New paper: media response
I was originally going to post more on the science in my new paper on theropods, but I thought it would be a good opportunity to present my observations on the media response to my paper (or whatever media coverage there was).
The Proceedings of the Royal Society releases their own press materials before the official online release, the abstract for which, I actually wrote myself (200 words) when I submitted the final corrected version of the manuscript. Following the said press release (but before the online release) I got contacted by two journalists, one for Discovery News and another for Australia's ABC News. I was quite impressed by both journalists' questions; they were very good questions. So I did my best to answer as much as I can in the limited time I had (I was in London visiting the Natural History Museum).
The resulting articles that came out on the day of the online release of the paper were quite good. I was pleasantly surprised. If you haven't already, you should have a look at both articles (Discovery News and ABC News). They are quite accurate in the science reporting.
However, having said that, there were a couple of things that amused me; like the phrase 'ripping the heads off of prey'. But annoyingly (though very mildly), I am misquoted in one to have claimed Velociraptor is an ostrich dinosaur. I was actually listing theropod groups and Velociraptor just followed ostrich dinosaurs, separated by a comma. And I don't particularly understand the relevance of mentioning the bite force of Carcharodon megalodon.
All other news stories following these two were just basically copy-paste jobs, with a few minor changes; like me apparently being a British palaeontologist...
There were a few nice blog pieces on this too (here, here and here). So thanks to those bloggers that picked up and commented on my study.
I must say, I was a bit disappointed in the somewhat pervasive phrasing 'unsurprisingly' or 'confirms what palaeontologists have already suspected'; this makes it look like my study was a waste of funding (BTW if you do think this, then you are so wrong about funding...or pretty much a lack thereof). But more importantly, just because everyone thought so, doesn't mean you shouldn't test it. In fact, this type of analysis has never been done before; that's why Proc B thought it was worth their page space. Furthermore, Allosaurus has previously been considered to have weak muscle driven bites, so unique biting strategies have been suggested (Bakker, 1998; Rayfield et al., 2001; Anton et al., 2003), but my results would indicate that Allosaurus and other allosaurs actually had very efficient biting. Of course, the sizes of the jaw adductors are ignored in my study but the it remains that allosaurs are not inefficient biters. One of the other surprises I had was the tendency in allosaurs (especially in the carcharodontosaurs) to have extremely high-efficiency biting at the back of the toothrow. Carcharodontosaurus and Giganotosaurus have always been compared to Tyrannosaurus and people commonly said, 'well Giganotosaurus may have been bigger but T. rex would have ripped its head off!'. Now my study confirms that Tyrannosaurus is a high-efficiency biter but it also shows that carcharodontosaurs had higher efficiency at the back of the tooth row than Tyrannosaurus. That's kind of contrary to what a lot of people have suggested in the past; for instance, Carcharodontosaurus doesn't have strong enough dentition for powerful biting (don't ask me for the source because I don't know; it's one of those 'what palaeontologists have long suspected' type statements that I really don't know where it originated). So personally, I think we should be putting more focus on carcharodontosaur functional morphology.
Anyway, enough of the rant, and thanks to the journalists at Discovery News and ABC News for their quality reporting and also to all that blogged about it!!!
References:
Anton., M., Sanchez, I.M., Salesa, M.J., & Turner, A. 2003. The muscle-powered bite of Allosaurus (Dinosauria: Theropoda): an interpretation of cranio-dental morphology. Estudios Geologicos 59: 313-323.
Bakker, R.T. 1998. Brontosaur killers: Late Jurassic allosaurids as sabre-tooth cat analogues. Gaia 15: 145-158.
Rayfield, E.J., Norman, D.B., et al. 2001. Cranial design and function in a large theropod dinosaur. Nature 409: 1033-1037.
Saturday, June 12, 2010
New paper: jaw biomechanics and the evolution of biting performance in theropod dinosaurs
My new paper on theropod jaw biomechanics was finally published as an early online edition of the Proceedings of the Royal Society B. It became available on Wednesday, 9th June, but I was busy studying cat skulls at the NHM in London and I didn't have much time to comment on it until now. It is a modified version of the study I presented at SVP 2009; although I don't know how many people remember my talk. I made a few revisions to the analyses afterwards (as a response to my reviewers), but the main points are pretty much the same.
This study is pretty much a revamp of my MSc thesis where I compared biting efficiency using a novel numerical method. In my MSc thesis, I looked at how the crushing component of the bite force is affected by jaw margin morphology and how they compare across different theropod taxa. Since it was back in my early days of quantitative comparative analyses, I had no idea (or never occurred to me) how I would go about and compare them numerically (e.g. statistically). So I just basically plotted out bite force profiles and compared them qualitatively (e.g. this squiggly line looks a bit more squiggly than this other squiggly line). Furthermore, after I started my PhD studies, as I was looking back at my MSc material for possible publication, I realised that the calculations were based on an erroneous fundamental assumption; or more strictly speaking, my quantification procedure depended entirely on a subjective orientation of the skull image. Attempts at computing orientation-free force profiles were faced with complications (i.e. the computer scripts didn't work). At that time, I didn't have much computing skills (not that I am any better now) but I needed to come up with another way of quantifying the same thing. But I couldn't really think of any so I kind of gave up and focused on my PhD.

It didn't occur to me until after I defended my PhD thesis that all I needed to do was to look at the patterns of mechanical advantages along the entirety of the tooth row, and I would effectively be looking at something very similar to what I intended in my MSc thesis. Mechanical advantage is an established biomechanical metric and is much easier to compute. Most importantly, mechanical advantage is simply a ratio of the in-lever and the out-lever so it is free of jaw/skull orientations; this solves my problem of orientation. Further, since mechanical advantage is a ratio, it is size-independent as well. This makes it possible to directly compare taxa spanning several orders of magnitude in size.

But using mechanical advantage isn't free of complications. The most prominent when applying to theropods is the issue of homologous biting positions. In mammals, where use of mechanical advantage has been common practice for a very long time, mechanical advantages can be taken at homologous or functionally analogous biting positions, such as the canines, the molars or the carnassials. In theropods, tooth counts are variable, and the only homologous/analogous biting positions are the anterior-most and posterior-most biting positions (any other biting position is not directly comparable; for instance how does a 7th maxillary tooth position of Tyrannosaurus relate to in Allosaurus? Or what if the 'longest mid-maxillary tooth/teeth' aren't immediately obvious?). The only way to compare biting positions in between these two points would be to use a proportional positioning system such as percentages. One can use the position of the tooth in relation to the total length of the jaw but that would have undesirable effects of making the posterior-most biting positions incomparable across taxa with differing tooth row length relative to jaw length. So the most consistent way of standardising biting position is to fit every biting position along a percentage scale of the tooth row length with the posterior-most biting position being the 0% tooth row position and the anterior-most biting position being the 100% tooth row position.
To make it clear here (because one of my reviewers confused this) the standardised biting position is employed after mechanical advantages are computed using absolute distances. The standardisation is to make comparisons of mechanical advantages across taxa possible, not to standardise the mechanical advantages themselves.

The result is that you get a plot of mechanical advantage against standardised biting positions. And you can compare how mechanical advantages change along the tooth row (see figure to the right). To be clear again, the 0% tooth row position at the left hand side of the plot is the tooth at the back of the tooth row. As expected, that is where the biting efficiency is highest in any taxon studied. Towards the right hand side of the horizontal axis, we get further along the tooth row towards the front of the snout. As usual in lever mechanics, towards the tip of the snout, mechanical advantage gets lower, or less efficient. This is kind of like trying to cut something tough with a long pair of scissors; it is easy to cut/crush through a strong twig at the back of the scissors but really difficult at the tip of the scissors.
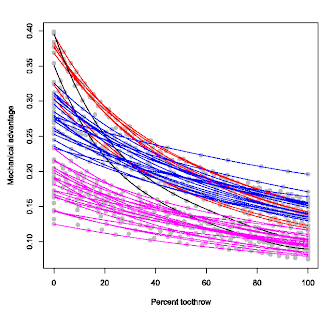 It may occur to some that maybe this profile would be consistent across theropod taxa; after all, they all seem to have longish tooth rows with little variation in tooth row morphology. This is true to some extent that the majority of the profile morphology is relatively similar, in that they are all simple parabolic curves. However, there are major differences in the vertical positioning of the curves, or the absolute values of the mechanical advantages (see figure to the left). This graph is from an older version so it's not exactly the same one in the final paper, but the overall pattern is the same; you see a vertical separation in biting profiles, albeit along a continuum (kind of like a smear I guess). Nonetheless, there are prominently unique profiles. For instance the ones in black, Coelophysis and Syntarsus. These profiles criss-cross the entire vertical spectrum from very high mechanical advantages at the back of the tooth row to very low mechanical advantages at the front of the tooth row. Other theropods do not have this extreme combination.
It may occur to some that maybe this profile would be consistent across theropod taxa; after all, they all seem to have longish tooth rows with little variation in tooth row morphology. This is true to some extent that the majority of the profile morphology is relatively similar, in that they are all simple parabolic curves. However, there are major differences in the vertical positioning of the curves, or the absolute values of the mechanical advantages (see figure to the left). This graph is from an older version so it's not exactly the same one in the final paper, but the overall pattern is the same; you see a vertical separation in biting profiles, albeit along a continuum (kind of like a smear I guess). Nonetheless, there are prominently unique profiles. For instance the ones in black, Coelophysis and Syntarsus. These profiles criss-cross the entire vertical spectrum from very high mechanical advantages at the back of the tooth row to very low mechanical advantages at the front of the tooth row. Other theropods do not have this extreme combination.The ones in red are close but not as extreme. These are represented by the two primitive taxa, Herrerasaurus and Plateosaurus. So this is quite likely the ancestral condition in theropods. Dilophosaurus is also showing a similar profile, consistent with this profile being ancestral (although coelophysoids are an exception). Interestingly, some more derived theropods also share similar profiles with these basal taxa; most notably and surprisingly, Carcharodontosaurus. This type of biting is typified by an extremely high mechanical advantage at the back of the tooth row with relatively high to moderate mechanical advantage at the front of the tooth row. What this reflects is that these taxa have their muscle attachments relatively close to a long tooth row, so the overall mechanical advantage is high at the same time the range in mechanical advantage along the length of the tooth row is high as well. In a way the coelophysoid-type profile is an extreme form of this profile.
The rest of the theropods (the blue and pink ones) are almost indistinct from one another in terms of profile (except for a few), but are different in vertical positions, spread out along a continuum. But there is a noticeable gap, hence the different colour designation. This distinction can be a bit arbitrary so I don't make such a distinction in the paper and is discussed in terms of a gradual spectrum; the high-efficiency end of the spectrum towards the top and the low-efficiency (weak/fast biting) end towards the bottom. The high-efficiency function types (blue) have relatively high mechanical advantages along short tooth rows, while conversely the weak/fast function types (pink) have low mechanical advantages along their short tooth rows.
At this point we are still comparing squiggliness of squiggly lines, so in my next post, I shall introduce what I did in my paper in order to make a more meaningful comparison.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.