Taxonomia e filogenia da hominina: o que há em um nome?



A prática da classificação biológica apareceu no início da evolução da vida: é seguro comer / não seguro comer ? Isso é um predador / não um predador ? Esses parceiros potenciais são / não são parceiros potenciais ? Está em nossa natureza, então, classificar nosso ambiente. No contexto da biologia, essa prática assume importância especial, à medida que cientistas de muitas disciplinas biológicas trabalham em direção a um único sistema de classificação que incorpora todos os organismos que já viveram. Tanto a prática quanto o produto dessa grande classificação biológica são chamados de taxonomia .
O sistema taxonômico mais amplamente usado foi formalizado por Carolus Linnaeus e compreende uma hierarquia aninhada simples em que organismos semelhantes se agrupam em uma classificação taxonômica , e esses grupos se agrupam em grupos sucessivamente mais amplos em classificações superiores. A Figura 1 mostra uma taxonomia de ancestrais humanos e seus parentes macacos africanos existentes. Essa hierarquia aninhada permite que diferentes graus de similaridade sejam representados em diferentes classificações. Observe, entretanto, que a taxonomia Linnaeana antecede a teoria evolucionária moderna; Considerando que Linnaeus organizou os organismos vivos de acordo com diferentes níveis de similaridade, foram Darwin e outros que explicaram esses diferentes graus de similaridade por meio de ancestrais comuns. Quanto mais próximos dois grupos forem, mais semelhanças eles provavelmente compartilharão. A série de relações evolutivas entre um grupo de organismos é denominada filogenia .

A relação entre taxonomia e filogenia
É comumente entendido que a taxonomia deve refletir a filogenia - os organismos devem ser agrupados de acordo com sua história evolutiva, sua relação. Isso faz sentido intuitivo porque a ancestralidade comum é o único fator que une quatro bilhões de anos de vida na Terra. Essa conexão entre taxonomia e filogenia significa que novas descobertas ou outros dados que mudam nossa compreensão da história evolutiva frequentemente resultam na mistura de táxons e de nomes taxonômicos.
Um excelente exemplo desse embaralhamento é a mudança no uso do termo "hominídeo". Tradicionalmente, apenas ancestrais humanos foram colocados na família Hominidae (e, portanto, chamados de hominídeos). Isso refletia uma visão de que os humanos são substancialmente diferentes dos grandes macacos, que foram colocados na Família Pongidae (pongídeos) (Figura 2a). No entanto, evidências genéticas esmagadoras têm demonstrado que humanos, chimpanzés e gorilas são muito mais intimamente relacionados entre si do que com o orangotango (por exemplo, Sarich, 1971; Caccone & Powell, 1989; Ruvolo, 1994). Portanto, não há suporte genético para agrupar os grandes macacos em um grupo distinto do homem. Por esta razão, muitos pesquisadores agora colocam todas as espécies de grandes símios e humanos em uma única família, Hominidae - tornando-os todos "hominídeos" adequados (Figura 2b).
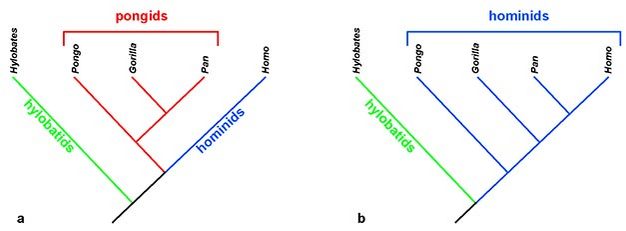
Essas
mudanças taxonômicas também têm efeitos a jusante: macacos e humanos
africanos agora são distinguidos dos orangotangos na classificação da
subfamília Homininae (hominídeos), e a linhagem humana é separada em uma
classificação ainda mais baixa da Tribo Hominini. Assim, o termo comum
para a linhagem de humanos fósseis e modernos é "hominídeo". Além disso,
o termo tradicional "australopithecine", que agrupava Australopithecus e Paranthropus
na subfamília Australopithecinae, torna-se inválido sob esta taxonomia
revisada; no contexto da taxonomia de Lineu, não se pode aninhar uma
subfamília de classificação superior (Australopitecíneo) dentro de uma
tribo de classificação inferior (Hominini) (ver Figura 1).
Por
um lado, essa mudança na terminologia ao longo dos anos ilustra o
avanço adequado da ciência, por meio do qual novas evidências sobre a
história evolutiva ajudam a refinar nossos modelos taxonômicos. No entanto, a mudança linguística necessária demorou a ganhar aceitação entre alguns pesquisadores. Da
mesma forma, a popularidade de termos como “hominídeo” e
“australopitecíneo”, que se espalharam pela consciência pública nos anos
1960-70, significa que esses termos provavelmente persistirão em seus
significados tradicionais para comunicação com o público.
Taxonomia e filogenia da hominina
Um modelo de evolução dos hominídeos é ilustrado pela filogenia na Figura 3, apresentada aqui como um veículo para discussão ao invés de uma hipótese robusta. Um óbvio ponto de discórdia é o status dos primeiros hominíneos, conhecido apenas por pequenas amostras isoladas; as relações entre essas espécies e os hominíneos posteriores ainda não foram resolvidas. E, enquanto outros ramos da árvore evolutiva humana são vigorosamente debatidos na literatura, os sinais usuais de incerteza filogenética (por exemplo, linhas tracejadas, ramos alternados, pontos de interrogação) foram omitidos da Figura 3 para simplificar. Em vez disso, algumas dessas questões são destacadas abaixo, especificamente no contexto de obstáculos taxonômicos e filogênicos: ancestralidade, variação e complexidade biológica.
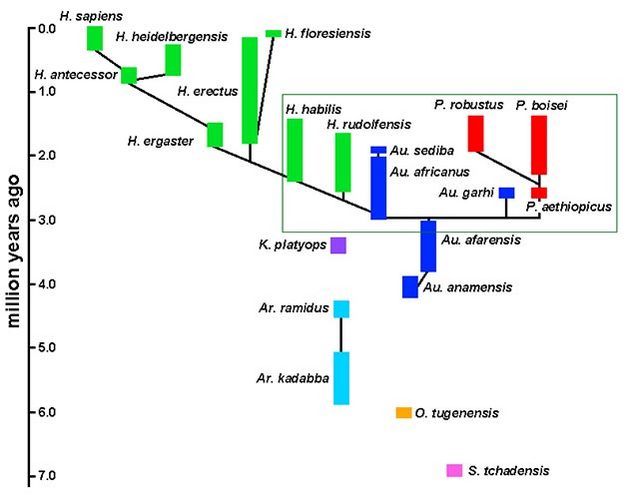
Australopithecus e o problema da ancestralidade
Cinco espécies de Australopithecus são reconhecidas aqui, embora novas descobertas provavelmente aumentem este número. Há boas evidências de que A. anamensis e A. afarensis representam evolução dentro de uma linhagem ampla (Kimbel et al ., 2006), mas outras relações no gênero não são bem compreendidas. Pode ser que o gênero seja um amálgama de espécies de caule que não estão intimamente relacionadas (veja abaixo), embora pareçam compartilhar um grau adaptativo comum . A maioria dos pesquisadores concordaria que o Paranthropus e o Homo evoluíram de alguns Australopithecuscomo ancestral (es), embora isso ainda seja controverso. Essa ideia de ancestralidade, no entanto, apresenta dificuldades para nossos esforços em casar a taxonomia com a filogenia. Por exemplo, se a filogenia na Figura 3 estivesse correta, então A. africanus e seus descendentes estariam mais intimamente relacionados ao gênero Homo do que outras espécies de Australopithecus ; da mesma forma, A. garhi seria mais parente do Paranthropus . Como, então, nossa taxonomia pode ser feita para refletir essas relações evolutivas?
A solução mais comum é a divisão. Por exemplo, a Figura 4a ilustra as relações cladogenéticas entre as espécies em caixas na Figura 3, com espécies de " Australopithecus " aqui dispersas entre três gêneros. Porque o nome do gênero Australopithecus está especificamente associado ao Au. africanus , esta espécie e seu descendente Au. sediba mantém esse nome de gênero. " A. " garhi é transferido para Paranthropus para refletir essa relação, enquanto " A. " afarensis requer uma nova designação de gênero, Praeanthropus (ver, por exemplo, Strait et al., 2007). Essa divisão horizontal em nível de espécie de nossa taxonomia também implica uma divisão vertical em classificações mais altas, onde cada nó (junção de ramos) em nosso cladograma requer uma classificação distinta para denotar uma relação evolutiva separada. Assim, para replicar exatamente as relações mostradas no cladograma, seria necessário adicionar seis classificações adicionais (e seus nomes de táxons correspondentes ) entre os níveis de tribo e espécie que representam os eventos de divisão histórica nesta linhagem. Dessa forma, uma hierarquia taxonômica pode ser feita para refletir precisamente as relações cladogenéticas da história evolutiva.
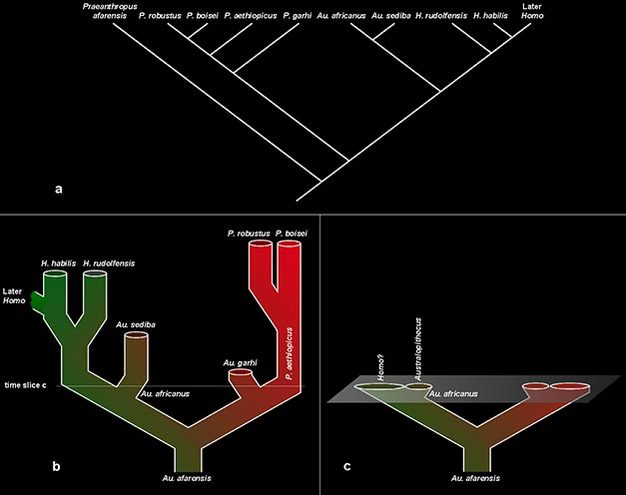
Essa abordagem leva a um curioso dilema teórico, entretanto. Ao organizar nossos nomes taxonômicos para corresponder às relações filogenéticas, acabamos organizando as espécies de acordo com eventos que aconteceram depois de sua existência, em vez de sua biologia contemporânea. Considere, por exemplo, a filogenia na Figura 4b, representando essas populações evoluindo ao longo do tempo. Considerando uma fatia de tempo dessa filogenia (Figura 4c), as duas populações de Au. africanus seria - por qualquer medida contemporânea - a mesma espécie. Mas, porque uma população posteriormente evoluiu para espécies de Homo, pesquisadores em uma posição moderna tendem a transferir essa população para este gênero. Tal ação pode refletir melhor as relações filogenéticas subsequentes entre os descendentes, mas não leva em conta totalmente o contexto biológico dos organismos em questão.
Homo primitivo e o problema da variação
A divisão entre australopitecos e o gênero Homo já foi bastante clara, o último grupo distinguido por um cérebro notavelmente maior. A adição de Leakey et al . (1964) ao Homo do H. habilis de cérebro menor , entretanto, obscureceu essa fronteira e gerou um debate sobre o escopo da variação que poderia ser acomodada dentro do gênero humano. Com a descoberta de muito mais espécimes do Homo primitivo , entretanto, esses argumentos deram lugar a outras considerações taxonômicas: quantas espécies foram representadas na amostra diversa de Homo de cérebro pequeno ? eles poderiam ser acomodados dentro da única espécie H. habilis ?
Entre os fósseis, a variação dentro de uma espécie é amplamente uma questão taxonômica, em vez de filogenética, simplesmente porque a preservação fóssil raramente é boa o suficiente para desenvolver hipóteses evolutivas robustas para um grande número de espécimes individuais. Infelizmente, não há diretrizes claras quanto à quantidade de variação apropriada para qualquer grupo taxonômico, e nenhuma maneira definitiva de testar se as alocações de espécies são apropriadas. Os pesquisadores geralmente usam a variação dentro das espécies modernas como referência para intervalos e padrões de variação aceitáveis (por exemplo, McNulty, 2003, 2005; Baab et al., 2010), mas mesmo isso pode ser problemático: a variação em espécies extintas não precisa ser semelhante à variação em espécies modernas (Kelley, 1993). Este problema pode ser especialmente agudo em estudos com hominídeos, onde a variação em espécies comparativas vivas, como gorilas e chimpanzés, pode ter sido radicalmente impactada pelo declínio das populações.
Qual é, então, a natureza da variação no Homo primitivo ? Uma série de comparações com macacos modernos e humanos descobriram que a amostra tradicionalmente agrupada em H. habilis excedeu a variação esperada dentro de uma única espécie moderna (por exemplo, Stringer 1986; Bilsborough e Wood 1988; Lieberman et al . 1988; Chamberlain 1989; Kramer et al. 1995). Isso levou Alexeev (1986) a propor uma nova espécie, H. rudolfensis , para acomodar os espécimes maiores desta amostra, mantendo os menores em H. habilis . Com uma espécie grande e uma pequena espécie de Homo primitivo , o contra-argumento lógico era que a amostra representa uma única espécie sexualmente dimórficaespécies (por exemplo, Howell 1978); Wood et al. (1991), entretanto, forneceram evidências provisórias de que os padrões de variação dentro da ampla amostra do Homo inicial não são consistentes com os padrões conhecidos de dimorfismo. Por esta razão, os paleoantropólogos têm amplamente aceito a presença de duas espécies de Homo primitivos , e pode haver evidências de diversidade taxonômica adicional (Smith e Grine, 2008).
Como uma reviravolta final na narrativa taxonômica, Wood e Collard (1999) forneceram uma justificativa coerente para remover H. habilis e H. rudolfensis do Homo inteiramente, e colocar um ou ambos no Australopithecus (ver também Leakey et al ., 2001). Até agora, porém, sua proposta não obteve ampla aceitação.
Neandertais e o problema da complexidade biológica
Um dos debates mais antigos da paleoantropologia gira em torno do status taxonômico dos neandertais. Enquanto a maioria dos pesquisadores concorda que a morfologia neandertal distinta indica um grau de separação de outros hominíneos contemporâneos, há discordância sobre se os neandertais pertencem ao H. sapiens ou à sua própria espécie, H. neanderthalensis . Há ampla justificativa morfológica para o reconhecimento de uma espécie distinta (por exemplo, Harvati et al ., 2004), e as primeiras análises de DNA mitocondrial antigo de neandertais apoiaram essa conclusão (por exemplo, Krings et al ., 1997). Mais recentemente, no entanto, o sequenciamento do genoma nucleardemonstrou que as populações humanas modernas fora da África retêm até 4% dos genes neandertais distintos (Green et al ., 2010). Se alguém define uma espécie como populações de organismos que se cruzam (um critério conhecido como o conceito biológico de espécie ), então a presença de genes neandertais entre os humanos modernos argumenta fortemente para incluí-los dentro de H. sapiens , porque implica que neandertais contemporâneos e humanos anatomicamente modernos estavam cruzando.
O caso dos neandertais demonstra um obstáculo final na prática padrão de taxonomia e filogenia: a biologia nem sempre adere aos nossos modelos simplistas. Nossa preferência por arbitrar grupos taxonômicos discretos e divisões filogenéticas instantâneas ignora a continuidade e a complexidade da evolução biológica. Na verdade, uma das demonstrações mais convincentes de evolução e ancestralidade comum é precisamente a falta de demarcações estritas entre espécies intimamente relacionadas: cães e lobos cruzam; diferentes espécies de macacos hibridizam; até mesmo a mula (ou hinny) pode produzir descendentes, embora raramente. Esses limites borrados são exatamenteo que seria esperado de populações que evoluem continuamente e divergem ao longo do tempo. Essa realidade biológica, no entanto, destrói as noções de que as linhagens evolucionárias se dividem instantaneamente, ou de que podemos circunscrever grupos de organismos perfeitamente em grupos taxonômicos distintos.
Os neandertais se dividiram em espécies diferentes ou eram parte de nossa linhagem direta? A resposta é provavelmente "ambos". O isolamento reprodutivo e, portanto, a especiação, é um processo pelo qual as populações experimentam graus de infertilidade - impedimentos comportamentais e estruturais, redução da viabilidade dos híbridos, incompatibilidade gamética etc. - ao longo de muitas gerações. Nesse sentido, as linhagens podem não se dividir tanto quanto reticular, com a rede de trocas genéticas entre as populações se dissipando com o tempo. Mesmo em neandertais, que provavelmente estavam geograficamente isolados da linhagem humana moderna, a capacidade de cruzar foi persistente por centenas de milhares de anos (Green et al., 2010). No entanto, com o DNA de neandertal fornecendo talvez a única oportunidade de testar diretamente se um hominíneo pré-moderno extinto cruzou com o moderno H. sapiens , estou inclinado aqui a agrupar neandertais em nossa espécie.
A importância da taxonomia na paleoantropologia
Estudamos a ancestralidade humana não apenas para ter rótulos para exposições em museus, mas para iluminar a complexa história de mudanças morfológicas, fisiológicas, comportamentais, cognitivas e culturais que deram origem aos humanos modernos. Assim, a taxonomia em si não é de interesse direto. Juntamente com seu uso principal como um substituto para a filogenia, a taxonomia pode parecer fácil de desconsiderar. Na verdade, se alguém trabalhasse apenas com espécies modernas - que têm anatomias completas, sequências de DNA, comportamentos - poderia ser possível confiar inteiramente nas relações filogenéticas para organizar os espécimes em grupos significativos.
Não é assim no estudo de fósseis, no entanto, onde encontrar qualquer remanescente de espécies antigas é raro e encontrar peças que estão relativamente intactas é extraordinário. Na falta de anatomia suficiente para análises filogenéticas robustas de cada fragmento ósseo em um estudo, são as hipóteses taxonômicas que formam a base biológica para organizar as amostras fósseis. Dentro da paleoantropologia, então, a taxonomia ainda desempenha três papéis cruciais:
1. Alfa-taxonomia . A classificação de espécimes individuais em espécies é talvez o papel mais importante da taxonomia na paleoantropologia moderna. Em uma amostra de fragmentos ósseos que inclui diferentes peças de anatomia, bem como indivíduos de diferentes tamanhos, idade, sexo, antiguidade geológica e talvez espécies, é a taxonomia alfa que fornece a justificativa para unir alguns espécimes em grupos enquanto separa outros - geralmente por meio de referência a modelos existentes robustos de variação dentro da espécie. É importante ressaltar que, como quase todas as pesquisas paleoantropológicas são baseadas em hipóteses taxonômicas, explícita ou implicitamente, as mudanças na taxonomia subjacente têm a capacidade de impactar fortemente nossa interpretação da evolução humana.
2. Mediando comparações biologicamente significativas no espaço / tempo . Além do nível de espécie, as análises taxonômicas fornecem um mecanismo para comparar grupos mais amplos de organismos fósseis. Trabalhar no registro fóssil significa analisar amostras de diferentes lugares, diferentes épocas e conhecidas em diferentes resoluções filogenéticas. No entanto, essa resolução determina a coerência biológica dos grupos filogenéticos: uma seção altamente resolvida de uma filogenia pode incluir espécies fósseis intimamente relacionadas, enquanto uma seção não resolvida pode agrupar uma gama muito maior de organismos. As hipóteses taxonômicas permitem que os pesquisadores mantenham algum nível de uniformidade nas comparações entre grupos tão grandes, independentemente da resolução filogenética.
3. Comunicação . Um dos maiores impactos da taxonomia de Lineu foi facilitar a comunicação na comunidade científica. Isso ainda é importante hoje, e atualmente não há um veículo amplamente aceito para substituí-lo. A terminologia básica da taxonomia também é reconhecida pelo público em geral, e é por meio desses termos que a pesquisa se torna amplamente disponível. Se o conhecimento gerado por meio de pesquisas em milhões de anos de evolução humana deve ter algum impacto além das paredes da academia, então os rótulos taxonômicos tradicionais provavelmente manterão um lugar na paleoantropologia.
O mito da mutação da filogenia hominina
Uma das queixas comuns dirigidas aos paleoantropólogos é que eles estão constantemente reorganizando a árvore evolucionária humana. Na verdade, parece que todo novo fóssil significativo tem o propósito de "mudar completamente nossa compreensão da evolução humana". Na realidade, entretanto, a filogenia dos hominíneos tem sido relativamente estável nas últimas duas décadas, apesar de novas descobertas significativas e controversas. A comparação das filogenias publicadas no início da década de 1990 com as publicadas em 2012 mostra um nível notável de conformidade. Isso sugere que - pelo menos em linhas gerais - nossas hipóteses sobre a evolução da linhagem humana são bastante robustas.
Considere novamente a amostra de espécies de hominídeos ao longo do tempo, mas concentrando-se nos três gêneros bem amostrados: Australopithecus , Paranthropus e Homo (Figura 5). Amostras fósseis substanciais de cada um têm se acumulado por quase um século - mais, no caso do fóssil Homo . A história dos estudos sobre esses espécimes gera uma grande dose de inércia estabilizadora para as relações básicas mostradas aqui. Embora as conexões específicas entre esses grupos, bem como as relações filogenéticas dentro deles, ainda estejam longe de serem resolvidas, o arranjo geral do Australopithecus , Paranthropus e Homoparece relativamente estável; na verdade, quaisquer mudanças neste arranjo provavelmente serão, bem, taxonômicas.
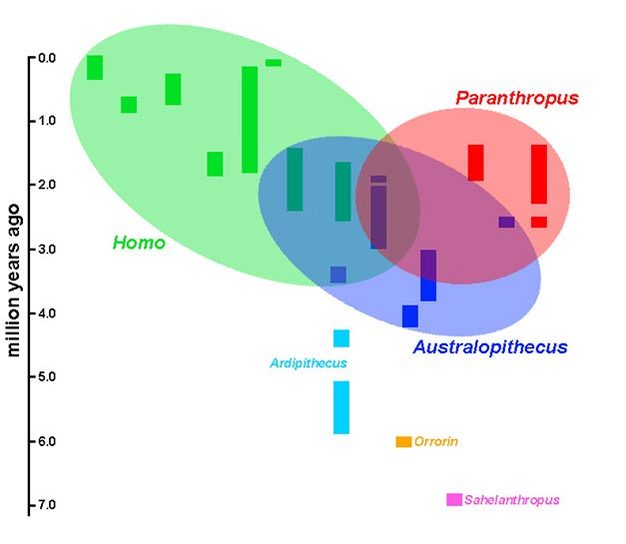
Dito isso, quais aspectos da pesquisa paleoantropológica têm maior probabilidade de ter um grande impacto em nosso conhecimento das relações evolutivas humanas? A resposta pronta vem de novas descobertas. Achados como o fóssil "hobbits" em Flores ( H. floresiensis ) e o enigmático Homoespécies de Dmanisi têm implicações dramáticas para a evolução humana, e o significado delas ainda não impactou totalmente a filogenia das homininas. O status dos primeiros hominídeos também está em um estado de fluxo, e novas descobertas na base da filogenia dos hominídeos certamente serão sentidas nos ramos superiores. Talvez o maior potencial para revisar nossa interpretação da evolução humana, entretanto, venha de alguns dos taxa mais conhecidos. Referindo-se novamente à Figura 5, as elipses foram desenhadas de modo a abranger debates taxonômicos e filogenéticos em andamento relevantes para cada gênero. Como um dispositivo heurístico, então, as regiões onde essas elipses se sobrepõem provavelmente produzirão o maior impacto em nossa compreensão da evolução humana. Resolvendo essas relações - determinando quais espécies pertencem a quais gêneros,e como esses gêneros estão relacionados - tem o potencial de transformar nossa compreensão das origens e diversificação humanas.
Glossário
Australopiths : um termo geral normalmente usado em referência às espécies de Australopithecus e Paranthropus . Este não é um nome taxonômico formal e, portanto, não precisa aderir às regras estritas da nomenclatura biológica.
Conceito de espécie biológica : Esta é a definição mais comumente usada de uma "espécie", por meio da qual populações de organismos são determinadas como pertencendo à mesma espécie se seus membros cruzarem e produzirem descendentes viáveis.
Cladogenético : adjetivo referente à evolução por meio de eventos de cisão. Se uma população diverge em dois ramos que depois evoluem independentemente um do outro, isso é chamado de cladogênese (ou evolução cladogenética). Se uma população evolui ao longo do tempo sem se dividir em vários ramos, isso é chamado de anagênese (ou evolução anagenética).
Cladograma : um diagrama de ramificação que representa apenas as relações filogenéticas entre os organismos (ou seja, não inclui a idade geológica ou outras informações contextuais). Formalmente, um cladograma é produzido por meio da análise cladística de um grupo de organismos, que determina as relações evolutivas usando apenas características derivadas (evoluídas) que são compartilhadas entre pelo menos alguns membros de um grupo.
Ancestralidade comum : a história evolutiva compartilhada de todos os organismos vivos de volta a uma única origem.
Incompatibilidade gamética : uma barreira para a reprodução sexual onde as células sexuais (gametas) de um homem (esperma) e uma mulher (óvulo) são muito diferentes para permitir a fertilização. Uma vez que duas populações tenham divergido e comecem a evoluir independentemente, eventualmente seus membros se tornarão incapazes de cruzar devido à incompatibilidade gamética. Um exemplo disso pode ser se a evolução resultar em diferentes números de cromossomos nas duas populações, de forma que seus membros não possam cruzar (observe que diferentes números de cromossomos nem sempre levam à incompatibilidade gamética).
DNA mitocondrial : informação genética associada à mitocôndria de uma célula, e não a seus cromossomos. As mitocôndrias residem fora do núcleo da célula (dentro do citoplasma) e são herdadas apenas da linha materna em organismos que se reproduzem sexualmente.
Nó : relacionamentos evolutivos são freqüentemente representados por padrões semelhantes a árvores de linhagens ramificadas. Um nó é onde dois ou mais ramos se unem em um ponto e, portanto, representa o último ancestral comum compartilhado por esses grupos antes de eles divergirem.
Genoma nuclear : o genoma nuclear é composto pelo DNA dos cromossomos de um organismo, que residem no núcleo da célula. Em criaturas que se reproduzem sexualmente, o genoma nuclear contém genes de ambos os pais.
Filogenia : as relações evolutivas entre, ou história evolutiva de, um grupo de organismos.
Sexualmente dimórfico : Frase adjetival que se refere às diferenças de tamanho e forma entre machos e fêmeas de uma espécie.
Táxon : Qualquer grupo de qualquer nível da classificação Linnaean pode ser geralmente referido como um táxon. Assim, a Família Hominidae é um táxon, assim como a espécie H. sapiens e a Ordem Primatas. O plural é taxa .
Classificação taxonômica : refere-se a diferentes níveis da classificação hierárquica Linnaeana. Por exemplo, espécie é uma classificação taxonômica; reino , filo e classe são outras categorias.
Taxonomia : a classificação dos organismos vivos, normalmente usando o sistema hierárquico inventado por Carolus Linnaeus.
Fonte: https://www.nature.com/scitable/knowledge/library/hominin-taxonomy-and-phylogeny-what-s-in-142102877/
Referências e leituras recomendadas
Alexeev, V. P. The Origin of the Human Race. Moscow: Progress Publishers, 1986.
Baab, K. L., McNulty, K. P., et al. A comprehensive comparison of the Homo floresiensis cranium to pathological humans and extinct hominins. PLOS One 8, e69119.
Bilsborough, A., & Wood, B. A. Cranial morphometry of early hominids I. Facial region. American Journal of Physical Anthropology 76, 61-86 (1988).
Caccone, A., & Powell, J. R. DNA divergence among hominoids. Evolution 43, 925-942 (1989).
Chamberlain, A.T. Variations within Homo habilis. In Hominidae: Proceedings of the 2nd International Congress of Human Paleontology. ed. Giacobini, G. (Milan: Jaca Books, 1989). 175-181.
Green, R. E., Krause, J., et al. A draft sequence of the neandertal genome. Science 328, 710-722 (2010).
Harvati, K., Frost, S. R., et al. Neanderthal taxonomy reconsidered: Implications of 3D primate models of intra- and interspecific differences. Proceedings of the National Academy of Sciences, USA 101, 1147-1152 (2004).
Howell, F. C. Hominidae. In Evolution of African Mammals. eds. Maglio, V. J., & Cooke, H.B.S. (Cambridge: Harvard University Press, 1978). 154-258.
Kelley, J. Taxonomic implications of sexual dimorphism in Lufengpithecus. In Species, Species Concepts and Primate Evolution. eds. Kimbel, W. H. & Martin, L. B. (New York: Plenum Press, 1993). 429-458.
Kimbel, W. H. , Lockwood, C. A., et al. Was Australopithecus anamensis ancestral to A. afarensis? A case of anagenesis in the hominin fossil record. Journal of Human Evolution 51, 134-152 (2006).
Kramer, A., Donnelly, S. M., et al. Craniometric variation in large-bodied hominoids: testing the single-species hypothesis for Homo habilis. Journal of Human Evolution 29, 443-462 (1995).
Krings, M., Stone, A., et al. Neandertal DNA sequences and the origin of modern humans. Cell 90, 19-30 (1997).
Leakey, L. S. B., Tobias, P. V., et al. New species of the genus Homo from Olduvai Gorge. Nature 202, 7-9 (1964).
Leakey, M. G., Spoor, F., et al. New hominin genus from Eastern Africa shows diverse middle Pliocene lineages. Nature 410, 433-440 (2001).
Lieberman, D. E., Pilbeam, D. R., et al. A probabilistic approach to the problem of sexual dimorphism in Homo habilis: a comparison of KNM-ER 1470 and KNM-ER 1813. Journal of Human Evolution B, 503-511 (1988).
McNulty, K. P. Geometric morphometric analyses of extant and fossil hominoid craniofacial morphology. Unpubl. Ph.D. thesis. City University of New York, New York (2003).
McNulty, K. P. A geometric morphometric assessment of the hominoid supraorbital region: Affinities of the Eurasian Miocene hominoids Dryopithecus, Graecopithecus, and Sivapithecus. In Modern Morphometrics in Physical Anthropology. ed. Slice, D. (New York: Kluwer Academic Publishers, 2005). 349-373.
Ruvolo, M. Molecular evolutionary processes and conflicting gene trees: the hominoid case. American Journal of Physical Anthropology 94, 89-113 (1994).
Sarich, V. M. A molecular approach to the question of human origins. In Background for Man. ed. Dolhinow, P. (Boston: Little, Brown, 1971). 60-81.
Smith, H. F., & Grine, F. E. Cladistic analysis of early Homo crania from Swartkrans and Sterkfontein, South Africa. Journal of Human Evolution 54, 684-704 (2008).
Strait, D., Grine, F. E. et al. Analyzing hominid phylogeny. In Handbook of Paleoanthropology, vol. 3. eds. Henke, W., & Tattersall, I. ( Berlin: Springer, 2007). 1781-1806.
Stringer, C. B. The credibility of Homo habilis. In Major Topics in Primate and Human Evolution . eds. Wood, B. A., Martin, L. B., et al. (Cambridge: Cambridge University Press, 1986). 266-294.
Wood, B. A., & Collard, M. The changing face of the genus Homo. Evolutionary Anthropology, 8, 195-207 (1999).
Wood, B. A., Yu, L., et al. Intraspecific variation and sexual dimorphism in cranial and dental variables among higher primates and their bearing on the hominid fossil record. Journal of Anatomy 174, 185-205 (1991).
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.