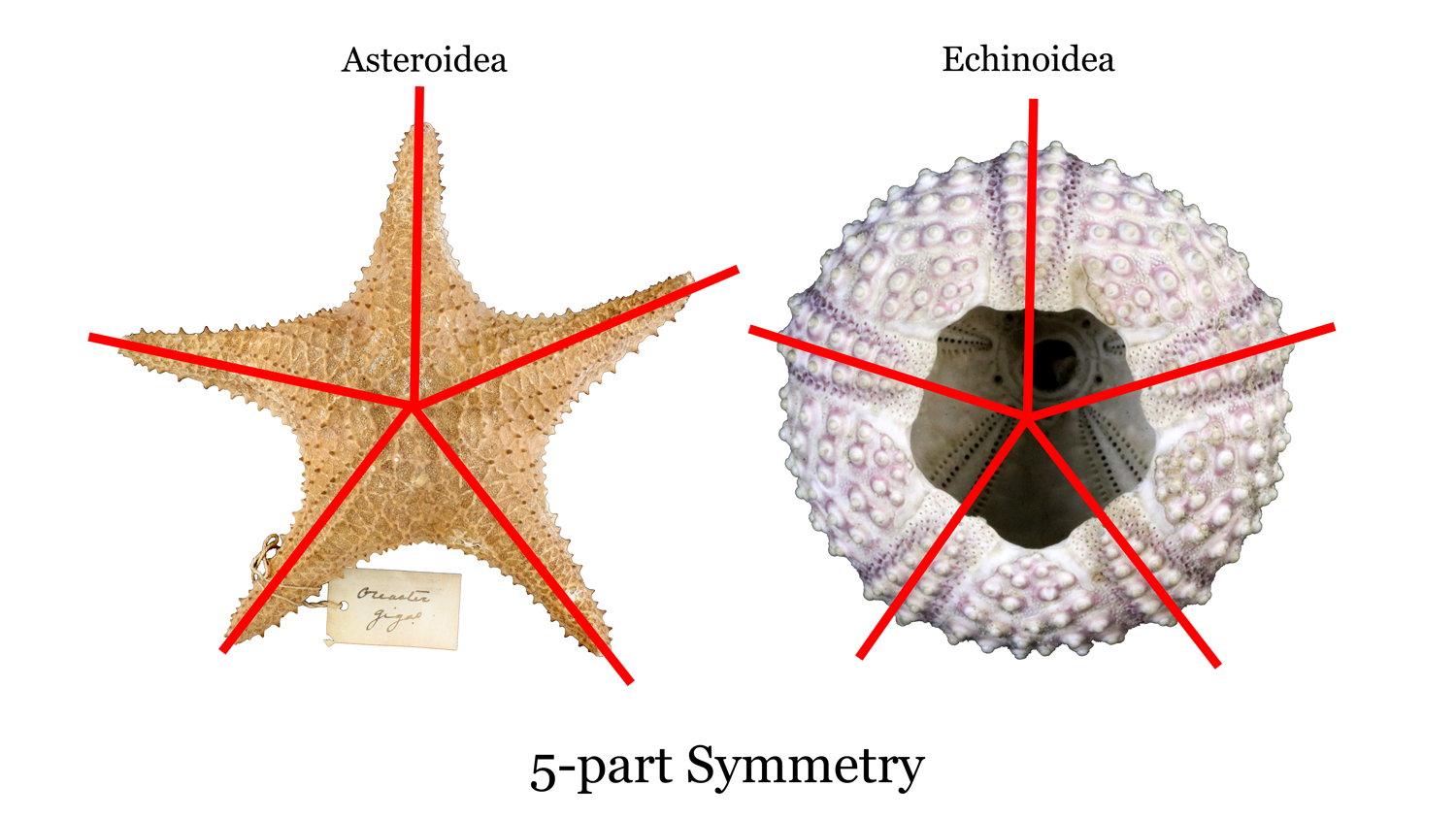
Imagem de simetria pentarradial de Asteroidea e Echinoidea. Imagem por: Jaleigh Q. Pier, licenciado sob uma Licença Creative Commons Atribuição-Compartilhamento pela mesma Licença 4.0 Internacional.
Se o conceito de simetria pentarradial parece difícil de entender, assista ao vídeo abaixo, que destaca as diferentes simetrias de 5 partes das principais classes de equinodermos.
'Animação de equinoderme: simetria em cinco partes' por: Shape of Life
Endoesqueleto de ossículos e tecido colagenoso
Os equinodermos possuem um verdadeiro endoesqueleto formado por ossículos (ou seja, pequenas placas rígidas) de carbonato de cálcio com microestrutura estereoscópica que atuam como suporte do corpo, locais de fixação do tecido conjuntivo e proteção. O esqueleto ressecado é chamado de "teste" (por exemplo, equinóides), "teca" (por exemplo, blastóides) ou "cálice" (por exemplo, crinóides). Holothuroideans (pepinos do mar) são compostos principalmente de tecido colágeno e seus ossículos são de tamanho microscópico. Os ossículos são conectados por músculos e ligamentos, que são feitos principalmente da proteína colágeno. Os equinodermos têm tecido colagenoso mutável especializado, permitindo-lhes alterar a rigidez de seu esqueleto. Por exemplo, eles podem se tornar muito duros e rígidos para se proteger de predadores ou macios e flexíveis ao se mover. O movimento é alcançado de várias maneiras, incluindo o uso dos pés tubulares do sistema vascular aquático, contrações ligamentares para movimentos lentos e contrações musculares de movimento rápido.

Imagem de testes de equinoderme (Ophiuroidea, Asteroidea, Echinoidea) vs equinoderma teca (Blastoidea e Crinoidea). Imagem por: Jaleigh Q. Pier, licenciado sob uma Licença Creative Commons Atribuição-Compartilhamento pela mesma Licença 4.0 Internacional.
Sistema Vascular de Água
O sistema vascular da água é importante para quase todas as funções vitais dos equinodermos modernos, incluindo movimento, alimentação e respiração. A pressão da água é controlada por este sistema para operar os pés do tubo, que são encontrados em todas as classes de equinodermos existentes. Esses apêndices com ventosas são permeáveis para permitir a transferência de oxigênio.
Normalmente, a pressão da água é controlada através do madreporito, que se conecta internamente ao canal de pedra e depois ao canal do anel. Numerosos canais radiais se estendem a partir do canal do anel, muitas vezes refletindo a simetria de 5 partes do plano corporal do equinodermo. Os pés tubulares se estendem dos canais radiais e se movem por meio de contrações musculares e relaxamentos das ampolas.
Regeneração
Uma característica fascinante de todos os equinodermos é sua capacidade de regenerar partes do corpo perdidas. Tanto as estrelas do mar quanto as estrelas quebradiças podem regenerar os braços e, em alguns casos, dividir-se ao meio para produzir dois novos indivíduos. Algumas espécies de pepino-do-mar também são capazes de se dividir ao meio para criar novos indivíduos e também podem regenerar todo o seu trato digestivo, que podem ejetar ativamente (mais sobre isso na página da classe Holothuroidea !). Os ouriços-do-mar são capazes de substituir os espinhos e os pés tubulares. Cada um desses métodos de regeneração será discutido nas páginas da classe de equinodermos.

Fotografia de uma estrela do mar regenerando pelo menos sete braços (raios menores, de cor branca). Imagem de: Jonathan R. Hendricks, licenciado sob uma Licença Creative Commons Atribuição-Compartilhamento pela mesma Licença 4.0 Internacional.
Registro fóssil
Hoje, Echinodermata é um filo diversificado, com inúmeras espécies em cinco classes: Asteroidea (peixe estrela), Crinoidea (lírios do mar e estrelas de pena), Echinoidea (ouriços do mar e dólares de areia), Holothuroidea (pepinos do mar) e Ophiuroidea (estrelas quebradiças ). ). No registro fóssil, essas classes são frequentemente representadas por apenas pedaços de seus esqueletos, que se desfazem rapidamente após a morte porque os vários pedaços são mantidos juntos por tecidos moles que se decompõem. As peças encontradas incluem diferentes componentes dependendo da classe:
- Crinoidea : ossículos do caule desarticulados mais comumente preservados, caule e braços intactos preservados em condições excepcionais.
- Asteroidea : placas ossiculares e endoesqueleto comumente preservados, especialmente como moldes.
- Ophiuroidea : disco central e ossículos com maior probabilidade de serem preservados intactos, braços e corpo fóssil preservados em condições excepcionais
- Echinoidea : espinhos soltos e teste mais comumente preservado, fósseis de corpos em condições excepcionais.
- Holothuroidea : raramente preservado, porém ossículos soltos podem ser comuns.
O filo foi representado exclusivamente por essas cinco classes desde o Triássico. No entanto, o registro fóssil de equinodermos inclui até 28 classes adicionais, que se originaram e se extinguiram durante o Paleozóico (veja alguns exemplos na imagem abaixo).
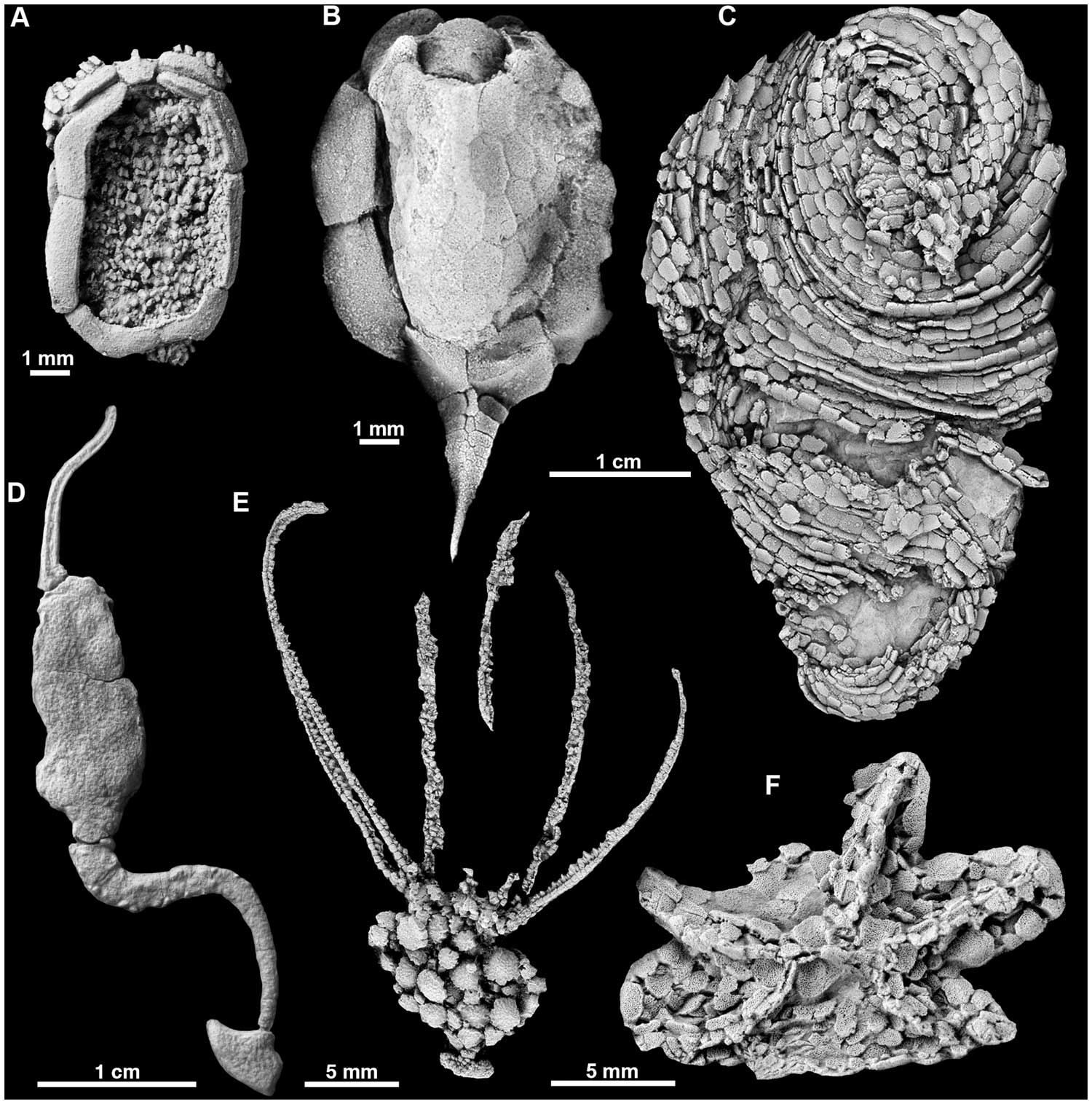
"Equinodermos radiados e assimétricos do Cambriano mostrando uma seleção de planos corporais de equinodermos primitivos. A , o ctenocistoide Ctenocystis ; B , o cinctan Gyrocystis ; C , o helicoplacoide Helicoplacus ; D , o soluto Coleicarpus ; E , o eocrinóide Gogia ; F , estromatocistide erioasteróide." Figura e legenda de Zamora et al. (2012) em PloS ONE ; Licença Creative Commons Attribution-Share Alike 4.0 International .
A evolução dos equinodermos pode ser pensada em quatro fases, conforme descrito por Ausich e Sumrall (2016) na 9th European Conference on Echinoderms . A primeira fase da história evolutiva dos equinodermos começa com a evolução dos planos corporais dos equinodermos durante o Cambriano – alegações putativas de equinodermos do Ediacarano (por exemplo, Arkarua) permanecem especulativos, na melhor das hipóteses. Como evidenciado no registro fóssil, os primeiros equinodermos exibiram uma ampla gama de diversidade morfológica, incluindo formas com planos corporais helicoidais, laterais e pentarradiais. A segunda fase é marcada pela diversificação de espécies com todos os três planos corporais durante o Cambriano, incluindo formas pentarradiais com e sem hastes. Essa diversificação resultou no primeiro pico na diversidade de equinodermos em nível de classe, com até 15 classes conhecidas da porção posterior do Cambriano.

Diversidade em nível de classe de equinodermos ao longo do tempo com fases importantes da evolução dos equinodermos indicadas. Figura da apresentação de Ausich e Sumrall (2016) na 9ª Conferência Europeia sobre Equinodermos , refletindo o trabalho do projeto Assembling the Echinoderm Tree of Life . Incluído com permissão de W. Ausich.
Várias classes de equinodermos foram perdidas no final do Cambriano, mas a diversificação em nível de classe durante a terceira fase da evolução dos equinodermos resultou em até 21 classes durante o Ordoviciano. Foi durante este tempo que as cinco classes existentes evoluíram pela primeira vez. Do final do Ordoviciano até o final do Permiano, o número de classes de equinodermos diminuiu constantemente. Além das numerosas classes paleozóicas extintas, a extinção em massa do final do Permiano quase eliminou os crinóides e equinóides, com algumas evidências sugerindo que apenas uma família de cada classe sobreviveu a esse evento catastrófico. A quarta fase da evolução dos equinodermos começou no Triássico e é definida como a diversificação dentro das cinco classes remanescentes existentes.
Conforme brevemente apresentado aqui, nem todas as diversas morfologias de equinodermos evoluíram ao mesmo tempo, sugerindo que as características definidoras deste filo, e potencialmente de outros filos, não foram gravadas em pedra, por assim dizer, na primeira aparição no registro fóssil. . Em vez disso, diferentes classes surgiram ao longo de milhões de anos, com a extinção diferencial filtrando os grupos que podemos considerar incomuns e cientificamente fascinantes hoje (veja a imagem acima). Por causa de seu registro fóssil limitado, as relações filogenéticas entre esses grupos permanecem difíceis de reunir (veja uma hipótese na imagem abaixo). Compreender essas relações tem sido o foco de um grande projeto de pesquisa colaborativo chamado Assembling the Echinoderm Tree of Life— visite a página deles para ver o progresso feito recentemente!
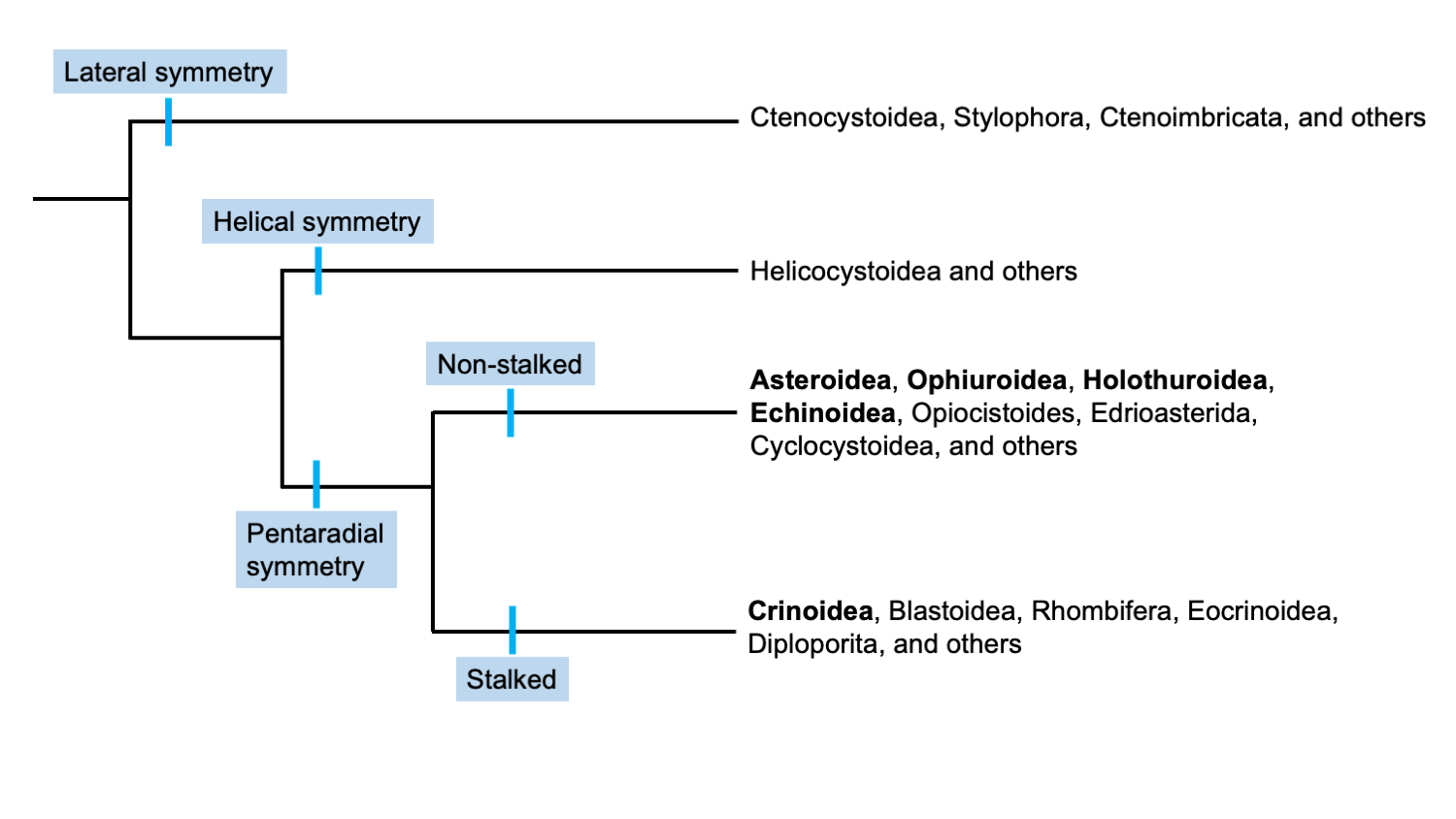
Uma hipótese sobre a relação entre diferentes grupos de equinodermos com base na simetria. Observe que as aulas vivas estão em negrito. Figura modificada da apresentação de Ausich e Sumrall (2016) na 9th European Conference on Echinoderms , refletindo o trabalho do projeto Assembling the Echinoderm Tree of Life . Incluído com permissão de W. Ausich.
Ao avaliar a evolução e desenvolvimento desse filo, Deline et al. (2020) identificaram quatro morfologias gerais de equinodermos: (1) não radial; (2) crinóides radiais pedunculados; (3) blastozoários radiais pedunculados; e, (4) pentarradial diretamente acoplado e móvel. Para os propósitos deste capítulo, consideraremos o segundo e o terceiro grupos juntos (isto é, radial com haste), pois eles são muito semelhantes em aparência e provavelmente formam um clado (isto é, Pelmatozoa). Nas páginas que se seguem, vamos dar uma olhada mais de perto nas classes existentes e, primeiro, uma seleção de classes exclusivamente fósseis dentro desses três grupos morfológicos: não-radiais, radiais pedunculados (crinóides e blastozoários) e pentarradiais fixos e móveis.
Agradecimentos
Agradecemos a Bill Ausich, Jen Bauer e Sarah Sheffield por fornecerem feedback sobre as versões anteriores deste capítulo para melhorar sua qualidade.
Referências e leitura complementar:
Boardman, RS, AH Cheetham e AJ Rowell. 1987. Invertebrados Fósseis. Publicações Científicas Blackwell. 713 p.
Deline, B., JR Thompson, NS Smith, S. Zamora, IA Rahman, SL Sheffield, WI Ausich, TW Kammer e CD Sumrall. 2020. Evolução e Desenvolvimento na Origem de um Filo. Current Biology , 30: 1672-1679 .
Nichols, D., 1967. Equinoderms. Biblioteca da Universidade Hutchinson, Londres.
Reich, A., C. Dunn, K. Akasaka, G. Wessel. 2015. Análises filogenômicas de Echinodermata apoiam os grupos irmãos de Asterozoa e Echinozoa. PLoSONE, 10: e0119627.
Smith, AB, S. Zamora e JJ Álvaro. 2013. As mais antigas faunas de equinodermos de Gondwana mostram que a diversificação do plano corporal dos equinodermos foi rápida. Nature Communications , 4: 1-7.
Telford, MJ, CJ Lowe, CB Cameron, O. Ortega-Martinez, J. Aronowicz, P. Oliveri e RR Copley. 2014. A análise filogenómica de relações de classe de equinodermos suporta Asterozoa. Anais da Royal Society B: Ciências Biológicas, 281: 20140479 .
Wright, DF, WI Ausich, SR Cole, ME Peter e EC Rhenberg. 2017. Taxonomia filogenética e classificação dos Crinoidea (Echinodermata). Journal of Paleontology , 91: 829-846 .
Zamora, S., B. Deline, JJ Álvaro, and IA Rahman. 2017. A Revolução Cambriana do Substrato e a evolução inicial da fixação em equinodermos que se alimentam de suspensão. Earth-Science Reviews , 171: 478-491 .
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.