Decifrando a diversidade dos hominídeos africanos do Pleistoceno Médio e a origem de nossa espécie
Nature Communications volume 10 , Número do artigo: 3406 ( 2019 )
Abstrato
A origem do Homo sapiens permanece uma questão de debate. A extensão e o padrão geográfico da diversidade morfológica entre os hominídeos africanos do Pleistoceno Médio Superior (LMP) são amplamente desconhecidos, impedindo assim a definição de limites de variabilidade no início do H. sapiens e a interpretação de fósseis individuais. Aqui usamos um método de modelagem filogenética para prever possíveis morfologias de um último ancestral comum de todos os humanos modernos, que comparamos com fósseis africanos LMP (KNM-ES 11693, Florisbad, Irhoud 1, Omo II e LH18). Nossos resultados apóiam um processo complexo para a evolução do H. sapiens , com o reconhecimento de diferentes populações e linhagens geograficamente localizadas na África – nem todas contribuíram para a origem de nossa espécie. Com base nos fósseis disponíveis, o H. sapiens parece ter se originado da coalescência das populações de origem do sul e, possivelmente, do leste africano, enquanto os fósseis do norte da África podem representar uma população que introgrediu os neandertais durante o LMP.
Introdução
A história e evolução do gênero Homo tem sido objeto de contínuos debates 1 , 2 . Descobertas recentes de fósseis 3 , 4 , novas técnicas analíticas 5 , 6 e grandes desenvolvimentos na genômica antiga 7 , 8 melhoraram consideravelmente nossa compreensão da evolução de nosso gênero, modificando profundamente o arcabouço teórico paleoantropológico. Portanto, os modelos para a origem de nossa espécie, H. sapiens , afastaram-se do confronto de dois pontos de vista antagônicos extremos: (1) o Modelo Multirregional das origens humanas modernas implicando a evolução gradual das populações globais de hominídeos arcaicos em direção a um moderno morfologia humana ao longo dos últimos 2 milhões de anos; 9 , 10 e (2) a hipótese de um único evento fora da África 1 , também conhecido como modelo de Origem Africana Recente (RAO), implicando uma única origem africana (200.000–100.000 anos atrás (ka)) de humanos modernos e subsequentes substituição de todas as populações arcaicas. Modelos subsequentes para abordar a complexa geografia evolutiva dos hominídeos do Quaternário tardio foram propostos, incluindo múltiplas dispersões fora da África e o papel da estrutura populacional na África nos processos de diversificação 11 . Juntamente com a compreensão dos mecanismos evolutivos que deram origem aos neandertais na Europa 12 e um número crescente de estudos genéticos 13 , 14 , esses modelos abriram caminho para uma mudança de ênfase da substituição para os resultados demográficos e genéticos potencialmente variados de interações recentes com hominídeos 7 , 9 , 15 , 16 , 17 .
No entanto, muitas questões permanecem sem resposta, em parte devido à escassez de fósseis africanos do Pleistoceno Médio Superior (LMP, de 350 a 130 ka), o que é uma grande restrição para qualquer estudo do registro fóssil africano do LMP. No norte da África, o sítio de Jebel Irhoud produziu vários fósseis desde a década de 1960 18 , incluindo um crânio completo (Irhoud 1), originalmente datado de 130–190 ka 19 . Escavações recentes no local renderam fósseis adicionais (em particular uma face superior parcial, Irhoud 10, e uma mandíbula, Irhoud 11), e uma nova estimativa de data de 315 ka 20 . Os hominídeos LMP bem preservados são mais numerosos na África Oriental. O calvário de Singa do Sudão é datado de 133 ka 21 . Na Etiópia, os espécimes de Omo Kibish, Omo I e Omo II 22 , são datados de 200 ka 23 , e os três espécimes de Herto, que incluem um crânio adulto completo (BOU-VP16/1) e uma calvária juvenil (BOU-VP16 /5), com data estimada de 160 ka 24 . No Quênia, a calvária de Guomde (KNM-ER 3884), que não possui a maior parte dos ossos faciais e frontais 25 , foi datado em 270–300 ka com espectrometria de raios ɤ 26 , enquanto uma idade de 200–300 ka foi sugerida para o crânio quase completo de Eliye Springs (KNM-ES 11693 27 ) com base em sua morfologia 28 . Mais ao sul, um crânio de 200–300 ka (LH18 29 , 30 ) foi descoberto em Ngaloba Beds em Laetoli (Tanzânia) e na África do Sul, o local de Florisbad produziu um crânio parcialmente preservado datado de 259 ka 31 . Por último, os restos recentemente descobertos de H. naledi , datados de 236–335 ka 32 , adicionam maior complexidade ao registro de hominídeos LMP da África Austral.
Mesmo excluindo o material de H. naledi , os fósseis africanos de LMP exibem morfologias extremamente variáveis. Os espécimes Omo I 22 e Herto 24 possuem uma anatomia de tipo moderno que inclui a presença das duas apomorfias crânio-mandibulares da espécie - proporções cranianas que resultam em uma abóbada alta (altura basi-bregmática) e um queixo, e são geralmente considerados os primeiros restos indiscutíveis de H. sapiens 16 , 17 . Todos os outros fósseis africanos LMP mostram um mosaico de caracteres derivados e arcaicos. Por exemplo, os restos de Jebel Irhoud foram originalmente descritos como mostrando fortes semelhanças com os Neandertais 33 , enquanto o estudo dos novos restos de Irhoud enfatiza suas afinidades com H. sapiens , apesar da ausência de apomorfias humanas modernas importantes (ou seja, abóbada alta e globular, e queixo T invertido) 18 . Os espécimes Guomde 25 , Ngaloba 30 , Eliye Springs 27 e Florisbad 34 , juntamente com Omo II 22 e possivelmente o patológico calvário de Singa 35 , têm sido referidos principalmente como 'arcaico H. sapiens ', uma categoria que agrupa fósseis isolados com morfologias díspares. Esta situação desafia qualquer tentativa de identificar os mecanismos evolutivos que possam explicar o padrão morfológico no registro fóssil africano LMP, bem como identificar a população ancestral, ou populações, dos humanos modernos.
Aqui, usamos um método de modelagem filogenética 36 para estimar estatisticamente a morfologia craniana completa dos hipotéticos Últimos Antepassados Comuns (vLCAs) virtuais para todos os humanos modernos com base em duas filogenias simplificadas do gênero Homo (Fig. 1 , Tabela 1 , Tabelas Suplementares 1 e 2 e Suplementar Fig. 1 ), e através deste fóssil africano LMP virtual, explore a diversidade morfológica dos cinco hominídeos LMP africanos reais mais completos para avaliar quantitativamente como as populações de onde esses fósseis foram retirados podem ter desempenhado um papel na origem do H. sapiens .
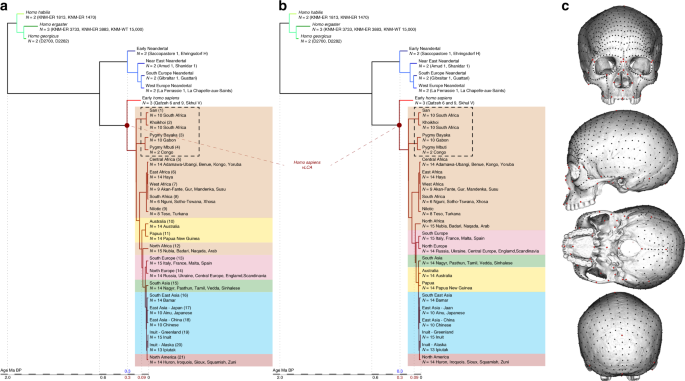
Hipóteses filogenéticas e distribuição de marcos no crânio. aeb totalmente b resolvida do gênero Homo de acordo com as hipóteses 1 ( a ) e 2 ( Filogenia ). c Posição dos 780 marcos e semimarcos usados no estudo para descrever os crânios da amostra filogenética
Nossos principais resultados indicam um processo complexo para a evolução do H. sapiens . Enquanto Florisbad compartilha a maioria das afinidades fenotípicas com os vLCAs computados, alguns dos fósseis LMP apresentam um perfil fenotípico diferente que suporta o reconhecimento de várias populações e linhagens de hominídeos africanos. Nem todas essas linhagens contribuíram igualmente para a origem do H. sapiens e nossos resultados tendem a apoiar a visão de que a origem do H. sapiens pode ser o resultado da coalescência das populações de origem do sul e, possivelmente, do leste africano. Nesse cenário, os hominídeos norte-africanos podem representar uma população que introgrediu os neandertais durante a LMP.
Results
Phylogenetic modelling
Figure 2 presents the phylo-morphospace and the computed position of the ancestral nodes of hypothesis 1 (black tree) and hypothesis 2 (grey tree) of the Homo phylogeny based on 84.6% of the total variation in the data (PC1 to PC3, Supplementary Table 3). Overall, the PCs discriminate accurately the different clades of the phylogenies used. The early Homo species (H. georgicus, H. ergaster, and H. habilis s.l.) and the Neandertals have negative values along PC1 (65.6%), contrasting with H. sapiens specimens. The associated shape deformation shows a low, elongated calvarium with a strongly projecting face for negative PC1 values, while the positive values are associated with a gracile morphology showing a high, rounded calvarium and an orthognathic face compatible with the San and Khoikhoi populations (#1 and 2). PC2 (13.2%) discriminates mainly the early Homo species, which occupy the upper left part of the chart, from modern humans and Neandertals in the lower part of the morphospace. Accordingly, PC2 negative values show a typical Neandertal shape (occipital bun, mid-facial prognathism), whereas PC2 positive values correlate with a small rounded calvarium, and a strongly-projecting face. The phenotypical variation within the modern human cluster is mostly explained by PC3 (5.8%) and follows closely the topology of hypothesis 1. The early H. sapiens group is morphologically close to the H. sapiens vLCAs and fits within the 95% confidence envelope computed around them. All sub-Saharan populations present positive values (#1 to 9), as well as the Oceanians (#10 and 11) and the North Africans (#12). The Khoisans (#1 and 2) are slightly isolated on PC1, showing the most gracile morphology, while the Pygmies (#3 and 4) have the most positive values on PC3. In the negative values of PC3, South Asians (#15) stand as intermediate between Africans-Oceanians and Eurasians, Europeans (#13 and 14) cluster with South-East Asians (#16) and East-Asians (#17 and 18), and Inuits (#19 and 20) are relatively close to North Americans (#21). The computed H. sapiens vLCAs are almost identical for both hypotheses and are more distant from the ancestral node hypothesising the common ancestry between modern humans and Neandertals than the Neandertal ancestor (Fig. 2).
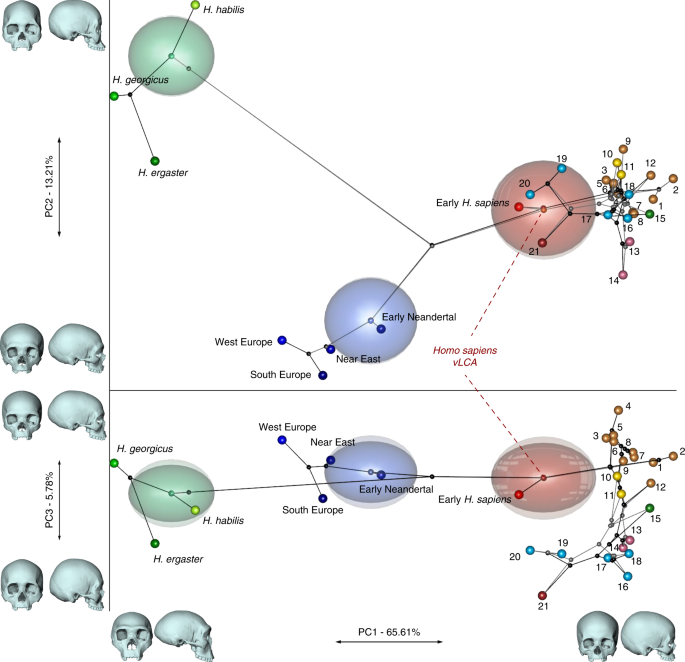
Projeção das hipóteses filogenéticas 1 (preto) e 2 (cinza) no morfoespaço. As deformações de forma associadas são exibidas ao lado de cada PC. Cada nó representa as formas dos ancestrais estimados junto com envelopes de confiança de 95%. Ambas as árvores são semelhantes em PC1 e 2, enquanto PC3 destaca as diferenças entre ambas as hipóteses dentro do clado humano moderno. Populações humanas modernas como segue: 1 a 9 África Subsaariana; 10–11 Oceania, 12 Norte da África, 13–14 Europa, 15 Sul da Ásia, 16 a 20 Leste Asiático, 21 América do Norte (ver Fig. 1 , Fig. 1 complementar e Tabela 1 complementar ). Os dados de origem são fornecidos como um arquivo de dados de origem
The differences between hypotheses 1 and 2 are subtle. However, the phylogenetic signals37,38 computed for hypothesis 1 appear stronger than the ones computed for hypothesis 2 (Supplementary Table 4). This may indicate a better fit of the phenotypic data to phylogenetic hypothesis 1, which includes an earlier wave of dispersal from Africa to Eurasia that preserves a phenotypic signal in Oceania39, supported by the similarity of Australians and Papuans with sub-Saharan Africans. The Khoisans and Pygmies branch from the main modern human populations close to the H. sapiens LCA13,40, but are anatomically derived and gracile41. Removing these two groups from the modelling (hypotheses 1b and 2b, Fig. 1) does not alter the coherence of the distribution of the main clades (early Homo, Neandertals and H. sapiens) or the topologies of both hypotheses in the phylo-morphospace (PC1 to PC3, 85.9%, Supplementary Table 5, Supplementary Fig. 2). However, hypothesis 2b (grey) generates differences impacting on most of the nodes of the tree, resulting in vLCA2b being closer to current modern humans than vLCA1b, and the Australian/Papuan phenotype requiring longer branches between ancestral nodes and terminal taxa to be fitted to the hypothesised topology. This is demonstrated by the computed phylogenetic signals for both hypotheses (Supplementary Table 4).
Morphologies of the vLCAs
As morfologias de vLCA1 e 2 são virtualmente as mesmas (Fig. 3a , desvio de superfície <0,16 mm, Tabela Suplementar 6 ). Ambos são relativamente graciosos em comparação com os fósseis africanos LMP; vLCA1 é um pouco mais robusto, com um frontal mais recuado e um occipital mais saliente (Fig. 3b ). Como esperado, os vLCAs apresentam a maioria das características morfológicas que seriam consideradas específicas de H. sapiens . Na calvária, o neurocrânio é arredondado e globular; o osso frontal apresenta um tubérculo frontal bem desenvolvido 42 e arcos superciliares descontínuos; os parietais mostram eminências parietais distintas; e o basicrânio é marcadamente flexionado. A face está retraída, mostrando um zigomático angulado e voltado para frente 43 , juntamente com uma fossa canina superior desenvolvida com duas curvas fortemente marcadas ( incurvatio horizontalis e sagitallis 44 ). No entanto, eles também compartilham características com fenótipos mais arcaicos: na norma lateral , o frontal é ligeiramente recuado, as sobrancelhas são projetadas para frente e separadas da escama frontal por um leve sulco, a face mostra prognatismo alveolar, o processo zigomático é alinhado com o crista supramastóidea 42 , os processos mastóideos são pouco desenvolvidos, e o occipital alongado mostra um grau de protrusão lambdóide, que juntamente com uma leve depressão no obelion, aparece como um bolo occipital incipiente. As duas últimas características lembram a morfologia Neandertal 12 . Na norma frontalis , a borda anteroposterior da maxila ( incurvatio inframalaris frontalis ) é pouco marcada, como observado em H. heidelbergensis sl 44 , e a distância interorbital é particularmente ampla 45 . Assim, os vLCAs capturam os aspectos exclusivamente derivados de uma morfologia humana moderna e a retenção atualmente geograficamente dispersa de caracteres plesiomórficos entre diferentes populações humanas. As hipóteses 1b e 2b, calculadas na ausência dos Khoisans e Pigmeus, geram formas ancestrais que diferem apenas ligeiramente daquelas baseadas nas hipóteses originais (Suplementar Fig. 3a e b ). O desvio máximo da superfície entre vLCA1 e 1b e vLCA2 e 2b é, no entanto, maior (respectivamente, <1,00 mm e <2,96 mm, tabela suplementar 6 ). O vLCA1b mostra sobrancelhas ligeiramente mais salientes, mas uma face menos prognática, enquanto o 2b é claramente menos robusto do que o vLCA2. Quando comparados entre si a diferença é maior (<3,13 mm, Tabela Complementar 6 ) e vLCA2b apresenta uma morfologia mais graciosa (Fig. 3c Complementar ).
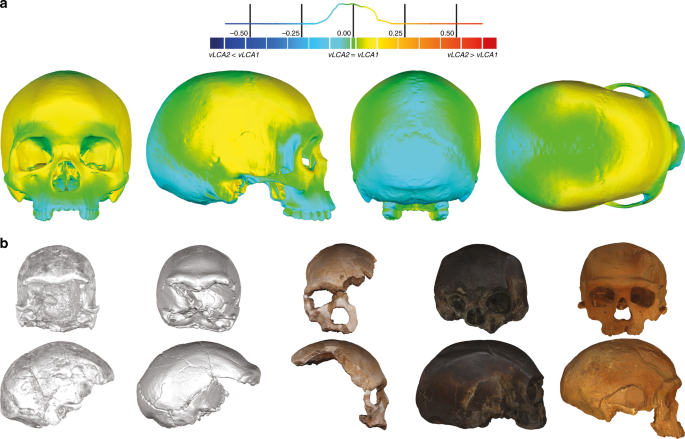
Morfologia dos vLCAs e dos fósseis LMP. a Norma frontalis , lateralis , verticalis , occipitalis de vLCA1 quando comparado com vLCA2 através de uma análise de desvio de superfície. O histograma indica a distribuição do desvio em mm entre cada vértice dos modelos 3d. b Da esquerda para a direita, norma frontalis e lateralis de Omo II, LH18, Florisbad, KNM-ES 11693 (a visualização da norma lateralis é espelhada) e Irhoud 1. Os dados de origem são fornecidos como um arquivo de dados de origem
LMP Diversidade de hominídeos africanos
Figures 4 and 5 show the first two bgPCs of four bgPCAs based on the Procrustes residuals of the individual specimens (to the exclusion of early Homo) grouped according to the terminal taxa of the phylogenetic hypotheses, along with the four computed vLCAs and the five LMP fossils. The four bgPCAs were run on different landmark configurations composed of type I homologous landmarks5 linked by curves of semilandmarks. The four landmark configurations reflect the preserved data of the LMP fossils (see Methods, Supplementary Table 7 and Supplementary Fig. 6). Figure 4a shows the first two bgPCs (52.9% of variance, Supplementary Table 8) of the bgPCA run on the full skull (Analysis A). BgPC1 discriminates the Neandertals (negative values) from modern humans (positive values), while bgPC2 explains the morphological variation within the modern human sample. The position of the modern human groups reflects their evolutionary history: sub-Saharan Africans (#1 to 9) present negative values, as do the early H. sapiens from Qafzeh and Skhūl; Oceanians (#10 and 11), North Africans (#12) and South Asians (#15) cluster around the centre of bgPC2, while all other Eurasians have positive values. The vLCAs cluster around the early H. sapiens, but vLCA2b is closer to Oceanians (#10 and 11). The LMP fossils, Florisbad, KNM-ES 11693 and to a lesser extent Omo II are similar to both early H. sapiens and the vLCAs, while Irhoud 1 shares more similarities with Neandertals. LH18 presents extreme negative values on bgPC2 due to a receding frontal profile and a short medio-lateral length. The Euclidean distances between group means (Supplementary Data 1) confirm these patterns: vLCAs 1, 2 and 1b are closer to early H. sapiens (respectively, 0.0254, 0.0255 and 0.0230), while vLCA2b is closer to Australians (0.0240). Florisbad is the African LMP specimen which resembles the vLCAs the most, particularly vLCA1 and 2 (0.0419), while Irhoud 1 is the only LMP specimen sharing the most similarities with Neandertals (South Europe Neandertals: 0.0617). Figure 4b mostra os dois primeiros bgPCs (60,7%, Tabela Suplementar 9 ) do bgPCA executado apenas na calvária e excluindo Florisbad (Análise B, Tabela Suplementar 7 e Fig. Suplementar 6 ). O morfoespaço é semelhante à análise A. VLCAs 1, 2 e 1b são semelhantes aos primeiros H. sapiens , enquanto vLCA2b é mais próximo das populações atuais. Omo II e KNM-ES 11693 mostram afinidades fenotípicas com os primeiros H. sapiens e os primeiros neandertais, enquanto a morfologia de Irhoud 1 compartilha semelhanças com os neandertais. VLCAs 1, 2 e 1b apresentam a menor distância euclidiana (Dados Suplementares 2 ) para os primeiros H. sapiens (respectivamente, 0,0233, 0,0235 e 0,0214), enquanto vLCA2b está mais próximo dos africanos orientais (0,0202). Dentro dos fósseis LMP, Omo II compartilha mais semelhanças com os vLCAs (0,0672 com vLCA1b), mas está mais próximo do H. sapiens primitivo (0,0665). A distância euclidiana mais curta de Irhoud 1 é para os neandertais do sul da Europa (0,0395).
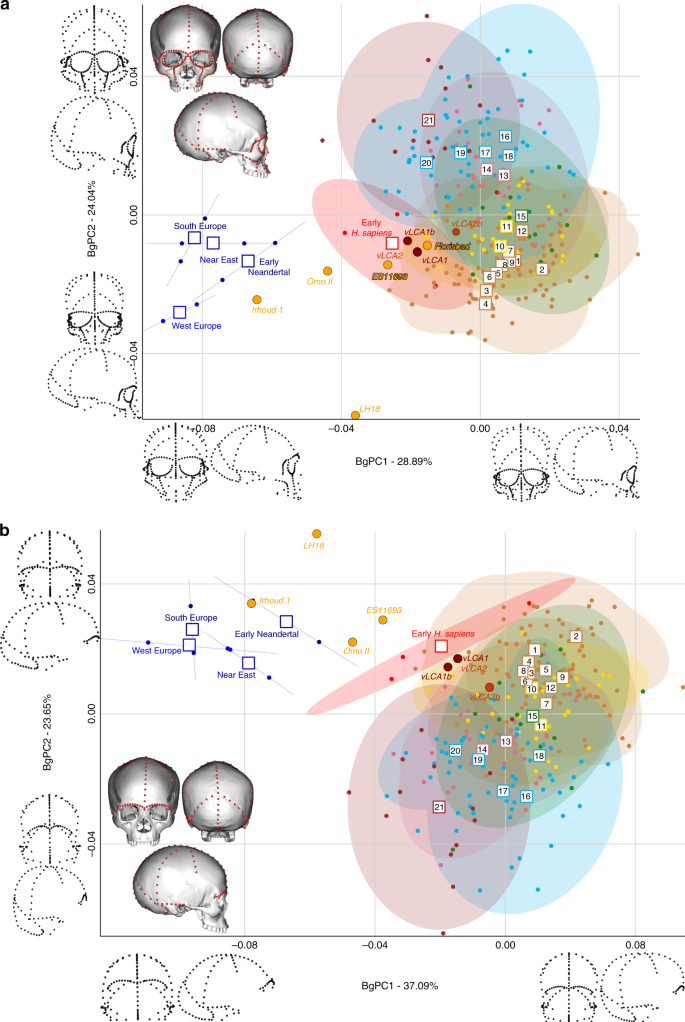
Morfoespaços dos bgPCAs para as análises A ( a) e B ( b) . As elipses representam o intervalo de confiança de 90% para a distribuição estimada dos espécimes de cada população. Os vLCAs têm uma forma mais próxima dos primeiros H. sapiens , bem como dos espécimes LMP africanos Flosibad, KNM-ES 11693 e Omo II, enquanto Irhoud 1 é mais semelhante aos neandertais. Populações humanas modernas como segue: 1 a 9 África Subsaariana; 10–11 Oceania, 12 Norte da África, 13–14 Europa, 15 Sul da Ásia, 16 a 20 Leste Asiático, 21 América do Norte (ver Fig. 1 , Fig. 1 complementar e Tabela 1 complementar ). Os dados de origem são fornecidos como um arquivo de dados de origem
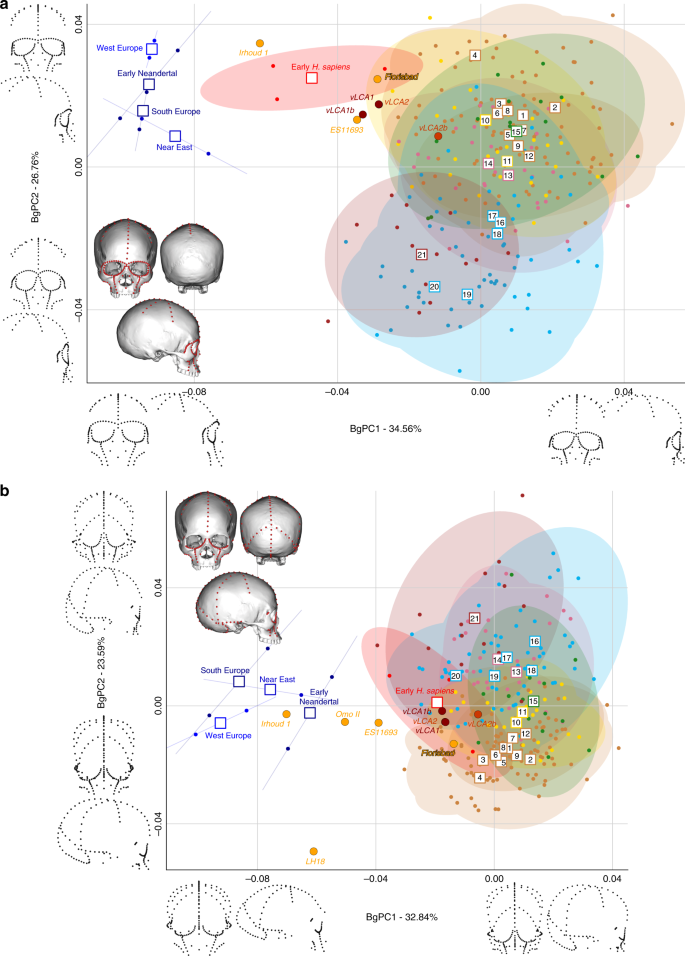
Morphospaces of the bgPCAs for analyses C (a) and D (b). The ellipses represent the 90% confidence interval for the estimated distribution of the specimens of each population. The vLCAs are closer in shape to the early H. sapiens, as well as the African LMP specimens Flosibad, KNM-ES 11693 and Omo II, while Irhoud 1 is more similar to Neandertals. Modern human populations as follow: 1 to 9 Sub-Saharan Africa; 10–11 Oceania, 12 North Africa, 13–14 Europe, 15 South Asia, 16 to 20 East Asia, 21 North America (see Fig. 1, Supplementary Fig. 1 and Supplementary Table 1). Source data are provided as a Source Data file
A Figura 5 exibe os dois últimos bgPCAs executados na configuração de referência preservada de Florisbad (Análise C, Fig. 5a ) e KNM-ES 11693 (Análise D, Fig. 5b ). Omo II e LH18 não estão incluídos na análise C. Ambos os morfoespaços preservam a mesma distribuição global de pontos. No entanto, na análise do morfoespaço de C (61,3%, Tabela Suplementar 10 ), os primeiros H. sapiens estão posicionados entre os humanos modernos e os Neandertais. VLCAs 1, 1b e 2 estão situados entre o H. sapiens moderno e o fóssil atual e compartilham fortes semelhanças com Florisbad e KNM-ES 11693. Ao contrário das análises A e B, Irhoud 1 está mais próximo do H. sapiens primitivo e é, em geral, menos semelhante aos Neandertais (Fig. 5a ). Isso é esperado, pois a análise C foca principalmente na morfologia facial que é mais moderna em Irhoud 1 do que em sua abóbada 18 . As distâncias Euclidianas (Dados Suplementares 3 ) sublinham este padrão de afinidades: vLCAs 1, 2 e 1b estão mais próximos dos primeiros H. sapiens , enquanto vLCA2b está mais próximo dos australianos. Florisbad é o mais semelhante aos vLCAs (0,0502 com vLCA2) e Irhoud 1 é o mais próximo do início H. sapiens (0.0480). Finally, Fig. 5b presents the morphospace (56.4%, Supplementary Table 11) of analysis D. The exclusion of the missing landmarks of KNM-ES 11693 modifies slightly the relative phenotypic affinities, with early H. sapiens being more closely grouped with current modern human populations. The vLCAs cluster close to the centre of early H. sapiens variation, with the exception of vLCA2b (Fig. 5b). Florisbad and KNM-ES 11693 are the closest to both the H. sapiens fossils and the vLCAs. Omo II occupies a central position between modern and Neandertal populations, while Irhoud 1 presents strong affinities to Neandertals. Finally, the associated Euclidean distances (Supplementary Data 4) underline the strong similarities between vLCAs 1, 2 and 1b and early modern humans (respectively, 0.0211, 0.0212 and 0.0187), while vLCA2b is closer to Papuans (0.0214). Florisbad is the closest African LMP specimen to the vLCAs (0.0364), followed by Omo II (0.0605) and KNM-ES 11693 (0.0678). Irhoud 1, as in analysis A and B, is closest to the South European Neandertals (0.0476).
Os boxplots baseados em distâncias de Procrustes (Figs. 4 e 5 complementares ) apresentam as semelhanças dos vLCAs com os espécimes africanos LMP, Neandertal e H. sapiens para as análises AB, C e D.
Na análise A (Fig. complementar 4a e b ), os vLCAs estão mais próximos dos grupos humanos modernos, especialmente do H. sapiens primitivo , com o qual os vLCAs 1, 2 e 1b compartilham a distância mediana mais curta. Esses três vLCAs estão ainda mais próximos de Florisbad, com distâncias de Procrustes mais curtas do que a distância média de Procrustes para qualquer outro grupo. As distâncias de LH18, KNM-ES 11693, Irhoud 1 e Omo II Procrustes para os vLCAs são maiores. O vLCA2b compartilha mais semelhanças com os humanos modernos atuais. O padrão geral de similaridade com os vLCAs é comparável na análise B (Fig. 4c ed Complementar ). No entanto, na ausência de Florisbad, nenhum dos fósseis LMP africanos compartilha fortes semelhanças com os vLCAs: a distância mais curta é para Omo II, mas é maior do que qualquer uma das distâncias médias para os humanos modernos. A análise C (Fig. 5a eb suplementar ) é comparável à análise A: vLCAs 1, 2 e 1b apresentam uma distância semelhante a Florisbad do que à mediana do H. sapiens primitivo , e vLCA2b está mais próximo dos humanos modernos recentes. KNM-ES 11693 e Irhoud 1 apresentam maiores distâncias de Procrustes para vLCAs 1, 2 e 1b do que suas distâncias medianas para humanos modernos. Finalmente, a análise D (Fig. 5c e d ) retratam uma imagem diferente, pois os vLCAs estão mais próximos dos humanos modernos do que dos fósseis LMP africanos (KNM-ES 11693, LH18, Omo II e Irhoud 1), com exceção de Florisbad.
Discussion
The methodological approaches used in the present study have inherent uncertainties—biases from using different means and/or operators46,47,48 to obtain 3D data, number of variables versus the number of specimens, sliding of semilandmarks49,50,51, and sampling error to obtain mean population shapes52. To minimise this, the data were collected by a single operator; we reduced the number of variables by using PCs53 instead of aligned 3D coordinates throughout the study, and test models were run on non-slid semilandmarks, as well as subsamples of landmarks and semilandmarks (Supplementary Figs. 7–10). Supplementary Fig. 8 shows that the number of landmarks and the use of slid semilandmarks50 do not impact the reliability of the results. While sample size remains limited to the available fossil sample, sampling error can be assessed in the bgPCA analyses where specimens were analysed individually (Figs. 4 and 5, Supplementary Data 1–4). Finally, the modelling approach used here computes ancestral shapes which are weighted averages derived from the terminal taxa of a phylogenetic hypothesis54 of given topology using a Brownian motion model for evolution that mostly estimates drift. Those ancestral estimations are not intended to accurately represent evolution, but to act as tools to study the African LMP hominin fossil record in order to bring new insights into the origin of H. sapiens.
Our findings show that, first, the phenotypic data presented in this study relate closely to the genetic history39,55 of the considered populations. This is illustrated by the phylo-morphospace based on mean population shape data (Fig. 2), but also by the distribution of individual specimens’ shapes (Figs. 4 and 5). In addition, hypothesis 1, which takes into account an early out-of-Africa which preserves a phenotypic signal among the people of Sahul39, and hypothesis 2, which follows a more classic out-of-Africa event55, led to the computation of ancestral morphologies which are virtually undistinguishable (Fig. 3a, Supplementary Table 6). Yet, the topology of the phylogenetic tree of hypothesis 1 fits more closely the phenotypic variation of the data, as exemplified by the phylogenetic signal computed for the different phylogenies (Supplementary Table 4). The exclusion of the Khoisans and Pygmies has little impact on the first hypothesis, as the phenotype of vLCA1 and 1b remains comparable (Supplementary Fig. 3). Hypothesis 2 is more affected by this change, resulting in a very gracile morphotype (Supplementary Fig. 3) and lower values for the phylogenetic signal metrics (Supplementary Table 4). Therefore, the scenario simulated in hypothesis 1 appears to be slightly better supported by the phenotypic data presented here, which is congruent with recent genomic results14,39.
Em segundo lugar, as morfologias dos vLCAs são graciosas e modernas. As características cranianas derivadas do H. sapiens são totalmente exibidas nos vLCAs - um neurocrânio abobadado, uma face reduzida e uma acentuada flexão basicraniana 17 , e apenas parcialmente equilibrada por características mais arcaicas (isto é, sobrancelhas salientes, prognatismo alveolar marcado, fracamente processos mastóides desenvolvidos, osso occipital alongado, incurvatio inframalaris frontalis fracamente marcado e ampla distância interorbital; ver refs. 12 , 42 , 44 , 45 ). Uma explicação para esta morfologia grácil pode ser encontrada na estrutura das hipóteses filogenéticas. Sob um modelo de evolução de Movimento Browniano, o comprimento do ramo contribui significativamente para a reconstrução ancestral nos nós 54 . Táxons terminais com ramos longos contribuirão mais para a estimativa ancestral. Nas presentes hipóteses, a influência mais forte na reconstrução ancestral deve ser dos Khoisans e dos Pigmeus e, em menor grau, dos primeiros H. sapiens . Os Khoisans e Pigmeus se ramificam muito cedo a partir do H. sapiens stem lineage13,40, which may give an ‘archaic’ weight to their particularly gracile morphologies41. The derived morphologies of these populations, partly due to adaptation to the environment56, should be different from their ancestors’ phenotypes. The shape of the vLCAs may hence be influenced by this Khoisan/Pygmy morphological pattern by overestimating the antiquity of this gracile phenotype. This effect may only be partly balanced by the comparatively large and robust individuals (Qafzeh 6 and 9, and Skhūl V) of the early H. sapiens terminal taxon, since they are chronologically older than extant human populations and have a consequently shorter branch length (i.e., ~100 ka19,57). However, the early H. sapiens taxon plots within the 95% confidence envelope of the vLCAs in the phylo-morphospaces (Fig. 2 and Supplementary Fig. 2). In addition, the divergent morphologies (Supplementary Figs. 2 and 3) estimated from the b hypotheses reinforce this idea: the least plausible phylogeny (hypothesis 2b) yields the least plausible ancestral estimation for modern humans (vLCA2b). Thus, the gracile morphology of the computed vLCAs, while influenced by the theoretical framework of the modelling approach used here, appears to be more than just an artefact of the derivation of the ancestral morphologies from the extant descendants.
Apesar das incertezas acima mencionadas, e dadas as afinidades fenotípicas apresentadas pelos hominídeos africanos LMP, uma interpretação paleoantropológica desses resultados é possível. A rápida fixação dos traços cranianos derivados compartilhados por todos os H. sapiens recentes com expressão variável de traços de robustez 58 poderia explicar os fenótipos dos vLCAs. Isso pode ter ocorrido por um processo de deriva localizada durante o final do Pleistoceno Médio, levando à estruturação populacional 16 . As morfologias do mosaico fóssil africano LMP combinam caracteres arcaicos e modernos, e a primeira ocorrência de um morfotipo moderno completo não está documentada antes de Omo I (195 ka) e espécime BOU-VP16/1 de Herto (160 ka). Nenhum dos outros fósseis LMP mostra a expressão combinada de traços derivados de humanos modernos, enquanto os vLCAs computados são humanos modernos e os caracteres cranianos universais compartilhados por todos os membros da espécie estão presentes. Isso poderia apoiar um rápido aparecimento de H. sapiens , consistente com a teoria do equilíbrio pontuado . 59 , porém não há evidências de um longo e estável período de estase no registro fóssil do Pleistoceno Médio. Pelo contrário, as evidências atuais sugerem um período biogeográfico dinâmico impulsionado por mudanças pronunciadas na aridez e na temperatura 60 , durante as quais a deriva e a seleção teriam agido para criar diversidade nos níveis de população e potencialmente de espécies.
This scenario is consistent with the high level of phenotypic diversity in the LMP African hominin record. The results presented here highlight this diversity and allow to hypothesise the existence of different LMP morphotypes in Africa reflecting a structured population history (Fig. 4b). Three broad morphological patterns may be identified in the LMP fossil record: one represented by the Eastern African fossil LH18, the African LMP fossil sharing the least affinities with modern humans and the computed vLCAs (although some distortions61 on the calvarium may partly explain these phenotypic affinities); another exemplified by the North African fossil Irhoud 1, whose morphology is intermediate between modern humans and the Neandertals; and a possible third morph, represented by the South African fossil of Florisbad, and, to a lesser extent by the East African fossils KNM-ES 11693 and Omo II. Those three specimens present the shortest Euclidean distances to modern humans in all four analyses when the vLCAs are not taken into consideration (Supplementary Data 1–4). Indeed, the second shortest Euclidean distances observed is between Florisbad and early H. sapiens, between KNM-ES 11963 and the San, and between Omo II and early H. sapiens, and for analysis D with Alaskan Inuit (Supplementary Data 1–4). On the other hand, shortest Euclidean distance between Irhoud 1 and other groups is always to the South European Neandertals, except for analysis B where it is with early H. sapiens. Analysis B focuses mostly on the face which is where modern human-like traits have been identified in Irhoud 118. Finally, Florisbad is the LMP fossil most similar to the vLCAs, as exemplified by the Euclidean (Supplementary Data 1, 3, and 4) and Procrustes distances (Supplementary Figs. 4 and 5), and by the presence of a well-defined canine fossa, a fully formed tuber frontale and weakly projecting brow-ridges (Fig. 3b).
Esses resultados ressoam com a idade dos do H. naledi restos mortais sul-africano 4 , um hominídeo de cérebro pequeno com um mosaico incomum de traços antigos e modernos, datado do início do LMP (ou seja, 335–236 ka 32 ). de aparência 'arcaica' do LMP Tanto a descoberta de uma espécie de Homo quanto os resultados do presente estudo sublinham a complexidade da variação morfológica dentro do gênero Homo durante o LMP africano. Além de um alto grau de estrutura populacional dentro da linhagem evoluindo sapiens , conforme sugerido por nossas análises, essa complexidade pode abranger a presença de diferentes linhagens Homo na África. Enquanto algumas populações descendentes de uma linhagem africana de H. heidelbergensis podem ter coalescido para formar H. sapiens seguindo a hipótese do multirregionalismo africano 9 , é provável que alguns fósseis africanos LMP não tenham sido associados a nenhuma população ancestral de H. sapiens . Devemos, portanto, esperar encontrar fósseis LMP na África que eram membros de ramos laterais sobrepostos cronológica e geograficamente da árvore da evolução humana do Pleistoceno, como o Os restos de H. naledi parecem ser 4 , refletindo diferentes profundidades de tempo de formação de linhagem e extensão de diferenças adaptativas. A eventual perda desses ramos laterais pode ser o resultado do efeito cumulativo da resiliência diferencial tanto às mudanças climáticas quanto à competição entre grupos. Embora o registro fóssil e arqueológico do LMP seja atualmente insuficiente para construir um modelo coerente de mudança diacrônica nas distribuições populacionais, ele poderia sugerir um processo multifásico 16 para a evolução dos humanos modernos e sua diversidade. Após a origem da espécie no final do Pleistoceno Médio, uma segunda fase caracterizada pela expansão bem-sucedida das populações humanas modernas 16 pode ter começado em 194 ka 3 , mas é claramente estabelecida em 130 ka, conforme documentado pelo aumento no número de sítios arqueológicos 62 , e a presença de uma população humana moderna no Levante 3 , 43 . Os fósseis de H. sapiens do leste da Ásia 63 , cujo limite cronológico inferior pode se estender até 120 ka 64 , podem ter feito parte dessa fase de expansão anterior. A análise genômica oferece evidências conflitantes sobre uma expansão tão precoce dos humanos modernos na Eurásia. A introgressão relatada do DNA humano moderno em neandertais que viveram ca . 100.000 anos atrás nas montanhas de Altai 65 sugeririam que as populações africanas realmente se expandiram além do Levante durante o último interglacial, assim como alguns dos resultados baseados na variação genotípica de australianos e papuas hoje 14 , 39 , 55 . A contribuição desta dispersão para fora da África durante o Marine Isotope Stage (MIS) 5 para a origem da atual diversidade humana na Eurásia permanece incerta. No entanto, outras populações e espécies de hominídeos (por exemplo, naledi ) na África parecem ter desaparecido em grande parte no MIS5, possivelmente devido às duras condições climáticas durante o MIS6 66 , que podem ter desencadeado eventos de extinção local e à competição com grupos em expansão de H. sapiens durante as fases iniciais do MIS5.
Além da complexidade das interações potenciais entre os hominídeos africanos LMP, o que o presente estudo pode nos dizer sobre a origem geográfica do primeiro H. sapiens ? Dados genômicos atuais apontam para uma origem do sul 67 ou da África oriental 68 , ou através de um cline leste-sul da África; 69 paleoantropologia sugere uma origem do norte 18 , 70 ou do leste africano 22 , 24 , 62 . Nossos resultados tendem a apoiar um padrão evolutivo complexo que pode ter envolvido diferentes populações de origem, possivelmente incluindo grupos do sul e do leste africano, como foi recentemente defendido pela genômica 69 . Tanto o fóssil Florisbad da África Austral quanto os espécimes da África Oriental KNM-ES 11693 e Omo II mostram semelhanças com os vLCAs e os primeiros H. sapiens . Uma origem do norte da África é menos provável 18 , pois Irhoud 1 apresenta um padrão de afinidade diferente, tornando-o morfologicamente mais próximo dos neandertais. Os espécimes recém-descritos de Jebel Irhoud trouxeram novas informações sobre a morfologia dos hominídeos LMP do norte da África. A mandíbula Irhoud 11 mostra algumas características derivadas (ou seja, aumento anterior na altura do corpo mandibular e seção estreita), mas carece de um queixo T invertido humano moderno completo 18 como foi observado antes no espécime juvenil de Irhoud 3 71 . A face superior fragmentária (Irhoud 10), é morfologicamente semelhante à face superior de Irhoud 1 18 , que, neste último espécime, se enquadra, mas no limite da distribuição das morfologias faciais humanas modernas 36 , ou, quando analisada como uma parte integrada do crânio no presente estudo, compartilha mais semelhanças com um possível humano moderno e Neandertal LCA do que com os computados H. sapiens vLCAs. Uma hipótese alternativa sobre a população norte-africana representada pelos fósseis de Jebel Irhoud poderia se basear em resultados genômicos recentes sugerindo que o DNA mitocondrial Neandertal ancestral (mtDNA), compartilhado com os fósseis Denisova 72 e Sima de los Huesos 73 , foi substituído por um africano mtDNA entre 460 e 219 ka 15 . Considerando as afinidades fenotípicas de Irhoud 1 com os neandertais e os primeiros H. sapiens , é possível que os fósseis de Irhoud representem descendentes locais de uma população africana que se dispersou para fora da África durante um evento do Saara Verde associado ao MIS9, portanto relacionado às populações africanas que se introgrediu na linhagem européia nesta época e, portanto, contribuiu para a evolução dos neandertais 'clássicos'.
Following from these results, it is possible to tentatively draw a framework for the origin of H. sapiens. The speciation process appears to have been complex, going through different phases16 that may not have contributed to the genetic and phenotypic structure of current modern human populations. A first stage of phenotypic diversification, from 350 to 200 ka, may have happened locally with different contemporary populations forming local morphs of pre-H. sapiens groups as they are represented in the LMP fossil record. This phase may have been followed by a period of fragmentation and differential expansion of populations leading to hybridisation and coalescence of groups, which could have resulted in the emergence of morphologically derived populations of anatomically modern humans between 200 to 100 ka, as exemplified by the fossils from Herto, Skhūl and Qafzeh. Nevertheless, our results suggest that it is unlikely that all LMP local populations would have contributed equally, or at all, to the lineage that gave rise to the population ancestral to H. sapiens; local extinctions and founder effects would have shaped considerably the emergence of anatomically modern humans16. The morphology of the vLCAs computed in the present study appears to be closer to this last phase (i.e., 200–100 ka) than to the former one. This may indicate that chronologically older fossils of anatomically modern H. sapiens, representing populations which outlived most of the LMP hominin groups, are yet to be found.
Methods
Materials
We used a sample of 263 3D crania (see below and Table 1, Supplementary Table 1, Supplementary Fig. 1) to produce mean populations shapes which serve as terminal taxa of the phylogeny of the genus Homo (Fig. 1a, b): seven early Pleistocene specimens representing the species H. habilis, H. ergaster, and H. georgicus; eight Neandertals from amongst different geographical populations (early, Near-East, South-Europe and West-Europe Neandertals), and three early H. sapiens fossils (Qafzeh 6 and 9, Skhūl V). 245 extant modern humans represent current H. sapiens groups and are organised through 21 populations (i.e., ten Africans, eight Eurasians, two Inuits and a North American group, see Supplementary Table 1 and Supplementary Fig. 1). The five African LMP targeted fossils (Table 1, Fig. 3) are from North Africa (Irhoud 1), from East Africa (KNM-ES 11693, LH18 and Omo II), and from South Africa (Florisbad), and they were not included in the estimation of the H. sapiens vLCAs. These specimens are among the few fossils that could represent the ancestral population to all modern humans: besides originating in Africa, their estimated dates fit within the estimated timeframe put forward by palaeoanthropological and genomic studies for the origin of H. sapiens9,69, as well as with the most recent date estimates based on the study of Khoisan ancient genomes suggesting that the last genetic common ancestral population of all living humans may have lived more than 260,000 years ago69. We did not include the fossils of Guomde25 and Omo I22 due to their preservation state, or Singa due to its pathological condition35. We did not have access to the BOU-VP16/1 specimen.
3D models building
Os modelos 3D foram obtidos seguindo três procedimentos (Tabela 1 ): (1) tomografias computadorizadas médicas (tamanho do voxel entre 0,449219 e 0,488281 mm) processadas no Amira (v5.5, FEI); (2) fotogrametria utilizando o software Photoscan (Agisoft, v1.2.6); e (3) varreduras de superfície 3D usando um scanner óptico (HDI Advance, precisão de 45 µ, LMI) e o software FlexScan 3D (v.3.3, LMI). A precisão da fotogrametria é suficientemente boa para produzir modelos 3D confiáveis com uma margem de erro semelhante à da varredura de superfície a laser 47 . A comparação de modelos 3D obtidos de tomografias computadorizadas médicas e varredura de superfície a laser também mostram que ambos os métodos produzem modelos de qualidade comparável 48 . Os modelos resultantes (em média 1,5 milhão de vértices), tanto da filogenia quanto das amostras LMP, são descritos por 780 marcos (software Landmark, IDAV 74 ), entre os quais 724 são semimarcos (116 em curvas localizadas na face e 608 em superfícies localizadas na a calvária) que podem deslizar 5 , 75 (Fig. 1b ). Para estimar o impacto de semimarcos deslizantes na presente análise, um teste foi executado na hipótese 1 comparando resultados de coordenadas de semimarcos não deslizantes com os resultados apresentados neste manuscrito. As diferenças entre ambas as análises são pequenas (Análise E, Figs Suplementares. 7 e 8 , Tabelas Suplementares 12 – 14 ). Grande número de variáveis pode ter efeitos em algumas análises 49 . Para verificar esse impacto na presente análise, dois testes foram executados na hipótese 1, e dois vLCAs (vLCA1Sub e vLCALd) foram calculados com base em uma subamostra de 239 semimarcos e apenas em 53 marcos. A diferença na morfologia dos vLCAs é pequena (Análise F, Figuras Suplementares. 8 – 10 , Tabelas Suplementares 14 – 17 ). O deslizamento de semimarcos minimiza as diferenças de forma entre cada espécime e a forma média na amostra (ou seja, variação devido ao espaçamento arbitrário das coordenadas dos pontos) 50 . A abordagem modifica ligeiramente as coordenadas dos dados brutos e tem sido debatida nos últimos 50 , 51 . Pontos de referência ausentes são estimados espelhando os pontos de referência existentes no outro lado e, quando isso não é possível, os poucos pontos de referência ausentes restantes foram estimados por interpolação de placa fina spline (isto é, TPS 76 ). Florisbad, Omo II e LH18 necessitaram de grandes reconstruções para serem alinhadas (GPA) com os vLCAs e a amostra filogenética. No entanto, a análise e comparação de sua morfologia é baseada na configuração anatômica preservada de cada espécime. Para corrigir a assimetria bilateral, usamos o componente simétrico da Análise Procrustes Generalizada (ou seja, GPA 77) in all subsequent analyses. The data collection, photogrammetry, surface scanning, and segmentation for 3D model building, along with collection of landmarks, were done by one of us (AM).
Phylogenetic modelling
Para calcular a morfologia dos vLCAs, usamos duas hipóteses que representam filogenias totalmente resolvidas do gênero Homo (Fig. 1a, b ). Ambas as hipóteses são baseadas na mesma topologia geral: três espécies ( H. habilis : KNM-ER 1813 e 1470, H. ergaster : KNM-ER 3733, KNM-ER 3883 e KNM-WT 15000, e H. georgicus : D2282 e D2700) compõem o grupo externo dos dois táxons irmãos, Neandertais (Early (Próximo ao Oriente (Sul da Europa, Europa Ocidental))) e humanos modernos. O clado Neandertal é construído para refletir possíveis subgrupos na população: 78 Neandertais ocidentais 'clássicos', mais derivados do que os indivíduos do sul, do Oriente Próximo e do início do Neandertal. O clado humano moderno é uma simplificação de nossa história evolutiva: 11 os espécimes Qafzeh e Skhūl são um grupo irmão do existente H. sapiens 43 , que são organizados seguindo suas relações genéticas relativas (ver Tabela Suplementar 2 e refs. 14 , 39 , 55 ). Os táxons terminais são posicionados em relação à sua cronologia; os nós da filogenia refletem cronologias consensuais baseadas em dados genéticos e paleoantropológicos 8,36,55, and the position of the early Homo outgroup is based on extensive palaeoanthropological studies2. The common ancestry between modern humans and Neandertals is positioned at 600 ka, following suggestions from latest genomic estimates8 and from previous phylogenetic modelling results;36 the Neandertal clade is rooted at around 300 ka and the modern human clade at 305 ka69 (Fig. 1 and Supplementary Table 2). Both hypotheses place the African populations at the base of the clade. Hypothesis 1 takes into account an extinct early out-of-Africa which can be detected in current Oceanian populations’ genomes39. This migration event, which was followed by a later out-of-Africa, left a significant but small signature in the current Sahul populations along with other signatures from admixture with Neandertal and/or Denisovan populations39. In order to simulate this complex scenario in hypothesis 1, the Australian and Papuan branches split from the African clade at 90 ka corresponding to the median genetic split time between West Africans and Papuans calculated by Pagani and colleagues39 (Fig. 1a). Hypothesis 2 ignores this possible extinct out-of-Africa, and follows a more traditional topology55, with non-African populations splitting from the Africans at 75 ka (Fig. 1b). Two alternative hypotheses (i.e., hypotheses 1b and 2b) removing the Khoisan and Pygmy populations (Fig. 1) from the phylogenies were also tested. The continuous variables used to describe the terminal taxa are principal components (i.e., PCs) representing the mean shape variables for each population obtained after performing a GPA and a Principal Component Analysis on the specimens’ landmark sets. The ancestral shapes are computed for each of the mathematically uncorrelated PCs using maximum likelihood79 within a model of evolutionary change assuming random walks in continuous time (i.e., Brownian motion model80) which mostly approximates genetic drift but can also reflect adaptive evolutionary changes;81 however, genetic drift seems to explain most of the cranial morphological divergence observed between some hominin populations82. The use of PCs to perform cladistics analyses83 has been criticised84, but PCs are considered more suitable for maximum likelihood based analyses85. Maximum likelihood reconstructs the ancestral states to maximise the probability of the states observed among the known taxa of a given hypothetical phylogeny (i.e., topology, branch length) following a statistical evolutionary divergence model79 (i.e., Brownian motion model80). The maximum likelihood approach computes for each node of the phylogeny the most likely ancestral shape along with a 95% confidence envelope (Fig. 2). The confidence envelope is a quantification of the uncertainty based on a standard probabilistic measure that depends on the rate of variance of the characters, the length of the branches, and the topology of the phylogeny. In the case of a deep ancestry between two sister taxa, the uncertainty is larger, a more recent common ancestry will present less uncertainty54. The ancestral PCs are rotated back into the landmark space86 giving a set of coordinates in which a unique modern human skull (specimen Kh-1739; see Supplementary Table 1) is warped (TPS warping76) to produce the different fully rendered 3D vLCAs (see ref. 36 and Fig. 3). Finally, to assess the correlation between each phylogenies and the data imputed, we computed their phylogenetic signal for the PCs and the aligned 3D coordinates using a multivariate K statistic37,38.
Desvio do espectro de superfície
Para avaliar as diferenças morfológicas entre os vLCAs obtidos através dos 4 modelos aqui testados (ou seja, hipóteses 1, 2, 1b e 2b), usamos um método de desvio de espectro de superfície que quantifica o desvio de superfície entre uma referência e um corpo de prova. Os marcos ancestrais são alinhados (GPA) e um modelo 3D de um humano moderno (Kh-1739) é distorcido nas configurações. A diferença de forma pode então ser explorada através de um espectro codificado por cores, que dá uma cor a cada um dos vértices do modelo 3D de acordo com o desvio, medido em milímetros, da referência. O desvio pode ser visualizado como histogramas, mostrando a distribuição dos vértices do corpo de prova de acordo com sua distância aos vértices do corpo de referência (Fig. 3 complementar e Fig. 3 ). A Figura 3a apresenta o desvio de vLCA2 (teste) contra vLCA1 (referência). A Fig. 3 complementar apresenta o desvio de superfície de vLCA1b vs 1 (A), vLCA2b vs 2 (B) e vLCA2b vs 1b (C). Finalmente, para cada comparação, calculamos as distâncias máximas positivas e negativas, as distâncias médias positivas e negativas e o desvio padrão (Tabela Suplementar 6 ).
Morfometria geométrica 3D
To assess the pattern of morphological variation of the vLCAs and of the LMP fossils, we ran four analyses using sub-samples of the landmark configuration used to compute the phylogenetic models. The first analysis uses 255 landmarks and semilandmarks distributed on the full skull along the cranial sutures (i.e., bregma-stephanion, bregma-lambda, lambda-asterion) along with additional curves joining craniometric points (i.e., bregma-asterion, asterion-porion, asterion-opsithion, opisthion-lambda, frontomalare orbitale-zygoorbitale, zygoorbitale-canin, zygoorbitale-maxillofrontale, frontomalare temporale-jugale, zygotemporale-lateral M1, glabella-frontozygomatic, Analysis A, Supplementary Fig. 6a); the second is based on the preservation state of Omo II and LH18 and uses only 148 landmarks and semilandmarks on the calvarium (Analysis B, Supplementary Fig. 6b); the third is based on the preservation state of Florisbad and uses 112 landmarks and semilandmarks on the calvarium and face (Analysis C, Supplementary Fig. 6c); and the last analysis is based on the preservation state of KNM-ES 11693 and uses 181 landmarks and semilandmarks on the calvarium and face (Analysis D, Supplementary Fig. 6d). Due to their state of preservation, Omo II and LH18 were discarded from analysis C and Florisbad was discarded from analysis B (Supplementary Table 7). For each analysis, the Neandertal and modern human specimens, used to generate the models, were aligned (GPA) individually along with the vLCAs and LMP fossils. The Procrustes distances from the GPA were examined through boxplots (Supplementary Figs. 4 and 5) and the Procrustes residuals were used to compute four between group Principal Components Analyses (i.e., bgPCA, Figs. 4 and 5) grouping the specimens according to their population, to the exception of the vLCAs and LMP fossils which were not given a group. The Euclidean distances between mean groups were considered to clarify the phenetic relationships of the vLCAs with LMP fossils and the hominin groups (Supplementary Data 1 – 4 ).
We used Geomagic Studio v.2013.0.1 to perform the surface deviation spectrum analysis and R for all other analyses (Morpho v2.1;87 geomorph v2.1.2;88 ape v3.2;89 phytools v0.4–3.190, Ade4 v1.7–491, phylocurve v2.0.992).
Reporting summary
Further information on research design is available in the Nature Research Reporting Summary linked to this article.
Data availability
The 3D model of the vLCAs are available in Supplementary Data 5–11 along with the 3D models showing the deviation patterns between the vLCAs (Supplementary Data 12–18). In order to visualise properly Supplementary Data 5–18, please use the open source Meshlab software or use the R software (see Description of Additional Supplementary Files). The 3D geometric morphometric data (i.e., landmarks coordinates) are available in the Source Data File.
References
Stringer, C. B. Modern human origins: progress and prospects. Philos. Trans. R. Soc. Lond. B Biol. Sci. 357, 563–579 (2002).
Wood, B. & Lonergan, N. The homini fossil record: taxa, grades and clades. J. Anat. 212, 354–376 (2008).
Hershkovitz, I. et al. The earliest modern humans outside Africa. Science 359, 456–459 (2018).
Berger, L. R. et al. Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. eLife 4, https://doi.org/10.7554/eLife.09560 (2015).
Bookstein, F. L. Morphometric Tools for Landmark Data: Geometry and Biology (Cambridge University Press, Cambridge, 1991).
Gómez-Robles, A. et al. espécies de hominídeos conhecidas correspondem à morfologia dentária esperada do último ancestral comum dos neandertais e dos humanos modernos. Proc. Natl Acad. ciência EUA 110 , 18196–18201 (2013).
Reich, D. et al. Mistura de Denisova e as primeiras dispersões humanas modernas no sudeste da Ásia e na Oceania. Sou. J. Hum. Genet. 89 , 516–528 (2011).
Meyer, M. et ai. Sequências de DNA nuclear dos hominídeos do Pleistoceno Médio Sima de los Huesos. Natureza 531 , 504–507 (2016).
Bräuer, G. A origem da anatomia moderna: por especiação ou evolução intraespecífica? Evolução Anthr. 17 , 22–37 (2008).
Wolpoff, MH, Thorne, AG, Smith, FH, Frayer, DW & Pope, GG em Origins of Anatomically Modern Humans (eds. Nitecki, MH & Nitecki, DV) 175–199 (Plenum Press, Nova York, 1994).
Lahr, MM & Foley, RA Rumo a uma teoria das origens humanas modernas: geografia, demografia e diversidade na evolução humana recente. Yb. Física Anthr. 41 , 137–176 (1998).
Condemi, S. em Hominidae (ed. Giacobini, G.) 357–362 (Jaca Book, Milão, 1989).
Lachance, J. et ai. História evolutiva e adaptação de sequências de genoma inteiro de alta cobertura de diversos caçadores-coletores africanos. Cela 150 , 457-469 (2012).
Malaspinas, A.-S. e outros Uma história genômica da Austrália aborígine. Natureza 538 , 207–214 (2016).
Posth, C. et ai. O genoma mitocondrial arcaico profundamente divergente fornece um limite de tempo menor para o fluxo de genes africanos nos neandertais. Nat. Comum. 8 , 16046 (2017).
Mirazón Lahr, M. A conformação da diversidade humana: filtros, fronteiras e transições. Philos. Trans. R. Soc. Lond. B Biol. ciência . 371 , https://doi.org/10.1098/rstb.2015.0241 (2016).
Stringer, CB A origem e evolução do Homo sapiens. Philos. Trans. R. Soc. Lond. B Biol. ciência 371 , https://doi.org/10.1098/rstb.2015.0237 (2016).
Hublin, J.-J. et al. New fossils from Jebel Irhoud, Morocco and the pan-African origin of Homo sapiens. Nature 546, 289–292 (2017).
Grün, R. & Stringer, C. B. ESR dating and the evolution of modern humans. Archaeometry 33, 153–199 (1991).
Richter, D. et al. The age of the hominin fossils from Jebel Irhoud, Morocco, and the origins of the Middle Stone Age. Nature 546, 293–296 (2017).
McDermott, F., Stringer, C. B., Grün, R., William, C. T. & Din, V. K. New Late-Pleistocene uranium-thorium and ESR dates for the Singa hominid (Sudan). J. Hum. Evol. 31, 507–516 (1996).
Day, M. H. Early Homo sapiens remains from the Omo River region of south-west Ethiopia: Omo human skeletal remains. Nature 222, 1135–1138 (1969).
McDougall, I., Brown, F. H. & Fleagle, J. G. Sapropels and the age of hominins Omo I and II, Kibish, Ethiopia. J. Hum. Evol. 55, 409–420 (2008).
White, T. D. et al. Pleistocene Homo sapiens from Middle Awash, Ethiopia. Nature 423, 742–747 (2003).
Bräuer, G. in Humanity from African Naissance to Coming Millennia (eds. Tobias, PV, Michael A. Raath, Jacopo Moggi-Cecchi e Gerald A. Doyle) 191–197 (Firenze University Press, Witwatersrand University Press, Rotterdam, 2001 ).
Bräuer, G., Yokoyama, Y., Falguères, C. & Cat, E. Origens humanas modernas datadas. Natureza 386 , 337–338.
Bräuer, G. & Leakey, R. O crânio ES-11693 de Eliye Springs, West Turkana, Quênia. J. Hum. Evolução 15 , 289-312 (1986).
Bräuer, G. et al. Alterações patológicas no arcaico crânio do Homo sapiens de Eliye Springs, no Quênia. Sou. J. Phys. Anthr. 120 , 200–204 (2003).
Manega, PC em Preservação e Uso de Olduvai Gorge, Laetoli, Arte Rupestre e Outros Recursos Paleoantropológicos na Tanzânia (Bellagio, Itália, 1995).
Day, M., Leakey, MD & Magori, C. Um novo crânio fóssil hominídeo (LH 18) de Ngaloba Beds, Laetoli, norte da Tanzânia. Natureza 284 , 55–56.
Grün, R. et al. Direct dating of Florisbad hominid. Nature 382, 500–501 (1996).
Dirks, P. H. G. M. et al. The age of Homo naledi and associated sediments in the Rising Star Cave, South Africa. eLife 6, e2423 (2017).
Ennouchi, E. LesN. éandertaliens du Jebel Irhoud (Maroc). C. R. Acad. Sci. Paris 256, 2459–2460 (1963).
Pearson, O. M. Statistical and biological definitions of “Anatomically Modern” Humans: Suggestions for a Unified Approach to Modern Morphology. Evol. Anthropol. 17, 38–48 (2008).
Spoor, F., Stringer, C. B. & Zonneveld, F. Rare temporal bone pathology of the Singa calvaria from Sudan. Am. J. Phys. Anthropol. 107, 41–50 (1998).
Mounier, A. & Mirazón Lahr, M. Virtual ancestor reconstruction: revealing the ancestor of modern humans and Neandertals. J. Hum. Evol. 91, 57–72 (2016).
Goolsby, E. W. Likelihood-based parameter estimation for high-dimensional phylogenetic comparative models: overcoming the limitations of “distance-based” methods. Syst. Biol. 65, 852–870 (2016).
Adams, D. C. A generalized K statistic for estimating phylogenetic signal from shape and other high-dimensional multivariate data. Syst. Biol. 63, 685–697 (2014).
Pagani, L. et al. Genomic analyses inform on migration events during the peopling of Eurasia. Nature 538, 238 (2016).
Schlebusch, C. M. et al. Genomic variation in seven khoe-san groups reveals adaptation and complex african history. Science 338, 374–379 (2012).
Franklin, D., Cardini, A. & Oxnard, C. E. A geometric morphometric approach to the quantification of population variation in sub-Saharan African crania. Am. J. Hum. Biol. 22, 23–35 (2010).
Mounier, A. Validité du taxon Homo heidelbergensis Schoetensack, 1908 Ph.D. thesis, Université de la Méditerrannée (2009).
Vandermeersch, B. Les Hommes de Qafzeh (Israël) (CNRS, Paris, 1981).
Mounier, A. Definição do táxon Homo heidelbergensis Schoetensack, 1908: análise fenética do maciço facial superior de fósseis do gênero. Pleistoceno Médio Homo BMSAP 23 , 115–151 (2011).
Weidenreich, F. O crânio de Sinanthropus pekinensis : um estudo comparativo de um crânio hominídeo primitivo. Paleontologia. Pecado. 10 , 1–485 (1943).
Fruciano, C. et al. Compartilhar é se importar? Erro de medição e problemas decorrentes da combinação de conjuntos de dados morfométricos 3D. Eco. Evolução 7 , 7034–7046 (2017).
Katz, D. & Friess, M. Nota técnica: 3D a partir de fotografia digital padrão de crânios humanos - uma avaliação preliminar. Sou. J. Phys. Antropol. 154 , 152–158 (2014).
Slizewski, A., Friess, M. & Semal, P. Varredura de superfície de espécimes antropológicos: comparação nominal-real com scanner a laser de baixo custo e sistemas de varredura de superfície de projeção de luz de franja de ponta. Quartar 57 , 179–187 (2010).
Bookstein, FL Uma fórmula recém-observada impõe limites fundamentais em análises morfométricas geométricas. Evolução Biol. 44 , 522–541 (2017).
Gunz, P. & Mitteroecker, P. Semilandmarks: um método para quantificar curvas e superfícies. Hystrix It. J. Mam. 24 , 103–109 (2013).
MacLeod, N. PalaeoMath: part 28-landmarks and semilandmarks. Palaeontol. Newsl. 82, 32–43 (2013).
Cardini, A., Seetah, K. & Barker, G. How many specimens do I need? Sampling error in geometric morphometrics: testing the sensitivity of means and variances in simple randomized selection experiments. Zoomorphology 134, 149–163 (2015).
Mitteroecker, P. & Bookstein, F. Linear discrimination, ordination, and the visualization of selection gradients in modern morphometrics. Evol. Biol. 38, 100–114 (2011).
Polly, P. D. Paleontology and the comparative method: ancestral node reconstructions versus observed node values. Am. Nat. 157, 596–609 (2001).
Mallick, S. et al. The simons genome diversity project: 300 genomes from 142 diverse populations. Nature 538, 201–206 (2016).
Perry, G. H. et al. Adaptive, convergent origins of the pygmy phenotype in African rainforest hunter-gatherers. Proc. Natl Acad. Sci. USA 111, E3596–E3603 (2014).
Grün, R. et al. U-series and ESR analyses of bones and teeth relating to the human burials from Skhūl. J. Hum. Evol. 49, 316–334 (2005).
Lahr, M. M. & Wright, R. V. S. The question of robusticity and the relationship between cranial size and shape in Homo sapiens. J. Hum. Evol. 31, 157–191 (1996).
Eldredge, N. & Gould, S. J. Models in Paleobiology (ed. Schopf, T. J. M.) 82–115 (Freeman, Cooper, San Francisco, 1972).
Tierney, J. E., deMenocal, P. B. & Zander, P. D. A climatic context for the out-of-Africa migration. Geology 45, 1023–1026 (2017).
Schwartz, J. H. & Tattersall, I. Craniodental Morphology of Genus Homo (Africa and Asia). Wiley-Liss edn., Vol. 2 (New York, 2003).
Basell, LS Distribuições de sítios da Idade da Pedra Média (MSA) na África Oriental e sua relação com a mudança ambiental do Quaternário, refúgios e a evolução do Homo sapiens . Quat. ciência Rev. 27 , 2484-2498 (2008).
Martinon-Torres, M., Wu, X., Castro, JM Bd, Xing, S. & Liu, W. Homo sapiens no Pleistoceno Superior da Ásia Oriental. atual Anthr. 58 , S434–S448 (2017).
Liu, W. et al. Os primeiros humanos inequivocamente modernos no sul da China. Natureza 526 , 696 (2015).
Kuhlwilm, M. et ai. Fluxo de genes antigos dos primeiros humanos modernos para os neandertais orientais. Natureza 530 , 429 (2016).
Oppo, DW, McManus, JF & Cullen, JL Evolução e desaparecimento do último calor interglacial no Atlântico Norte subpolar. Quat. ciência Rev. 25 , 3268–3277 (2006).
Henn, BM et al. A diversidade genômica de caçadores-coletores sugere uma origem sul-africana para os humanos modernos. Proc. Natl Acad. ciência EUA 108 , 5154–5162 (2011).
Ramachandran, S. et ai. Suporte da relação de distância genética e geográfica em populações humanas para um efeito fundador serial originário da África. Proc. Natl Acad. ciência EUA 102 , 15942-15947 (2005).
Schlebusch, CM et al. Os genomas antigos da África Austral estimam a divergência humana moderna entre 350.000 e 260.000 anos atrás. Ciência 358 , 652–655 (2017).
Hublin, J.-J. & Klein, R. G. Northern Africa could also have housed the source population for living humans. Proc. Natl Acad. Sci. USA 108, E277 (2011).
Schwartz, GT & Tattersall, I. O queixo humano revisitado: o que é e quem o tem? J. Hum. Evolução 38 , 367–409 (2000).
Reich, D. et al. História genética de um grupo hominídeo arcaico da Caverna Denisova, na Sibéria. Natureza 468 , 1053–1060 (2010).
Meyer, M. et ai. Uma sequência do genoma mitocondrial de um hominídeo de Sima de los Huesos. Natureza 505 , 403–406 (2014).
Wiley, DF Landmark v 3.0. Instituto de Análise e Visualização de Dados . (Universidade da Califórnia, Davis, 2005).
Gunz, P. Reconstrução estatística e geométrica de crânios hominídeos: reconstrução da ontogenia australopitecínea Ph.D. tese, Universidade de Viena (2005).
Bookstein, FL Warps principais: splines de placas finas e a decomposição de deformações. IEEE Trans. Padrão Anal. Mach. Intel. 11 , 567-585 (1989).
Gower, JC Análise generalizada de procrustos. Psychometrika 40 , 33-50 (1975).
Fabre, V., Condemi, S. & Degioanni, A. Genetic evidence of geographical groups among Neanderthals. PLoS ONE 4, e5151 (2009).
Pagel, M. in Morphology, Shape and Phylogeny Systematics Association Special Volumes (eds. MacLeod, N. & Forey, P. L.) 269–286 (CRC Press, London, New York, 2002).
Felsenstein, J. Phylogenies e o método comparativo. Sou. Nat. 125 , 1–15 (1985).
Nunn, CL A Abordagem Comparativa em Antropologia Evolutiva e Biologia . (A imprensa da Universidade de Chicago, 2011).
Weaver, TD, Roseman, CC & Stringer, CB As diferenças cranianas dos homens modernos e neandertais foram produzidas por seleção natural ou deriva genética? J. Hum. Evolução 53 , 135–145 (2007).
Gonzalez-Jose, R., Escapa, I., Neves, WA, Cuneo, R. & Pucciarelli, HM A análise cladística de traços modularizados contínuos fornece sinais filogenéticos na do Homo evolução . Natureza 453 , 775-778 (2008).
Adams, DC, Cardini, A., Monteiro, LR, O'Higgins, P. & Rohlf, FJ Morfometria e filogenética: componentes principais da forma de módulos cranianos não são caracteres cladísticos apropriados nem efetivos. J. Hum. Evolução 60 , 240–243 (2011).
Polly, PD, Lawing, AM, Fabre, A.-C. & Goswami, A. Análise de componentes principais filogenéticos e morfometria geométrica. Hystrix It. J. Mam. 24 , 33–41 (2013).
Rohlf, FJ Métodos comparativos para análise de variáveis contínuas: interpretações geométricas. Evolution 55 , 2143–2160 (2001).
Schlager, S. Reconstrução de tecidos moles do nariz humano: diferenças populacionais e dimorfismo sexual Ph.D. tese, Universität Freiburg (2013).
Adams, DC & Otárola-Castillo, E. Geomorph um pacote R para a coleta e análise de dados de formas morfométricas geométricas. Métodos Eco. Evolução 4 , 393–399 (2013).
Paradis, E., Claude, J. & Strimmer, K. APE: análises de filogenética e evolução em linguagem R. Bioinformática 20 , 289–290 (2004).
Revell, LJ phytools: um pacote R para biologia comparativa filogenética (e outras coisas). Métodos Eco. Evolução 3 , 217–223 (2012).
Dray, S. & Dufour, A.-B. The ade4 package: implementing the duality diagram for ecologists. J. Stat. Soft. 22, 20 (2007).
Goolsby, E. W. Phylogenetic comparative methods for evaluating the evolutionary history of function-valued traits. Syst. Biol. 64, 568–578 (2015).
Copes, L. E. Comparative and Experimental Investigations of Cranial Robusticity in Mid-Pleistocene Hominins. Ph.D Dissertation thesis, Arizona State University, (2012).
Acknowledgements
Este estudo foi parcialmente financiado pela Fyssen Foundation e um Advanced ERC Award (IN-AFRICA Project, ERC 295907). Pela permissão para estudar os espécimes sob seus cuidados, agradecemos aos diretores e curadores das seguintes instituições: Duckworth Collection (Cambridge, Reino Unido); Institut de Paléontologie Humaine (Paris, França); Musée de l'Homme (Paris, França); Museo di Antropologia G. Sergi (Sapienza Università di Roma, Itália); Museu pré-histórico-etonográfico “L. Pigorini” (Roma, Itália); Museu Nacional (Bloemfontein, República da África do Sul); Museus Nacionais do Quênia (Nairóbi, Quênia); Museu de História Natural (Londres, Reino Unido); OR: banco de dados ORSA, Penn Museum (Filadélfia, EUA); Peabody Museum, Harvard University (Cambridge, EUA), American Museum of Natural History (Nova York, EUA). Agradecemos a P. Verdu pela assistência em genômica humana e F. Détroit pela ajuda e ideias. Agradecemos a F. Lahr e F. Rivera pela assistência com a tomografia computadorizada.
Informação sobre o autor
Autores e Afiliações
Contribuições
AM e MML planejaram o estudo, AM coletou os dados e realizou a análise, AM e MML redigiram o artigo.
autor correspondente
Declarações éticas
Interesses competitivos
Os autores declaram não haver interesses conflitantes.
Informação adicional
Informações sobre revisão por pares: A Nature Communications agradece a Andrea Cardini e aos outros revisores anônimos por sua contribuição para a revisão por pares deste trabalho.
Nota do editor: A Springer Nature permanece neutra em relação a reivindicações jurisdicionais em mapas publicados e afiliações institucionais.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.