A maior cobra madtsoiida conhecida do período Eoceno quente da Índia sugere dispersão intercontinental de Gondwana
Relatórios Científicos volume 14 , Artigo número: 8054 ( 2024 )
Abstrato
Aqui relatamos a descoberta de fósseis representando a coluna vertebral parcial de uma cobra gigante madtsoiida de uma sucessão de linhita do início do Eoceno Médio (Lutetiano, ~ 47 Ma) em Kutch, oeste da Índia. O comprimento estimado do corpo de ~ 11–15 m torna este novo táxon ( Vasuki indicus gen et sp. nov.) a maior cobra madtsoiida conhecida, que prosperou durante um intervalo geológico quente com temperaturas médias estimadas em ~ 28 °C. Filogeneticamente, Vasuki forma um clado distinto com o táxon indiano do Cretáceo Superior Madtsoia pisdurensis e o norte-africano do Eoceno Superior Gigantophis garstini . Considerações biogeográficas, vistas em conjunto com a sua inter-relação com outros madtsoiídeos indianos e norte-africanos, sugerem que Vasuki representa uma linhagem relíquia que se originou na Índia. A subsequente colisão Índia-Ásia em ~ 50 Ma levou à dispersão intercontinental desta linhagem do subcontinente para o Norte da África através do sul da Eurásia.
Conteúdo semelhante sendo visualizado por outras pessoas
Introdução
Madtsoiidae são um clado extinto de cobras terrestres principalmente de Gondwana com uma faixa temporal que abrange cerca de 100 milhões de anos do Cretáceo Superior ao Pleistoceno Superior. 1 , 2 , 3 . A sua distribuição geográfica durante o Cretáceo Superior abrangia Madagáscar, América do Sul, Índia, África e o arquipélago europeu. 1 , 4 , 5 , 6 , 7 , 8 , 9 . As formas Cenozóicas estão restritas ao Norte da África, América do Sul, subcontinente indiano e Austrália 2 , 10 , 11 , 12 , 13 , 14 , 15 , 16 , 17 . Os Madtsoiídeos exibem um amplo espectro de tamanhos corporais e incluem algumas das maiores cobras terrestres conhecidas que já existiram. 2 , 7 , 9 . Embora seja um clado específico, a maioria dos táxons são conhecidos exclusivamente pelas vértebras, resultando em relações intragrupo pouco restritas. 2 , 8 , 16 . Além disso, a posição filogenética de Madtsoiidae dentro de Ophidia permaneceu controversa, já que alguns estudos o recuperam dentro de Serpentes, enquanto outros o colocam fora do grupo da coroa. 3 , 9 , 17 , 18 , 19 , 20 . Essas incertezas filogenéticas dificultaram nossa compreensão da biogeografia e dos eventos de radiação dos madtsoiídeos 2 , 8 .
No subcontinente indiano, os madtsoiídeos do Cretáceo Superior (Maastrichtiano) são conhecidos da província vulcânica de Deccan, incluindo o grande tamanho Madtsoia pisdurensis da Formação Lameta 6,8. Among Tertiary madtsoiids, indeterminate forms are known from the early Paleocene Khadro Formation (Pakistan16) and the early Eocene Cambay Shale (India15). The latter also yielded the large madtsoiid Platyspondylophis21. The Eocene and Late Oligocene records include indeterminate taxa from Kutch and Ladakh, respectively14,22. Here we report the discovery of a giant madtsoiid snake, one of largest snakes ever reported, from an interval corresponding to a warm Middle Eocene period (~ 47 Ma) of India. Fossils were collected from an early Lutetian grey shale unit from Panandhro Lignite Mine, Kutch, Gujrat State, western India (Supplementary Note 1, Fig. 1), and includes an excellently preserved, partial vertebral column. The discovery of a giant Eocene snake has important implications for madtsoiid biogeography in the context of Gondwanan inter-continental dispersal, and the evolution of large body-sizes possibly driven by high temperatures in the Middle Eocene tropical zones.
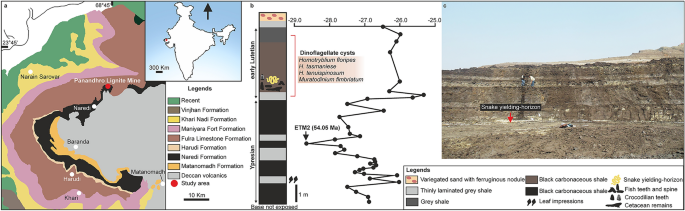
Mapa geológico da Bacia Kutch mostrando a localidade fóssil ( a ); coluna estratigráfica na mina de linhita Panandhro mostrando a posição do horizonte de produção de cobras madtsoiid com assembléia de cistos dinoflagelados para diagnóstico de idade e δ 13 Curva C marcando evento hipertérmico ETM2 (modificado após Agrawal et al. 23 ) ( b ); vista panorâmica do sítio fóssil ( c ). O mapa e a coluna estratigráfica foram desenhados por DD usando CorelDRAW 2019 (número de versão: 21.0.0.593, link URL: http://www.corel.com/en/ ). Estimativa de idade do ETM2 após Westerhold et al. 24 .
Resultados
Paleontologia sistemática
Squamata Oppel, 1811
Ophidia Brongniart, 1800
Madtsoiidae (Hoffstetter 1961) McDowell, 1987
Vasuki indicus gen. et sp. nov.
Etimologia
Nome genérico em homenagem à conhecida serpente mítica hindu 'Vāsuki' em volta do pescoço do Senhor Shiva; o nome específico é para o país de origem, ou seja, Índia.
Holótipo
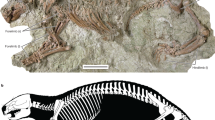
IITR/VPL/SB 3102-1-21; uma coluna vertebral parcial representando a região pré-cloacal (Figs. 2 , 3 ; Tabela Suplementar 1 ).

Vértebras anteriores do tronco de Vasuki indicus . IITR/VPL/SB 3102-3, vértebra parcial em vista anterior ( a ); vista posterior ( b ); vista lateral esquerda ( c ); vista dorsal ( d ); vista ventral ( e ). IITR/VPL/SB 3102-5, vértebra completa em vista anterior ( f ); vista posterior ( g ); vista lateral esquerda ( h ); vista dorsal ( eu ); vista ventral ( j ). IITR/VPL/SB 3102-7I-II, vértebra parcial em vista anterior ( k ); vista posterior; ( eu ); vista lateral esquerda ( m ); vista dorsal ( n ); vista ventral ( o ). IITR/VPL/SB 3102-6, vértebra posterior posterior completa do tronco em vista anterior ( p ); vista posterior ( q ); vista lateral esquerda ( r ); vista dorsal( s ); vista ventral ( t ). As setas cinza indicam a direção anterior. Setas e setas vermelhas indicam fossas na base espinhal neural e forames endozigântricos, respectivamente. Os algarismos romanos nas figuras ( m – o ) referem-se a vértebras individuais em espécimes articulados onde 'I' está voltado para a parte anterior. Ponta de seta branca e seta indicam fossa medial à diápófise e forame na superfície dorsal do arco neural. co cotyle, cn condyle, da diapophysis, hyp hypapophysis, izr interzygapophyseal ridge, msf median shaft, nc neural canal, nrl neural arch lamina, ns neural spine, pa parapophysis, pcof paracotylar foramen, pcofo paracotylar fossa, pcon paracotylar notch, po postzygapophysis, pr prezygapophysis, psl prespinal lamina, pzgf parazygantral foramen, pzgfo parazygantral fossa, scf subcentral foramen, scfo subcentral fossa, zg zygantrum, zs zygosphene. Scale bar represents 50 mm.

Vértebras pré-cloacais de Vasuki indicus . IITR/VPL/SB 3102-10I-II, vértebras posteriores completas do tronco anterior/médio do tronco em vista anterior ( a ); vista posterior ( b ); vista lateral direita ( c ); vista dorsal ( d ); vista ventral ( e ). IITR/VPL/SB 3102-9I-II, vértebras parciais do tronco médio em vista anterior ( f ); vista posterior ( g ); vista lateral esquerda (invertida) ( h ); vista dorsal ( eu ); vista ventral ( j ). IITR/VPL/SB 3102-4, vértebra média do tronco quase completa em vista anterior ( k ); vista posterior; ( eu ); vista lateral esquerda (invertida) ( m ); vista dorsal ( n ); vista ventral ( o ). IITR/VPL/SB 3102-8I-II, vértebras parciais do tronco médio em vista anterior ( p ); vista posterior ( q ); vista lateral direita ( r ); vista dorsal( s ); vista ventral ( t ). IITR/VPL/SB 3102-11I-III, vértebras parciais do tronco médio em vista posterior ( você ); vista lateral direita ( v ); vista dorsal ( w ); vista ventral ( x ). As setas cinza indicam a direção anterior. Os algarismos romanos nas figuras ( c – e, h – j, r – t, v – w ) referem-se a vértebras individuais em espécimes articulados onde 'I' está voltado para a parte anterior. Setas rosa e brancas indicam fossa e forame na superfície lateral do centro, respectivamente. A seta vermelha indica forame endozigântrico. Pontas de seta brancas indicam protuberância emparelhada na diáfise mediana ventral. cocótilo , côndilo cn , da diapófise, hip hipófise, izr crista interzigapofisária msf , diáfise mediana nc , canal neural nrl , lâmina do arco neural ns , espinha neural pa , parapófise pcof , forame paracotilar pcofo , fossa paracotilar , po pós-zigapófise, pr prezigapófise, psl lâmina prespinal scf , forame subcentral , fossa subcentral scfo, zg zygantrum, zs zigosfeno. A barra de escala representa 50 mm.
Horizonte e localidade
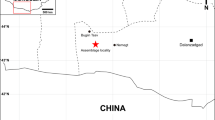
Formação Naredi; Mina de linhita Panandhro, distrito de Kutch, estado de Gujarat, oeste da Índia.
Diagnosis
Vasuki exhibits a unique combination of the following characters: presence of prominent paracotylar foramina (shared with Madtsoiidae); middle-sized cotyle (shared with Madtsoiidae); median prominence on ventral margin of centrum (shared with Madtsoiidae); prezygapophyseal process absent; high angle of synapophysis with horizontal in anterior view (avg. 71.5°); MTV diapophysis level with dorsoventral midpoint of neural canal (shared with Madtsoia madagascariensis, Madtsoia camposi, Wonambi barriei and Adinophis); prezygapophyseal buttress succeeded posteriorly by elliptical fossa (shared with Madtsoia pisdurensis); deep V-shaped embayment (shared with Gigantophis garstini and Madtsoia pisdurensis); oval precloacal cotyle (shared with Gigantophis garstini and Madtsoia pisdurensis); transversely wide vertebrae (shared with Gigantophis garstini and Madtsoia pisdurensis); neural spine posteriorly canted (shared with Gigantophis garstini and Madtsoia pisdurensis); broad hemal keel with posterior process (shared with Gigantophis garstini and Madtsoia pisdurensis); strongly notched anterior zygosphenal margin; endozygantral foramen present (shared with Madtsoia madagascariensis, Powellophis and Gigantophis garstini).
Autapomorfias: vértebras excepcionalmente grandes [comprimento do centro (cL): 37,5–62,7 mm e largura pré-zigapofisária (prW): 62,4–111,4 mm]; seção transversal da coluna neural em forma de pá; quilha hemal pouco desenvolvida que permanece dorsal às parapófises; processo posterior em forma de cinzel da quilha hemal.
Descrição
A coleção compreende 27 vértebras associadas, em sua maioria bem preservadas e incluindo algumas em articulação (Figs. 2 A – T, 3 A – W). 22 das 27 amostras podem ser atribuídas com segurança à região pré-cloacal com base na ausência de hemapófises, pleurapófises e linfapófises, e são ainda restritas a uma posição anterior à região posterior do tronco, como sugerido por uma maior largura mediolateral do arco neural em comparação ao comprimento do centro (sensu LaDuke 1 ; Rio e Mannion 2 ; Tabelas Suplementares 1 , 2 ; Figura Complementar 2 ). Essas dimensões vertebrais são geralmente encontradas em madtsoiídeos de grande porte, como Gigantophis 2 ; Yurlunggur 11 , Madtsoia 1 , 10 , 13 e Wonambi 25 . Além disso, o encerramento das suturas vertebrais sugere que estes espécimes provavelmente atingiram a maturidade esquelética, semelhante, por exemplo, a Madtsoia pisdurensis 8 .
Vasuki é caracterizado por vértebras excepcionalmente grandes, onde o comprimento do centro (cL) e a largura pré-zigapofisária (prW) variam entre 37,5–62,7 e 62,4–111,4 mm, respectivamente (Tabela Suplementar 2 ). Reconhecemos isso como uma autapomorfia, uma vez que essas proporções eclipsam todos os madtsoiídeos de grande porte [ Madtsoia (cL = 18–25 mm; prW = 35–65 mm; LaDuke et al. 1), Gigantophis (cL = 28–41 mm; prW = 44–66 mm; Rio and Mannion2), Platyspondylophis (cL = 18–21 mm; prW = 26–43 mm; Smith et al.21) e Yurlunggur (cL = 15–22 mm; prW = 19–41 mm)]. Alguma cautela, no entanto, é necessária aqui devido às incertezas sobre se o maior tamanho desses madtsoiídeos de grande porte foi capturado, embora o mesmo seja verdade para Vasuki.
Na forma geral, as vértebras do novo táxon indiano são maciças (prW >> cL) e compreendem um centro proceloso. Anteriormente, o centro preserva um cótilo inclinado anteroventralmente, enquanto o côndilo posterior é desviado póstero-dorsalmente, resultando em considerável visibilidade do côndilo e do cótilo nas vistas dorsal e ventral, respectivamente (Fig. 2 C, E). Na vista anterior, o cótilo é fortemente côncavo com sua margem ventral recuada em relação à dorsal. O cótilo é mediolateralmente mais largo do que dorsoventralmente alto (Figs. 2 P, 3 A, F, K; IITR/VPL/SB 3102-4, coW/coH = 1,2; Tabela Suplementar 2 ) como em todos os madtsoiídeos [por exemplo, Gigantophis garstini 2 (NHMUK R8344, vaca/coH = 1,2), Madtsoia madagascariensis (FMNH PR 2551, vaca/coH = 1,24) Yurlunggur (NTM P8695-243, vaca/coH = 1,22) e Wonambi (QMF23038, vaca/coH = 1,4]. Lateralmente, o cótilo é limitado em cada lado por uma fossa paracotilar bem desenvolvida e moderadamente profunda (Figs. 2 K, P, 3 A, K). As margens dorsal e ventral da fossa são proeminentes e definidas por suportes ósseos que emanam de). as margens cotilar dorsolateral e lateral, respectivamente. A margem lateral da fossa, entretanto, está nivelada com a superfície. Além disso, em alguns espécimes a fossa paracotilar é dividida em uma subfossa dorsal mais rasa e ventral mais profunda por uma haste secundária fraca que se estende. lateralmente da margem dorsolateral do cótilo. Um minúsculo forame paracotilar está presente na parte mais dorsal de uma ou ambas as fossas paracotiledonares, imediatamente lateral ao canal neural (Figs. 2 F, K, P, 3 A, K). Embora a presença de fossas e forames paracotiledonares seja uma sinapomorfia de Madtsoiidae 16 , 26 , a morfologia exata dessas características é variável em todo o clado. “ Gigantophis sp.” (CPAG-RANKT-V-1), Menarana nosymena e Adinophis fisaka (FMNH PR 2572) diferem de Vasuki pela presença de forames paracotiledonares pareados em cada lado 1 , 16 , 27 . Em Madtsoia e Eomadtsoia (MPEF-PV 2378) os forames são profundos e comparativamente grandes, enquanto em Yurlunggur estes ocorrem em aglomerados 7 , 8 , 10 , 11 , 13 . Eomadtsoia , no entanto, compartilha com Vasuki a presença de borda ventral proeminente da fossa paracotilar 7 . Em Gigantophis garstini, a fossa paracotilar não possui margem ventral e em Platyspondylophis o forame paracotilar está totalmente ausente. 2 , 21 .
The posterior condyle is transversely wider than high (IITR/VPL/SB 3102–4, cnW/cnH = 1.2; Supplementary Table 2) with the width progressively increasing from ATV (Fig. 2B,G; cnW/cnH = 1.1) to MTV (Fig. 3G, Q; cnW/cnH = 1.2–1.3). Similar proportions of the posterior condyle characterize most madtsoiids [e.g., Nidophis (LPB FGGUB v.547/3, ATV, cnW/cnH = 1.1; LPB FGGUB v.547/1, MTV, cnW/cnH = 1.2); Gigantophis garstini (NHMUK R8344, MTV, cnW/cnH = 1.2 Rio and Mannio2); Madtsoia camposi (DGM 1310b, MTV, cnW/cnH = 1,3] (Fig. 3 G, I, Q). Além disso, na vista posterior, duas pequenas fossas distintas são discerníveis na superfície lateral do centro imediatamente posterior à esquerda diapófise (Fig. 3 G, I, Q). As fossas estão dispostas verticalmente, umas sobre as outras, e separadas por uma crista proeminente. Se essas fossas unilaterais representam uma condição individual ou uma característica geral, não pode ser determinado atualmente e será necessário. espécimes adicionais de Vasuki .
A sinapófise é dorsoventralmente alta e compreende uma diápófise e uma parapófise distintas (Figs. 2 M, R, 3 C, R), ao contrário de Gigantophis garstini , Madtsoia madagascariensis e Madtsoia pisdurensis 1 , 2 , 8 . Na vista anterior, a orientação da sinapófise muda de ventrolateral (Fig. 2 F, K) para um pouco voltada lateralmente (Fig. 3 K, P, U) ao longo da série pré-cloacal. Essa mudança é marcada pelo aumento do ângulo sinapofisário (α), com a horizontal, de ATV (α = média 56,6°) para MTV (α = média 71,5°). Um ângulo sinapofisário mais estreito foi observado na maioria dos táxons madtsoiídeos comparativos, incluindo Eomadtsoia [MPEF-PV 2378 (MTV), α = 45°], Gigantophis garstini [NHMUK R8344 (MTV) α = 48], Madtsoia madagascariensis [FMNH PR 2549 ( ATV), α = 47°; FMNH PR 2551 (MTV), α = 56°], “ Gigantophis sp.” [CPAG-RANKT-V-1 (MTV), α = 56°], Madtsoia camposi [DGM 1310c (MTV), α = 57°], Wonambi [QMF23038 (MTV) α = 58°] e Madtsoia bai [AMNH 3155 (MTV), α = 62°]. Em vista lateral, a sinapófise está inclinada em (β) 20°–27° da vertical em Vasuki . Isto é semelhante a Wonambi [QMF23038, β = 25°], Nanowana [QMF19741, β = ~ 25°], Madtsoia camposi [DGM 1310c, β = 26°] e Yurlunggur [P8695, β = 22°–26°]. Em contraste, ângulos mais amplos caracterizam Gigantophis garstini [NHMUK R8344, β = 30° 2 ], Platyspondylophis [β = 30°–35°], Madtsoia madagascariensis [FMNH PR 2549, β = 33°] e “ Gigantophis sp.” [CPAG-RANKT-V-1, β = ~ 90°], enquanto em Patagniophis [β = 7°–9°], Powellophis [PVL 4714–4, β = 18°] e Madtsoia pisdurensis [225/GSI/PAL /CR/10, β = 12°] os ângulos são mais estreitos.
Um entalhe paracotilar arqueado (sensu LaDuke et al. 1 ), entre a borda cotilar ventral e a parapófise, está consistentemente presente em todos os espécimes (Fig. 2 A, F). A parapófise compreende uma faceta sub-retangular, em vista lateral, e se estende abaixo da borda cotilar ventral no ATV (Fig. 2 F,H,P,R). No MTV, situa-se dorsalmente à borda cotilar ventral (Fig. 3 F, P), ao contrário de Madtsoia pisdurensis e Gigantophis garstini, onde a base parapofisária é ventral e no nível da borda cotilar ventral, respectivamente 2,8. The diapophysis is bulbous and extends laterally beyond the prezygapophysis (Figs. 2F,H, 3P,R), contrary to Powellophis3, Patagoniophis australiensis28, Madtsoia pisdurensis8, Madtsoia madagascariensis1 and Nidophis9. The dorsal margin of the diapophysis remains ventral to the dorsal cotylar margin in ATV (Fig. 2A,F), but becomes level with the dorsoventral midpoint of the neural canal in MTV (Fig. 3K,P). A similar disposition of the MTV diapophysis is observed in Madtsoia madagascariensis, Madtsoia camposi, Wonambi barriei and Adinophis1,2,13,27. The dorsal diapophyseal margin lies between the ventral margin of the neural canal and the dorsoventral midpoint of the cotyle in “Gigantophis sp.”16, Gigantophis garstini2 , Nidófis 9 , Yurlunggur 11 e Powellophis 3 . Em Platyspondylophis, a diapófise estende-se além da margem ventral do canal neural em todas as vértebras pré-cloacais preservadas. 21 .
O contraforte pré-zigapofisário é maciço, não possui um processo pré-zigapofisário e apresenta uma crista oblíqua e romba anteriormente (Fig. 2 F, K). Em vista lateral, o contraforte é sucedido posteriormente por uma fossa elíptica (Fig. 2 C,H,R). A fossa ocorre imediatamente ventral à crista interzigapofisária e medial à diapófise, semelhante a Madtsoia pisdurensis (Mohabey et al. 8 ). As facetas pré-zigapofisárias são elípticas (5022–4, przL/przW = 1,3) e inclinadas ventromedialmente (prα = 20°–28°; Fig. 2 A,D,F,I). Em vista dorsal, essas facetas divergem em 45° do plano sagital, ao contrário das facetas orientadas transversalmente em Madtsoia bai 10 , Madtsoia madagascariensis 1 , Platyspondylophis 21 e Yurlunggur 11 . Pré-zigapófises fortemente divergentes também são observadas em Gigantophis garstini 2 (~ 70°) e Eomadtsoia 7 (60°–80°). As facetas pós-zigapofisárias em Vasuki também são elípticas (IITR/VPL/SB 3102-8II, pozL/pozW = 1,2; Tabela Suplementar 2 ) e orientadas medioventralmente (poα = 12°–26°; Figs. 2 G, J, 3 B, E). A crista interzigapofisária é espessa e direcionada póstero-dorsalmente, atuando como uma ponte entre as pré e pós-zigapofises. Um pequeno forame lateral está presente ventralmente à crista (Fig. 3 L, Q), como em Powellophis 3 . Em vista dorsal, as cristas interzigapofisárias são retas e diferem das cristas arqueadas vistas na maioria dos madtsoiídeos [por exemplo, Madtsoia , Gigantophis garstini , Wonambi , Yurlunggur e Platyspondylophis ] 2 , 8 , 10 , 11 , 13 , 18 , 21 , 28 .
The neural canal is reniform (Figs. 2P,Q, 3F,G) in cross-section and significantly wider than high (ncW/ncH = 3–3.6). It differs from the comparatively narrower and trilobate neural canal in Gigantophis garstini2 (NHMUK R8344, ncW/ncH = 2.3), Platyspondylophis (WIF/A 2271, ncW/ncH = 2.1), Madtsoia (ncW/ncH = 1.3–2.3), Yurlunggur (NTM P8695-243, ncW/ncH = 2.3), “Gigantophis sp.” (CPAG-RANKT-V-1, ncW/ncH = 1.8) and Powellophis (PVL 4714–4, ncW/ncH = 1.6), and the sub-elliptical canal in Wonambi (QMF23038, ncW/ncH = 1.3).
O zigósfeno é trapezoidal e mediolateralmente mais largo que alto (zsW/zsH = 1,4–1,8; Fig. 2 A, K), como em Gigantophis garstini (NHMUK R8344, zsW/zsH = 2 2), Madtsoia bai (AMNH 3155, zsW/zsH = 1.8) and Madtsoia madagascariensis (FMNH PR 2551, zsW/zsH = 1.9). Transversely much wider zygosphenes characterize Nidophis (LPB FGGUB v.547/1, zsW/zsH = 5), Madtsoia camposi (DGM 1310a, zsW/zsH = 2.8), Eomadtsoia (MPEF-PV 2378, zsW/zsH = 2.6), Platyspondylophis (WIF/A 2269, zsW/zsH = 2.2) and Patagoniophis (QMF 19717, zsW/zsH = 5). In Vasuki, the zygosphene is wider than the cotyle, contrary to Gigantophis garstini, “Gigantophis sp.”, Platyspondylophis and Madtsoia1,8,10,13,16,21. Na vista anterior, a margem dorsal do zigósfeno é reta e as facetas articulares são fortemente inclinadas (~ 40° da vertical; Figs. 2 F, P, 3 A). Essas facetas são ovais em vista lateral (IITR/VPL/SB 3102–6, zsfL/zsfW = 1,1). A margem zigofenal anterior é marcadamente entalhada em vista dorsal (zsα = 118°–128°; Figs. 2 I, N, 3 N) e difere do zigofeno não entalhado em Madtsoia pisdurensis 8 , Madtsoia camposi 13 , Eomadtsoia 7 e Platyspondylophis 21 . Em “ Gigantophis sp.” (zsα = 145°) e Madtsoia madagascariensis (zsα = 145°–147°) o zigofeno é fracamente entalhado.
O zigante é mediolateralmente mais largo do que alto, com facetas fortemente inclinadas (50°–60° da horizontal; Fig. 2 B, G, Q). As facetas são elípticas em vista posterior, mas desprovidas de parede mediana presente em Gigantophis garstini 2 . Uma fossa direcionada ântero-ventralmente está presente na base de cada faceta e acomoda um forame endozigantral (Figs. 2 G, 3 B). Este último também está presente em Madtsoia madagascariensis 1 , Powellophis 3 e Gigantophis garstini 2 . Em Vasuki , o teto zygantral acima de cada faceta é médio-dorsalmente convexo e desce como cristas subverticais até o zygantrum (Fig. 2 Q), como em Madtsoia madagascariensis 1 . O teto é ventralmente convexo em Eomadtsoia e Madtsoia pisdurensis , e reto em Powellophis , Platyspondylophis , Yurlunggur e Gigantophis garstini 3 , 7 , 8 , 11 , 21 . Uma grande fossa parazigantral elíptica, orientada dorsolateralmente, flanqueia o zigante lateralmente em ambos os lados e possui um pequeno forame parazigantral (Fig. 2 B, G, Q).
A coluna neural é dorsoventralmente alta (MTV, nsH/tvH = 0,21–0,29, Tabela Suplementar 2 ) e reforçada posteriormente pelas lâminas do arco neural (Fig. 3 B – D, V, W). Estas últimas estendem-se ântero-dorsalmente desde a margem dorsolateral das pós-zigapófises até a margem espinhal dorsal, resultando em um encaixe mediano profundo. Na vista lateral, a coluna é fortemente inclinada póstero-dorsalmente (12°–19° da vertical) com uma margem anterior côncava e uma margem posterior reta. Enquanto uma coluna neural alta caracteriza a maioria dos grandes madtsoiídeos [ Madtsoia camposi (DGM, 1310b, MTV, nsH/tvH = 0,22), Madtsoia madagascariensis (FMNH PR 2551, MTV, nsH/tvH = 0,33), Madtsoia bai (AMNH 3154, MTV, nsH/tvH = 0,22), Wonambi (QMF23038, MTV, nsH/tvH = 0,27)], é mais suavemente inclinado nesses táxons de grande porte [por exemplo, Madtsoia madagascariensis (27°–33°), Wonambi (30°) , Gigantophis garstini (30°)]. Uma margem anterior convexa em Madtsoia madagascariensis, bem como em Powellophis e Nanowana, os distingue ainda mais de Vasuki . Furthermore, the presence of a sharp postspinal lamina (sensu Tschopp29) on the posterior spinal surface and a spade-shaped cross-section of the spine differentiates Vasuki from other madtsoiids (Figs. 2D,S, 3D). In dorsal view, the neural spine base is flanked on either side by a prominent fossa (Fig. 2I,S), as in Madtsoia pisdurensis8 e Madtsoia madagascariensis 1 . As fossas ocorrem imediatamente posteriores ao zigósfeno e são delimitadas ventralmente por suportes ósseos arredondados e fracos que emanam da margem zigosfenal póstero-lateral. Ventralmente a essas hastes, um forame proeminente está presente na superfície dorsal do arco neural posterior ao zigósfeno (Fig. 2 I), semelhante a Madtsoia madagascariensis 1 .
Na vista ventral, o centro é triangular e mais largo nas parapófises. Grandes fossas subcentrais pareadas, mais proeminentes nas vértebras anteriores do tronco (ATV), ocupam a maior parte da superfície ventral do centro (Figs. 2 J, O, 3 E, T). As fossas são delimitadas lateralmente por robustas cristas subcentrais que se estendem posteromedialmente das parapófises até o ponto médio dorsoventral do côndilo. Essas cristas são retas a fracamente convexas na vista ventral e diferem das cristas côncavas em Patagoniophis 28 e Madtsoia madagascariensis 1 . As fossas subcentrais são separadas por uma quilha hemal baixa transversalmente convexa (Figs. 2 T, 3 E, O, T). Este último é largo, ligeiramente elevado e termina anterior à constrição pré-condilar. A quilha hemal não é proeminente, ao contrário da quilha estreita/afiada em “ Gigantophis sp.”, Eomadtsoia , Nidophis , Nanowana e Powellophis 2 , 3 , 7 , 9 , 16 . Em Vasuki , esta quilha permanece dorsal à margem parapofisária ventral (Figs. 2 M, R, 3 M, R, V), ao contrário da quilha hemal de outros madtsoiídeos que desce abaixo da parapófise. Consequentemente, identificamos a disposição da quilha hemal como uma autapomorfia de Vasuki .
A small subcentral foramen is present on either side of the ventral shaft in Vasuki (Fig. 2E,O,T), as in Madtsoia madagascariensis1, Madtsoia camposi13, Nidophis9, and Patagoniophis28. The hypapophysis is paddle-like with sharp lateral margins and extends up to the level of the ventral condylar rim in ATV (Fig. 2G,H,J,L,M,O). The hypapophysis is directed posteroventrally unlike the ventrally directed hypapophysis in Madtsoia madagascariensis1 and Patagoniophis28. Across the precloacals, the hypapophysis progressively reduces in prominence and is replaced by a chisel shaped structure with paired protuberances separated from the ventral condylar rim by a short, sharp ridge in the mid-trunk vertebrae (MTV; Fig. 3J,O,T,X). This chisel shaped structure appears autapomorphic for Vasuki as it differs from the condition in other madtsoiids.
Phylogenetic analysis
A posição de Vasuki dentro de Madtsoiidae foi testada em uma versão modificada da matriz caráter-táxon de Zaher et al. 30(Análise 1; ver seção “ Métodos ” e Nota Complementar 2 ). Foram recuperadas 50 árvores mais parcimoniosas com comprimento de árvore de 1610, índice de consistência (CI) de 0,386 e índice de retenção (IR) de 0,73. As topologias de árvore resultantes são amplamente consistentes com Zaher et al. 30 como Madtsoiidae foi recuperado como um clado distinto dentro da coroa Serpentes (Fig. 4 , Fig. Complementar 3 ). Madtsoiidae, no entanto, foi mal resolvido e não forneceu informações sobre o inter-relacionamento de Vasuki com os outros membros do clado. A má resolução é provavelmente um reflexo da ausência de material craniano na maioria dos madtsoiídeos e uma função da grande matriz onde muito poucos caracteres vertebrais puderam ser pontuados para a maioria dos táxons de madtsoiídeos. Nós, portanto, realizamos uma segunda análise (Análise 2) removendo todas as Serpentes não-madtsoiid e combinando os caracteres cranianos e vertebrais de Zaher et al. 30 e Garberoglio et al. 3 , respectivamente (ver seção “ Métodos ” e Nota Complementar 3 ). O último conjunto de dados foi usado porque o estudo se concentrou nas relações intragrupo madtsoiid. Nossa análise recuperou apenas duas árvores mais parcimoniosas com comprimento de árvore de 191, IC de 0,634 e IR de 0,62. Ambas as árvores (Fig. 5 , Fig. Complementar 4 ) foram em sua maioria bem resolvidas e as topologias resultantes são amplamente consistentes com estudos recentes 2 , 3 , 7 sobre inter-relacionamentos madtsoiid. Madtsoiidae mostra agrupamento baseado em tamanho com táxons de corpo pequeno (<2 m) e médio-grande (> 3 m) recuperados como clados separados (Fig. 5 ). Vasuki está aninhado dentro de um clado distinto (suporte de Bremer = 3) como um táxon irmão do Cretáceo Superior Indiano Madtsoia pisdurensis + Eoceno Superior do Norte da África Gigantophis garstini .
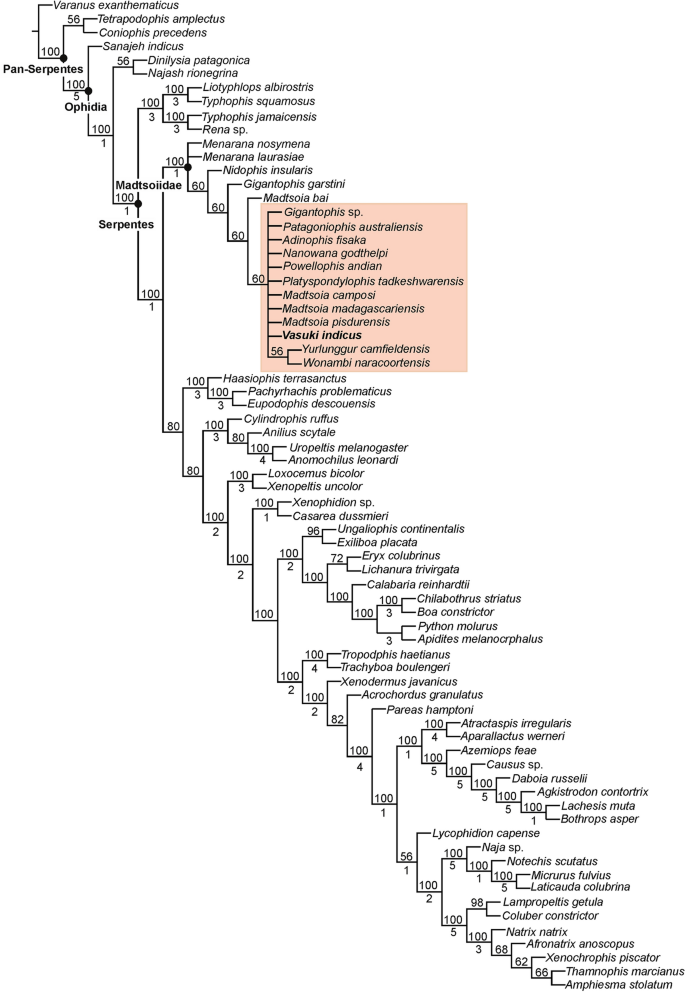
Posição filogenética de Vasuki indicus gen. e sp. novembro. IITR/VPL/SB 3102 na árvore de análise 1 com regra majoritária de 50%. Clado compreendendo Vasuki indicus destacado em rosa. Os números acima e abaixo dos nós indicam a frequência com que um clado é representado nas árvores mais parcimoniosas e nos valores de suporte de Bremer, respectivamente.
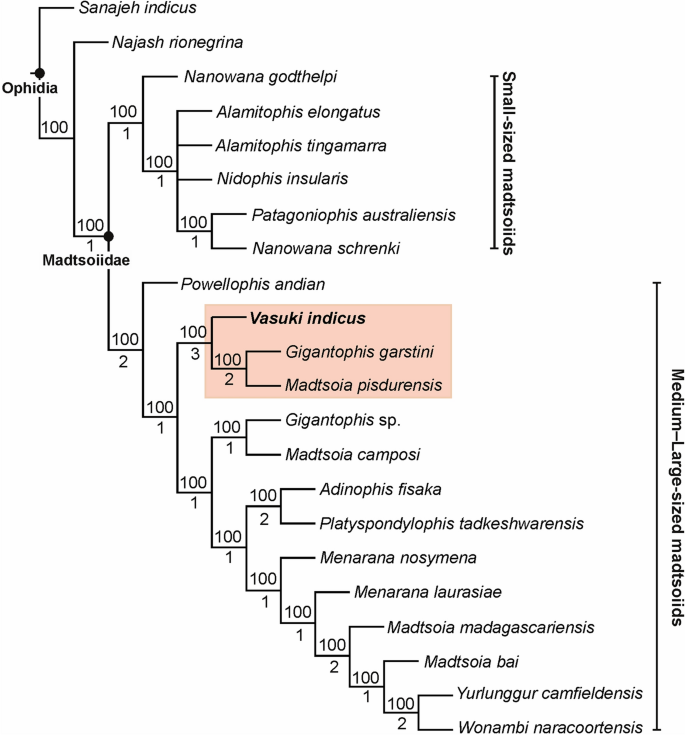
Posição filogenética de Vasuki indicus gen. e sp. novembro. IITR/VPL/SB 3102 na árvore de análise 2 com regra majoritária de 50%. Clado compreendendo Vasuki indicus destacado em rosa. Os números acima e abaixo dos nós indicam a frequência com que um clado é representado nas árvores mais parcimoniosas e nos valores de suporte de Bremer, respectivamente.
Estimation of body length
Estimativas quantitativas do comprimento total do corpo (TBL) de Vasuki foram feitas com base em dois métodos separados que têm sido usados nos últimos anos para estimativa de tamanho de cobras extintas de grande porte (ver seção “ Métodos ” e Tabelas Suplementares 3 – 5 ). Nestes métodos, o TBL foi regredido na largura pós-zigapofisária (seguindo Head et al. 31; Rio and Mannion2) and the prezygapophyseal width (= trans-prezygapophyseal width; following McCartney et al.32, Garberoglio et al.3), respectively. In the present study estimates were made from MTV (IITR/VPL/SB 3102-4, 3102-8I–II, 3102-11II–III), the largest specimens in the collection, following Rio and Mannion2, McCartney et al.32 and Garberoglio et al.3. Both regression models were statistically significant (p < 0.05) and had a high explanatory power (r2 = 0.83–0.96) which asserts their validity. The TBL estimates following Head et al.31 ranges between 10.9 and 12.2 m (Fig. 6A,B), whereas those following McCartney et al.32 is between 14.5 and 15.2 m (Fig. 7A). These estimates, however, should be treated with caution as the collection lacks posterior precloacal and cloacal vertebrae, and an understanding of the intracolumnar variation in madtsoiids is currently non-existent.
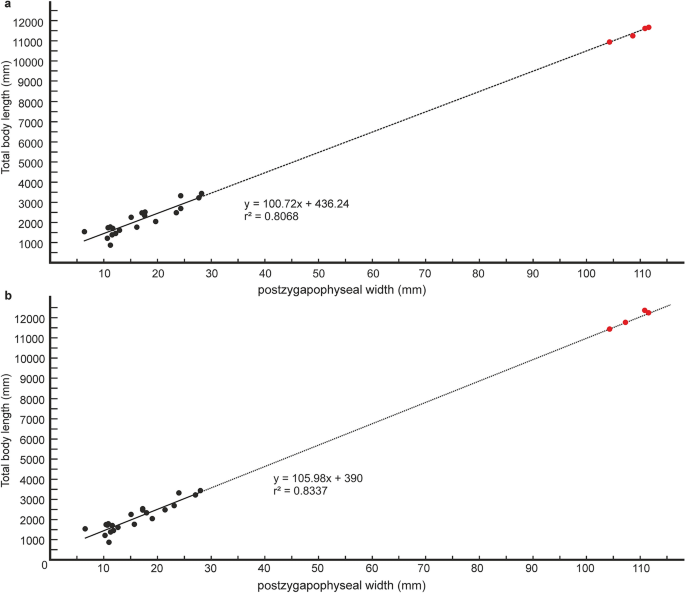
Regressões de métricas vertebrais no comprimento total do corpo em táxons bovinos existentes. Regressão da largura pós-gapofisária no comprimento total do corpo em táxons bovinos existentes das vértebras 60% posteriormente ao longo da coluna vertebral; p = 0,00000003, erro padrão = ± 0,3 m ( a ). Regressão da largura pós-gapofisária no comprimento total do corpo em táxons bovinos existentes das vértebras 65% posteriormente ao longo da coluna vertebral; p = 0,00000001, erro padrão = ± 0,2 m ( b ). Medições de cobras boinas existentes retiradas de Head et al. 31 and plotted as black circles. Estimated body lengths of Vasuki indicus shown in red.
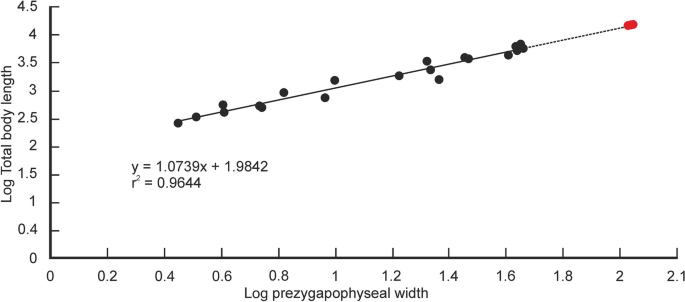
Regressão do comprimento total do corpo na largura pré-zigapofisária em cobras existentes. Medições de cobras existentes retiradas de McCartney et al. 32 e plotados como círculos pretos. Comprimentos corporais estimados de Vasuki indicus mostrados em vermelho. p = 0,000000000000003; erro padrão = ± 0,09m.
It is worth noting that the largest body-length estimates of Vasuki appear to exceed that of Titanoboa, even though the vertebral dimensions of the Indian taxon are slightly smaller than those of Titanoboa. We acknowledge that this observation may be a reflection of the different datasets used to formulate the predictive equations. However, we do not disregard the results based on the dataset of MacCartney et al.32, since the equations derived from the dataset of Head et al.31 involve measurements of extant boine taxa that are taken from vertebrae 60–65% posteriorly along the column. Caution is warranted here because of the uncertainties surrounding the phylogenetic position of Madtsoiidae relative to crown snakes which make estimations based on a model depicting intracolumnar variation in vertebral morphology of a particular extant family/taxa tentative. Consequently, predictive regression equations following McCartney et al.32, which comprise vertebral data from an array of extant snakes, are also considered in our study.
Discussão
Implicações filogenéticas
As análises aqui apresentadas recuperaram um Madtsoiidae monofilético com o clado colocado dentro da coroa Serpentes na Análise 1 (Fig. 4 , Figura Complementar 3 ). Isto está de acordo com a maioria dos estudos filogenéticos que avaliaram a relação do grupo total de cobras dentro de Squamata 30 , 33 , 34 , 35 . Além disso, semelhante a Zaher et al. 30 , a topologia da árvore na Análise 1 recuperou Sanajeh , Diniliysia , Najash em direção ao caule da coroa Serpentes. Embora o clado Madtsoiidae permaneça mal resolvido na Análise 1, encontramos uma combinação de cinco sinapomorfias apoiando a colocação de Vasuki dentro de Madtsoiidae [centro amplo e subtriangular (cap. 613); reentrância profunda em forma de V ao longo da margem posterior do arco neural (cap. 614); presença de forames paracotiledonares bem desenvolvidos (cap. 615); ausência de processo acessório pré-zigapofisário (cap. 616); presença de forame parazygantral (cap. 617)].
Por outro lado, a Análise 2 forneceu insights sobre as relações intragrupais de Madtsoiidae (Fig. 5 , Figura 4 Complementar ). As topologias resultantes são amplamente comparáveis com resultados filogenéticos anteriores 2 , 3 , 7 , 9 , já que descobriu-se que os táxons se dividiam em dois clados baseados no tamanho (grande versus pequeno). Embora a possibilidade de características relacionadas ao tamanho conduzirem tais agrupamentos não possa ser descartada, a recuperação de táxons de pequeno a médio porte (por exemplo, Adinophis , Menarana , Powellophis ) dentro do clado de corpo grande sugere a presença de caracteres independentes de tamanho que apoiam esses clados. Argumento semelhante também foi apresentado por Garberoglio et al. 3ao discutir a ocorrência de clados baseados em tamanho dentro de Madtsoiidae. Contudo, nenhum dos gêneros específicos (por exemplo, Madtsoia , Nanowana , Menarana ) incluídos neste estudo formou clados monofiléticos. O suporte de Bremer para a maioria dos nós internos dentro de Madtsoiidae permanece baixo, embora alguns tenham um suporte comparativamente maior (Fig. 5 ). Estes resultados destacam a necessidade de uma amostragem mais rigorosa envolvendo uma melhor cobertura anatômica dos madtsoiídeos, levando a relações filogenéticas mais robustas.
Uma combinação única de 7 sinapomorfias aninha Vasuki dentro de Madtsoiidae [forame paracotilar bem desenvolvido (cap. 610); proeminência mediana na margem ventral do centro (cap. 611); vaca:dW entre 0,5 e 0,3 (cap. 634); crista lateral nas vértebras pré-cloacais abaixo do forame lateral (cap. 635); zigosfeno espesso (cap. 645); coluna neural moderadamente alta (cap. 648); forames laterais presentes dorsalmente às cristas subcentrais (cap. 650)]. Além disso, uma combinação de 6 sinapomorfias inequívocas [margem posterior do arco neural com embutimento profundo em forma de V (cap. 614); cótilo pré-cloacal oval (cap. 615); vértebras transversalmente largas (cap. 629); quilha hemal não afiada e estreita (cap. 633); coluna neural inclinada posteriormente (cap. 652); presença de processo posterior de quilha hemal (cap. 653)] apoiam a colocação de Vasuki com Gigantophis garstini e Madtsoia pisdurensis . Além disso, uma única autapomorfia caracteriza Vasuki - processo em forma de cinzel de quilha hemal (cap. 654).
Vale ressaltar que algumas das sinapomorfias citadas acima podem ser caracteres individualmente plesiomórficos, é a combinação única de caracteres que justifica a recuperação de Vasuki dentro de Madtsoiidae. Estudos anteriores (por exemplo, Head et al. 31 ; Mohabey et al. 8 ) usaram combinações de caracteres para diagnosticar Madtsoiidae e outros táxons de cobras.
Estimativa do comprimento corporal e paleoecologia
Nossas estimativas de TBL mostram que Vasuki não foi apenas o maior madtsoiid (Tabela 1 ), mas uma das maiores cobras já relatadas. Suas dimensões vertebrais perdem apenas para o Boinae Titanoboa do Paleoceno (Head et al. 31 ). Tentamos inferir a paleoecologia deste grande madtsoiídeo indiano a partir da morfologia vertebral, uma vez que vários estudos anteriores sobre outras cobras extintas (por exemplo, Palaeophis colossaeus , Powellophis e Madtsoia madagascarensis ) destacaram a importância das vértebras nas reconstruções paleoecológicas. 1 , 3 , 32 . As vértebras transversalmente largas de Vasuki apresentam principalmente sinapófises direcionadas lateralmente que teriam sido associadas a costelas direcionadas lateralmente, sugerindo um corpo largo e cilíndrico (ver McCartney et al. 32). These features suggest a non-aquatic lifestyle for Vasuki as opposed to aquatic snakes which may possess high pterapophyses and have laterally compressed vertebrae with ventrally facing synapophyses, thereby placing the ribs beneath the vertebrae3,32,36. A high pterapophysis, however, is absent in many aquatic snakes and changes in the orientation of synapophyses from ventral to lateral across the vertebral column have been previously noted in aquatic snakes such as Simoliophis37. In hydrophiine sea snakes the vertebrae show true lateral compression only in the caudal region. Therefore, the possibility of an aquatic lifestyle for this giant Indian madtsoiid cannot be completely ruled out. An arboreal lifestyle is unlikely, judging from the large size of Vasuki and the fact that arboreal snakes tend to have elongated vertebrae with short zygapophyses38. A non-fossorial habitat is inferred here for Vasuki based on large body-size and non-depressed neural arch-spine complexes which would have placed the dorsal muscles (e.g., M. Semispinalis et spinalis, M. Interarticularis superior) away from the sagittal plane (sensu Auffenburg39)1,3. Isto é ainda apoiado pela presença inferida de M. multifidus espesso dorsoventralmente , que se origina da superfície espinhal neural ântero-dorsal e se insere anteriormente na margem posterior das lâminas do arco neural da vértebra anterior. Semelhança grosseira na morfologia vertebral com pitinídeos de grande porte existentes (por exemplo, Python e Malayopython ) 40 sugere um paleohabitat terrestre/semi-aquático para Vasuki . Evidências corroborativas vêm do ambiente de deposição do horizonte de produção de Vasuki , que foi reconstruído como um pântano de fundo 23 , 41 , 42 , 43 , 44 , 45 , semelhante ao habitat dos grandes pitonídeos modernos.
Vasuki é visto como uma cobra de movimento lento que possivelmente adotou um mecanismo locomotor retilíneo, conforme indicado por seu grande tamanho, vértebras anteroposteriormente curtas e transversalmente largas e ausência de processos pré-zigapofisários acessórios. 1 , 38 , 46 . Uma inferência semelhante, baseada na anatomia, também foi feita para o grande malgaxe Madtsoia madagascariensis 1 , embora a locomoção retilínea também tenha sido documentada em cobras existentes com processos pré-zigapofisários bem desenvolvidos, como víboras 47 . Apesar das incertezas associadas ao mecanismo locomotor de Vasuki , talvez fosse grande demais para ser um forrageador ativo e era mais provavelmente um predador de emboscada que subjugaria suas presas por meio de constrição, semelhante às sucuris modernas e aos pitinídeos de grande porte. 1 , 42 , 48 .
O novo madtsoiid indiano sugere um clima relativamente quente (~ 28 ° C) para a posição paleogeográfica da Índia no Eoceno Médio (início do Lutetiano) dentro da zona tropical 49 , 50 . Esta inferência decorre principalmente da dependência empiricamente derivada da temperatura corporal poiquilotérmica da temperatura ambiente, que por sua vez controla o tamanho máximo do corpo 31 , 32 , 51 . Seguindo Head et al. 31 , a paleotemperatura média anual (MAPT) para o Eoceno Médio foi estimada com base em uma relação entre a temperatura média anual atual (MAT), a diferença TBL entre Vasuki e a píton reticulada ( Malayopython reticulatus , a cobra existente mais longa conhecida) 42 e a taxa metabólica específica de massa das pítons (ver seção “ Métodos ”). O MAPT previsto fica entre 27,2 e 28,6 °C, correspondendo à faixa de temperatura necessária para a sobrevivência de uma cobra de 11 a 15 m, e sugere que os trópicos do Eoceno Médio eram 0,7 a 2,1 °C (ΔT) mais quentes do que atualmente (MAT = 26,5ºC 52 ). Essas estimativas são amplamente comparáveis às do Paleoceno e do Cretáceo Superior com base na extinta Titanoboa (ΔT = 1,9–3,7 °C) e na rã Beelzebufo ampinga (ΔT = 2,1 °C), respectivamente. 52 . Estudos baseados em δ 18 Razões isotópicas de O de foraminíferos e índice TEX86 53 , 54 , 55 , 56 previram altas temperaturas tropicais da superfície do mar (≥ 30 ° C) durante o Eoceno Médio em ~ 47 Ma, enquanto algumas estimativas sugerem resfriamento tropical para o início do Eoceno Médio e Final, mas particularmente durante 45-34 Ma 57 , 58 . A paleotemperatura inferida aqui (<30 °C) é inferior às estimativas acima mencionadas (≥ 30 °C), mas sugere que o clima do Eoceno Médio (início do Lutetiano, ~ 47 Ma) era mais quente do que o atual.
Uma possível limitação deste estudo poderia ser o uso de um pitinídeo ( Malayopython reticulatus ) como análogo moderno, especialmente porque Pythonoidae e Madtsoiidae são filogeneticamente distantes. No entanto, a nossa escolha de um análogo moderno baseia-se no modo de forrageamento inferido e no paleohabitat terrestre/semi-aquático de Vasuki , utilizando dados anatômicos e o ambiente de deposição do horizonte fossilífero. Estes últimos são semelhantes aos dos grandes pitinídeos modernos que habitam pântanos, pântanos e florestas de várzea. 41 , 42 , 43 , 45 .
Na Índia, eventos hipertérmicos do Paleógeno, como PETM e ETM2 estão bem documentados nas bacias de Kutch e Cambay, no oeste da Índia, com base em δ 13 C excursões negativas 23 , 59 , 60 , 61 . Em comparação, os estudos sobre as paleotemperaturas do Paleógeno são escassos. Com base nas razões isotópicas de oxigênio, temperaturas superiores a 30 °C foram determinadas para o Paleoceno tardio e início do Eoceno, enquanto temperaturas mais baixas, variando entre 22 e 28 °C, foram relatadas para o Eoceno Médio-Final (~ 45-37 Ma). 62,63. Our new estimates show that while the paleoclimate during the Middle Eocene (~ 47 Ma) became cooler compared to the Late Paleocene and early Eocene, it was still higher than at present. Further studies on Paleogene climates in the context of squamate speciation and extinction pattern are necessary in view of their suggested correlation with temperature patterns64,65,66.
Paleobiogeography
Madtsoiids eram um grupo importante de cobras terrestres cuja extensão temporal abrange a fronteira do Cretáceo-Paleógeno. As ocorrências fósseis retratam uma distribuição distorcida dessas cobras, já que a maioria dos táxons são conhecidos nas massas terrestres de Gondwana, exceto na Antártida. 1,2 (Figs. 8, 9). The Laurasian record is extremely poor with madtsoiids known only from the Late Cretaceous (upper Campanian–Maastrichtian) of southern Europe9. O padrão de distribuição também mostra o aparecimento de táxons em massas de terra que foram separadas durante o Cretáceo Superior e o Cenozóico, mas que compartilham relações filogenéticas estreitas indicando ligações biogeográficas (sensu LaDuke et al. 1 ). Este enigma é apropriadamente ilustrado pela presença de Madtsoia no Cretáceo Superior (Maastrichtiano) de Madagascar e da Índia e no Paleógeno Inferior da América do Sul, e Menarana no Maastrichtiano de Madagascar e Espanha 1 , 8 (Fig. 8 ). Estudos anteriores apresentaram vários cenários para a paleobiogeografia dos madtsoiídeos, incluindo: uma distribuição pan-Gondwana, embora sem amostragem, durante o Cretáceo Inferior, seguida por extinções regionais e/vicariância; presença de pontes terrestres que permitem a dispersão entre diferentes massas terrestres de Gondwana e para a Europa; dispersão de sorteios entre continentes separados por barreiras oceânicas 1 , 8 , 9 . No entanto, Rio e Mannion 2 argumentou a favor de uma distribuição pan-Gondwanana inicial e de dispersões transtethianas entre a África e a Europa no Cretáceo Superior. O novo madtsoiid indiano do Eoceno Médio aumenta ainda mais a complexidade da biogeografia dos madtsoiid devido aos seus estreitos laços filogenéticos com o Madtsoia pisdurensis do Norte da África do Eoceno Superior do Cretáceo Superior da Índia e o Gigantophis garstini (Figs. 5 , 8 ).
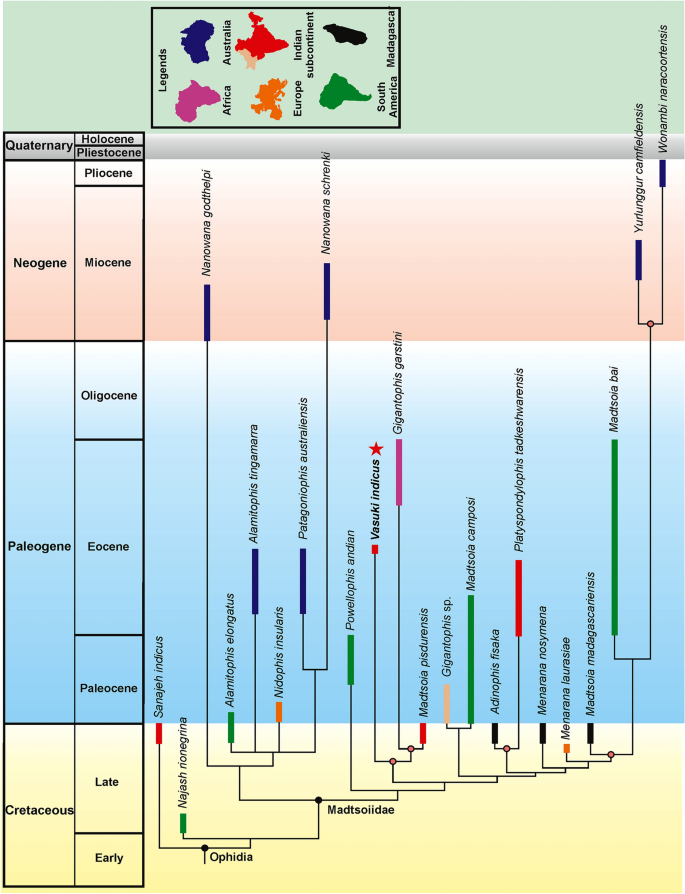
Árvore filogenética calibrada no tempo, com base na árvore de regra majoritária de 50% da Fig . A estrela vermelha indica a posição do Vasuki indicus . Os clados para os quais foram discutidos cenários biogeográficos são marcados com nós coloridos.
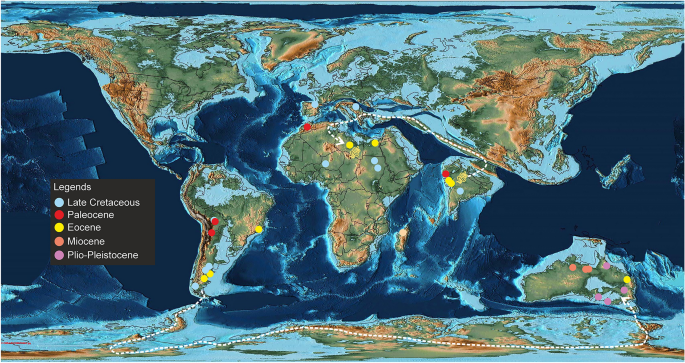
Distribuição paleogeográfica de madtsoiídeos com táxons de diferentes idades plotados juntos em um mapa simplificado do Eoceno Médio (50 Ma) para mostrar suas ocorrências espaço-temporais globais. As linhas tracejadas indicam possíveis rotas de dispersão entre a América do Sul e a Austrália e o subcontinente indiano e o Norte da África. Mapa paleogeográfico após Scotese 43 e proveniente de https://www.earthbyte.org/paleomap-paleoatlas-for-gplates/ [Este trabalho está licenciado sob a Licença Internacional Creative Commons Attribution 4.0. http://creativecommons.org/licenses/by/4.0/ ]. Fonte de informações sobre distribuição de madtsoiid no banco de dados Paleobiologia ( https://www.paleo-biodb.org/ ).
Para avaliar o significado biogeográfico de Vasuki construímos uma árvore filogenética calibrada no tempo, uma vez que esta abordagem tem sido amplamente utilizada em vários estudos anteriores para avaliar o significado paleobiogeográfico de diferentes grupos de vertebrados, incluindo cobras e dinossauros 2 , 67 , 68 , 69 . A lógica por trás desta abordagem é que as relações filogenéticas são amplamente consideradas sugestivas de laços biogeográficos. 1 , 70 , 71 , 72 . A árvore filogenética usada aqui é baseada em dados anatomicamente esparsos porque a maioria dos táxons de madtsoiid são conhecidos exclusivamente a partir de vértebras e carecem de material craniano, resultando em suporte fraco (suporte de Bremer) para a maioria dos nós internos dentro de Madtsoiidae. Por esta razão, restringimos as nossas interpretações biogeográficas apenas aos nós que tinham suporte comparativamente maior (suporte de Bremer ≥ 2; Fig. 8 ). No geral, os cenários paleobiogeográficos aqui apresentados devem ser tratados com cautela, pois futuras descobertas de fósseis podem alterar a posição filogenética de alguns táxons de madtsoiídeos e, por sua vez, as atuais inferências biogeográficas.
Não obstante as limitações acima mencionadas, a árvore resultante em nosso estudo é consistente com o consenso atual sobre as origens dos madtsoiid, pois sugere uma origem Gondwana, refletindo o fato de que todos os táxons divergentes iniciais conhecidos são de antigas massas terrestres de Gondwana (Fig. 8 ). A topologia da árvore defende trocas bióticas entre a América do Sul, Madagascar e Austrália, uma vez que o malgaxe Madtsoia madagascariensis (Cretáceo Superior) e o sul-americano Madtsoia bai (Eoceno) são grupos externos sucessivos ao clado que compreende o Neógeno Yurlunggur e Wonambi da Austrália. As reconstruções paleogeográficas retratam a fragmentação da maioria das principais massas de terra de Gondwana pelo início do Cenomaniano, com o Indo-Madagascar separando-se da Austrália-Antártica em ~ 110 Ma 73 , 74 , 75 . No entanto, estudos anteriores sugeriram que as conexões terrestres entre a América do Sul e a Austrália, facilitando a dispersão da fauna através da Antártica, persistiram até o início do Eoceno. 1 , 75 (Fig. 9 ). Por outro lado, a ligação biótica malgaxe-sul-americana-australiana pode provavelmente ser explicada pela presença de madtsoiídeos ou dos seus ancestrais comuns mais recentes nestes blocos continentais antes da sua dissolução. Estudos recentes sobre a biogeografia de madtsoiid prevêem uma dispersão pan-Gondwana dessas cobras no Cretáceo Inferior, com linhagens fantasmas de árvores calibradas no tempo prevendo uma origem aptiana de Madtsoiidae 1 , 2 , 8 , 9 , 19 . O registro fóssil, no entanto, é inconsistente com as hipotéticas origens dos madtsoiídeos do Cretáceo Inferior, uma vez que seus primeiros representantes atualmente conhecidos são do Coniaciano-Santoniano do Níger. 1 , 2 , 9 , 76 , 77 . A amostragem futura dos horizontes pré-Maastrichtianos da África e do Indo-Madagascar pode ajudar a resolver este enigma.
Os madtsoiídeos indianos, nomeadamente Vasuki indicus , Madtsoia pisdurensis e Platyspondylophis tadkeshwarensis , são divididos em dois subclados distintos (Fig. 8 ). Platyspondylophis (Ypresian) e o malgaxe Adinophis fisaka (Maastrichtiano) são recuperados como táxons-irmãos, enquanto Vasuki (antigo lutetiano) é o primeiro membro divergente de um clado que compreende Madtsoia pisdurensis (Maastrichtiano) e o norte-africano Gigantophis garstini (Priaboniano). Essas relações filogenéticas sugerem trocas bióticas do Cretáceo Superior-Paleógeno entre o subcontinente indiano, Madagascar e o Norte da África. Entre as várias hipóteses concorrentes que explicam tais ligações faunísticas, Krause et al. 74 conexões hipotéticas ( trampolins ) entre o subcontinente indiano, Madagascar e África durante o Cretáceo Superior, que foram possivelmente destruídas em eventos tectônicos subsequentes (por exemplo, subducção, vulcanismo relacionado a hotspots). O arco Omã-Kohistão-Ladakh (OKL) é outra via biogeográfica que se considera ter facilitado os intercâmbios bióticos entre o Norte da África e a Índia após a colisão do subcontinente com OKL em ~ 80 Ma 78. While there is some support from paleomagnetic and radiometric data for the 80 Ma Indo–OKL collision78, subsequent studies based on detrital zircon ages and dating of post-collisional olasses have provided alternate explanations bearing on the sequence of accretion of the OKL with India/Asia79,80. These studies support OKL–Eurasia collision by ~ 100–80 Ma, with India colliding with Asia + OKL only during the Paleogene. This makes the possibility of Late Cretaceous Indo–African faunal exchange less likely2. More recent studies based on paleomagnetic data propose an initial collision between India and Kohistan-Ladakh arc at ~ 60–50 Ma followed by their final collision with Asia at ~ 45–50 Ma, with the arc being positioned at 8.3 ± 5.6°N at ~ 66–62 Ma81,82.
Among the scenarios discussed above we consider the following to be the most plausible explanation for the Indo-Madagascar-North African biotic links suggested by phylogenetic disposition of the Indian madtsoiids:
-
(eu)
Uma relação de táxons irmãos entre o Maastrichtiano Malgaxe Adinophis fisaka e o indiano Platyspondylophis (Ypresian) sugere um evento de dispersão durante ou antes da separação Indo-Madagascar em ~ 88 Ma 83 . A direcção da dispersão, no entanto, permanece incerta, uma vez que a evidência fóssil disponível não permite uma avaliação crítica desta hipótese devido ao fraco registo de amostragem de depósitos malgaxes e indianos pré-Maastrichtianos. No entanto, a recuperação de Madtsoia do Maastrichtiano da Índia e de Madagascar 8 (Fig. 8 ) apoia a prevalência de suas ligações bióticas, como também sugerido por outros grupos, incluindo lagartos cordiliformes e o nigerofídeo Indophis 75 , 84 .
-
(ii)
Após a separação Indo-Madagascar em ~ 88 Ma, houve longos períodos de isolamento que terminaram com a colisão do subcontinente indiano + arco Kohistan-Ladakh com a Ásia no início do Paleógeno 50 , 81 , 82 resultando em caminhos biogeográficos com o Norte da África através do sul da Eurásia (Fig. 9 ).
-
(iii)
Vasuki , Madtsoia pisdurensis e Gigantophis garstini formam um clado distinto, com exclusão de outros, com os primeiros táxons divergentes da Índia (Fig. 8 ). Este clado também mostra ligações filogenéticas estreitas entre os táxons indianos do Cretáceo Superior e do Eoceno Médio, sugerindo uma possível origem indiana para este clado. A colocação de Gigantophis garstini dentro deste clado indica possíveis eventos de dispersão da Índia para o Norte da África após a colisão Índia-Ásia, consistente com o Eoceno Superior (Priaboniano, 37-35 Ma 2 ) age of Gigantophis and recent paleobiogeographic reconstructions showing dispersal routes between India and North Africa via southern Eurasia following the collision43 (Fig. 9). Whereas an African origin of Gigantophis garstini cannot be ruled out considering the recovery of madtsoiids from the Late Cretaceous deposits of that continent, the taxonomic and phylogenetic uncertainties offer little support for this hypothesis. However, Rio and Mannion’s2 A explicação alternativa de que uma dispersão pan-Gondwana do Cretáceo Inferior e longas linhagens fantasmas podem ter levado a relações filogenéticas estreitas entre Gigantophis garstini e os madtsoiídeos indianos, embora potencialmente válida, é atualmente fracamente apoiada devido à má amostragem.
Para resumir, identificamos uma linhagem de madtsoiídeos excepcionalmente grandes (representados pelos maiores madtsoiídeos conhecidos, Vasuki e Gigantophis garstini ) que se originaram no subcontinente indiano e posteriormente se espalharam para a África através do sul da Eurásia durante o Eoceno. A descoberta de Vasuki e a escassa cobertura anatômica dos madtsoiídeos conhecidos destacam a necessidade de uma amostragem rigorosa dos depósitos do Cretáceo Superior e do Paleógeno Gondwana. A recuperação de material adicional e novos táxons (incluindo formas de grande porte) pode fornecer mais informações sobre a sistemática e a biogeografia dos madtsoiídeos.
Métodos
Descrição osteológica
A descrição osteológica dos espécimes esqueléticos foi realizada seguindo a nomenclatura de LaDuke et al. 1 , Rio e Mannion 2 e Mohabey et al. 8 . Diferentes parâmetros dos espécimes fósseis foram medidos (Figura 2 Complementar ) utilizando paquímetros digitais Mitutoyo com precisão de 0,01 mm. Desenhos explicativos são usados sempre que necessário. A terminologia para lâminas vertebrais e fossas segue Rio e Mannion 2 e Tschopp 29 .
Análise filogenética
A afinidade filogenética de Vasuki foi avaliada em duas análises separadas (Análise 1 e 2). Na Análise 1 (Conjunto de Dados Suplementares 1 ), a matriz caráter-táxon de Zaher et al. 30 foi usado. Todos os toxicoferans não-Pan-Serpentes foram removidos, exceto Varanus exanthematicus , que foi usado como grupo externo. 15 táxons madtsoiid, incluindo Vasuki , foram adicionados. A matriz caractere-táxon incluiu 72 táxons e 785 caracteres. A análise filogenética foi realizada no TNT versão 1.6 85 onde a memória do software foi configurada para reter 10.000 árvores e um buffer de exibição de 10 Mb. A opção Pesquisa Tradicional foi utilizada para analisar o conjunto de dados. As restrições para a análise incluíram 50 replicações de árvores de Wagner, nas quais o algoritmo de troca foi a reconexão de bissecção com 10 árvores salvas por replicação. Para determinar a robustez dos nós, os valores de suporte de Bremer foram calculados usando o script bremer.run no qual apenas árvores abaixo do ideal em 20 etapas foram retidas.
In Analysis 2 (Supplementary Dataset 2) all non-madtsoiid Serpentes were removed except for the basal ophidians Najash and Sanajeh. The latter taxon was used as the outgroup. The dataset combined the cranial and vertebral characters of Zaher et al.30 and Garberoglio et al.3, respectively. 3 additional madtsoiid taxa were included. The character-taxon matrix included 22 taxa and 656 characters. The analysis was performed using TNT version 1.685 following the software settings and search parameters of Analysis 1. The script bremer.run was used to calculate Bremer support values in which only trees suboptimal by 20 steps were retained.
Árvore calibrada por tempo
Isto foi construído traçando os intervalos temporais dos táxons de cobras na árvore de regras majoritárias da Análise 2 em relação a uma escala de tempo geológica numericamente calibrada. Os intervalos temporais dos táxons utilizados neste estudo foram obtidos do Banco de Dados de Paleobiologia ( https://www.paleobiodb.org/ ), Rio e Mannion 2 e Garberoglio et al. 3 .
Estimativa do comprimento do corpo
As estimativas do comprimento corporal de Vasuki foram baseadas nos conjuntos de dados de Head et al. 31 e McCartney et al. 32 . O conjunto de dados de Head et al. 31 compreende medidas de largura trans-pós-pós-zigapofisária (poW) e TBL de 21 táxons bovinos existentes, enquanto a de McCartney et al. 32 incluem medidas de largura trans-prezigapofisária e comprimento total do corpo de 21 cobras existentes.
As seguintes equações de regressão preditiva foram formuladas após
-
1.
Cabeça et al. 31 :
where postzygapophyseal width (x) is equated with the total body length (y). The dataset was from vertebrae 60% posteriorly along the vertebral column, and was not log transformed as the measured parameters were approximately normally distributed (sensu Head et al.31).
where postzygapophyseal width (x) is equated with the total body length (y). The dataset was from vertebrae 65% posteriorly along the vertebral column, and was not log transformed as the measured parameters were approximately normally distributed (sensu Head et al.31).
McCartney et al. 32 and Garberoglio et al.3:
onde a largura trans-prezigapofisária (x) é igualada ao comprimento total do corpo (y). Os valores transformados em log dos parâmetros medidos foram usados para normalizar o conjunto de dados.
Em estudos anteriores, os comprimentos corporais foram estimados para cobras extintas, que fazem parte de clados existentes, usando métodos de máxima verossimilhança 31 , 32 . Cabeça et al. 31 desenvolveram um modelo que descreve a variação intracolunar da morfologia vertebral em bovinos existentes para atribuir espécimes vertebrais do boide gigante extinto Titanoboa à sua posição mais provável na coluna vertebral. Com base nos marcos vertebrais, os espécimes de Titanoboa foram combinados em uma posição 60-65% posteriormente ao longo da coluna (MTV, sensu Rio e Mannion 2 ), e as estimativas de tamanho foram obtidas regredindo o TBL no poW com base nas vértebras dos ossos existentes nessas posições. No entanto, tais modelos que mostram variação intracolunar em madtsoiídeos são atualmente inexistentes, pois muito poucas dessas cobras são conhecidas da coluna vertebral completa/quase completa. 2 . As estimativas de tamanho de Vasuki foram calculadas neste estudo usando MTV seguindo Rio e Mannion 2 , embora essas estimativas devam ser consideradas provisórias, pois os espécimes de Vasuki não podem ser atribuídos à mesma posição que as vértebras boinas usadas para formular as equações. Além disso, pode haver diferenças na relação entre poW e TBL entre boines existentes e Vasuki . Além disso, as incertezas associadas à posição filogenética de Madtsoiidae em relação às cobras-coroa impedem a formulação de modelos que mostram variação intracolunar na morfologia vertebral com base em qualquer clado existente. Consequentemente, equações de regressão preditiva, baseadas em dados de uma série de cobras existentes de McCartney et al. 32 , foram usados para determinar o comprimento do corpo do novo táxon indiano e, portanto, os comprimentos estimados, embora razoáveis, também devem ser tratados com cautela.
Estimativa da paleotemperatura
As estimativas da paleotemperatura foram obtidas usando a seguinte equação fornecida em Head et al. 31 :
onde MAPT é a paleotemperatura média anual; MAT é a temperatura média anual atual (26,5 °C 52 ); TBL M = 10,05 m é o comprimento total máximo do corpo de Malayopython reticulatus 41 ; TBL V é o comprimento corporal máximo estimado de Vasuki (15,2 m); Q 10 (taxa metabólica específica de massa de pitinídeos) = 2,6 86 ; α (componente de escala metabólica) = 0,17 52 , 87 .
Como Madtsoiidae é um clado extinto, o comprimento do corpo de Malayopython reticulatus (Serpentes, Pythonidae) foi usado no estudo, pois é a cobra existente mais longa conhecida 42 . A escolha do Malayopython como o análogo moderno é baseada na semelhança na morfologia vertebral grosseira e no modo de vida e habitat inferido entre Vasuki e os pythonídeos de grande corpo existentes 40 , 41 , 42 , 43 , 45 . No entanto, na ausência de representantes existentes de madtsoiídeos ou de seus parentes próximos, os valores estimados da paleotemperatura devem ser tratados com cautela.
Disponibilidade de dados
Todos os dados associados ao manuscrito são fornecidos no Arquivo Suplementar.
Code availability
Nomenclatural acts. This published work and the nomenclatural acts it contains have been registered in Zoo- Bank, the proposed online registration system for the International Code of Zoological Nomenclature (ICZN). The LSIDs for this publication are urn:lsid:zoobank.org:act: 2F44E9BE-AE99-45E8-A132-D36A935D3B36 (Vasuki) and urn:lsid:zoobank.org:act: 0DD3FB9F-A500-4FFE-842C-EFE51EC76E4D (V. indicus).
Abbreviations
- AMNH:
-
American Museum of Natural History, New York
- CPAG:
-
Centre of Pure and Applied Geology, University of Sindh, Pakistan
- DGM:
-
Departamento Nacional de Produção Mineral, Rio de Janeiro, Brazil
- FMNH:
-
The Field Museum, Chicago, USA
- IITR/VPL/SB:
-
Laboratório de Paleontologia de Vertebrados, Instituto Indiano de Tecnologia Roorkee, Roorkee, Índia
- LPB FGGUB:
-
Laboratório de Paleontologia, Faculdade de Geologia e Geofísica, Universidade de Bucareste, Bucareste, Romênia
- MPEF-PV:
-
Coleção Paleontológica de Vertebrados, MuseoPaleontológico Egidio Feruglio, Trelew, Província de Chubut, Argentina
- NTM:
-
Museu do Território do Norte, Austrália
- NHMUK:
-
Museu de História Natural, Londres, Reino Unido
- QMF:
-
Museu de Queensland, Brisbane, Austrália
- PVL:
-
Coleção Paleontológica de Vertebrados do Instituto Miguel Lillo, San Miguel de Tucumán, Argentina
- WIF/A:
-
Instituto Wadia de Geologia do Himalaia, Dehradun, Índia
Referências
Laduke, T. C., Krause, D. W., Scanlon, J. D. & Kley, N. J. A late cretaceous (Maastrichtian) snake assemblage from the Maevarano formation, Mahajanga basin, Madagascar. J. Vertebr. Paleontol. 30, 109–138 (2010).
Rio, J. P. & Mannion, P. D. The osteology of the giant snake Gigantophis garstini from the upper Eocene of North Africa and its bearing on the phylogenetic relationships and biogeography of Madtsoiidae. J. Vertebr. Paleontol. 37, e1347179 (2017).
Garberoglio, FF, Triviño, LN & Albino, A. Uma nova cobra madtsoiida do Paleógeno da América do Sul (noroeste da Argentina), baseada em um esqueleto pós-craniano articulado. J. Vertebr. Paleontol. 42 , e2128687 (2022).
Albino, AM Simpósio sobre a Evolução dos Vertebrados Mesozóicos. Em Anais IV Congresso Argentino de Paleontologia e Bioestratigrafia (ed. Bonaparte, JF) (Formação Los Alamitos, 1986).
Albino, AM Uma nova cobra (Reptilia) no Cretáceo Superior da Patagônia, Argentina. Pesquisas 21 , 58–63 (1994).
Rage, JC, Prasad, GVR & Bajpai, S. Cobras adicionais do Cretáceo Superior (Maastrichtiano) da Índia. Cretaco. Res. 25 , 425–434 (2004).
Gómez, RO, Garberoglio, FF & Rougier, GW Uma nova cobra do Cretáceo Superior da Patagônia: Filogenia e tendências na evolução do tamanho corporal de cobras madtsoiid. CR Palevol. 18 , 771–781 (2019).
Mohabey, DM, Head, JJ & Wilson, JA Uma nova espécie de cobra Madtsoia do Cretáceo Superior da Índia e suas implicações paleobiogeográficas. J. Vertebr. Paleontol. 31 , 588–595 (2011).
Vasile, Ş, Csiki-Sava, Z. & Venczel, M. Uma nova cobra madtsoiid do Cretáceo Superior da Bacia Haţeg, oeste da Romênia. J. Vertebr. Paleontol. 33 , 1100–1119 (2013).
Simpson, GG Um novo fóssil de cobra dos leitos de Notostylops da Patagônia. Touro. Sou. Mus. Nat. História. 67 , 1–22 (1933).
Scanlon, JD Uma nova grande cobra madtsoiid do Mioceno do Território do Norte. O Beagle 9 , 49–60 (1992).
Scanlon, JD Nanowana gen. nov., pequenas cobras madtsoiid do Mioceno de Riversleigh: espécies simpátricas com dentição divergentemente especializada. Mem. Qd Mus. 41 , 393–412 (1997).
Rage, J.-C. Fossil snakes from the Palaeocene of São José de Itaboraí, Brazil. Part I. Madtsoiidae, Aniliidae. Palaeovertebrata 27 , 109–144 (1998).
Rage, J.-C. e outros. Cobras do início do Eoceno de Kutch, Índia Ocidental, com uma revisão dos Palaeophiidae. Geodiversitas 25 , 695–716 (2003).
Rage, J.-C. e outros. Uma fauna diversificada de cobras do início do Eoceno da Mina Vastan Lignite, Gujarat, Índia. Acta Paleontol. Pol. 53 , 391–403 (2008).
Rage, J.-C. et al. First report of the giant snake Gigantophis (Madtsoiidae) from the Paleocene of Pakistan: Paleobiogeographic implications. Geobios 47, 147–153 (2014).
Scanlon, JD & Lee, MSY A serpente Wonambi do Pleistoceno e a evolução inicial das cobras. Natureza 403 , 416–420 (2000).
Wilson, JA, Mohabey, DM, Peters, SE & Head, JJ Predação de dinossauros filhotes por uma nova cobra do Cretáceo Superior da Índia. PLoS Biol. 8 , e1000322 (2010).
Longrich, NR, Bhullar, BAS & Gauthier, JA Uma cobra de transição do período Cretáceo Superior da América do Norte. Natureza 488 , 205–208 (2012).
Martill, DM, Tischlinger, H. & Longrich, NR Uma cobra quadrúpede do Cretáceo Inferior de Gondwana. Ciência 349 , 416–419 (2015).
Smith, T. et al. Uma nova assembleia de vertebrados do início do Eoceno do oeste da Índia revela uma fauna mista de afinidades europeias e gondwanas. Geociências. Frente. 7 , 969–1001 (2016).
Wazir, WA et al. Uma descoberta no Ladakh Himalaia revela a sobrevivência de cobras madtsoiid (Serpentes, Madtsoiidae) na Índia até o final do Oligoceno. J. Vertebr. Paleontol. 41 , e20584012021 (2021).
Agrawal, S. et al. Depósitos de linhita da Bacia de Kutch, oeste da Índia: assinaturas isotópicas e palinológicas de carbono do evento hipertérmico ETM2 do início do Eoceno. J. Ciência da Terra Asiática. 146 , 296–303 (2017).
Westerhold, T., Röhl, U., Donner, B. & Zachos, JC Extensão global dos eventos hipertérmicos do início do Eoceno: Um novo registro de isótopos foraminíferos bentônicos do Pacífico de Shatsky Rise (ODP Site 1209). Paleoceanogr. Paleoclimatol. 33 , 626–642 (2018).
Smith, MJ Pequenos vertebrados fósseis de Victoria Cave, Naracoorte, Sul da Austrália IV. Répteis. Trad. R.Soc. S. Aust. 100 , 39–51 (1976).
Scanlon, JD Crânio da grande cobra não macrostomatana Yurlunggur do Oligo-Mioceno australiano. Natureza 439 , 839–842 (2006).
Pritchard, A. C., McCartney, J. A., Krause, D. W. & Kley, N. J. New snakes from the Upper Cretaceous (Maastrichtian) Maevarano Formation, Mahajanga Basin, Madagascar. J. Vertebr. Paleontol. 34, 1080–1093 (2014).
Scanlon, J. D. Australia’s oldest known snakes: Patagoniophis, Alamitophis, and cf. Madtsoia (Squamata: Madtsoiidae) from the Eocene of Queensland. Mem. Queensl. Mus. 51, 215–235 (2005).
Tschopp, E. Nomenclatura de lâminas vertebrais em lagartos, com comentários sobre variação ontogenética e serial em Lacertini (Squamata, Lacertidae). PLoS ONE 11 , e0149445 (2016).
Zaher, H., Mohabey, DM, Grazziotin, FG e Wilson Mantilla, JA O crânio de Sanajeh indicus , uma cobra do Cretáceo com uma barra temporal superior e a origem da alimentação ofídica de boca larga. Zoológico. J. Linn. Soc. 197 , 656–697 (2023).
Cabeça, JJ et al. A cobra boid gigante dos neotrópicos do Paleoceno revela temperaturas equatoriais passadas mais quentes. Natureza 457 , 715–717 (2009).
McCartney, JA, Roberts, EM, Tapanila, L. & Oleary, MA Grandes cobras paleofiídeos e nigerofiídeos dos depósitos do Paleógeno Trans-Saharan Seaway do Mali. Acta Paleontol. Pol. 63 , 207–220 (2018).
Zaher, H. & Scanferla, CA O crânio da cobra do Cretáceo Superior Dinilysia patagonica Smith-Woodward, 1901, e sua posição filogenética revisitada. Zoológico. J. Linn. Soc. 164 , 194–238 (2012).
Hsiang, AY et al. A origem das cobras: revelando a ecologia, o comportamento e a história evolutiva das primeiras cobras usando genômica, fenômica e registro fóssil. BMC Evol. Biol. 15 , 1–22 (2015).
Harrington, SM & Reeder, TW Inferência filogenética e datação por divergência de cobras usando moléculas, morfologia e fósseis: Novos insights sobre a evolução convergente da morfologia alimentar e redução de membros. Biol. J. Linn. Soc. 121 , 379–394 (2017).
Georgalis, G. L. First potential occurrence of the large aquatic snake Pterosphenus (Serpentes, Palaeophiidae) from Nigeria, with further documentation of Pterosphenus schweinfurthi from Egypt. Alcheringa 1, 1–9 (2023).
Rage, J. C., Vullo, R. & Néraudeau, D. The mid-Cretaceous snake Simoliophis rochebrunei Sauvage, 1880 (Squamata: Ophidia) from its type area (Charentes, southwestern France): Redescription, distribution, and palaeoecology. Cretac. Res. 58, 234–253 (2016).
Johnson, RG O significado adaptativo e filogenético da forma vertebral em cobras. Evolução 9 , 367–388 (1955).
Auffenberg, W. Observações adicionais sobre a evolução da musculatura do tronco em cobras. Sou. Médio. Nat. 65 , 1–16 (1961).
Szyndlar, Z. & Georgalis, GL Um atlas ilustrado da morfologia vertebral de cobras não cenofídicas existentes, com ênfase especial nas porções cloacal e caudal da coluna. Vértebra. Zoológico. 73 , 717–886 (2023).
Murphy, JC & Henderson, RW Contos de cobras gigantes: uma história natural de sucuris e pítons (Krieger, 1997).
Auliya, M., Mausfeld, P., Schmitz, A. & Böhme, W. Revisão da píton reticulada ( Python reticulatus Schneider, 1801) com a descrição de novas subespécies da Indonésia. Naturwissenschaften 89 , 201–213 (2002).
Reed, RN & Rodda, GH Giant Constrictors: Perfis biológicos e de manejo e uma avaliação de risco de estabelecimento para nove grandes espécies de pítons, sucuris e jibóias (Departamento do Interior dos EUA, US Geological Survey, 2009).
Babar, M. E. et al. Biology, habitat and conservation of Indian Rock Python—A brief review. J. Anim. Plant Sci. 29, 349–352 (2019).
Burger, R. Ecologia da Python Reticulada (Malayopython reticulatus): Vida em uma Paisagem Alterada 212. Dissertação de doutorado, Universidade de Cardiff (2022).
Mosauer, W. Sobre a locomoção das cobras. Ciência 76 , 583–585 (1932).
Marvi, H., Bridges, J. & Hu, DL Cobras imitam minhocas: Propulsão usando ondas viajantes retilíneas. JR Soc. Interface 10 , 20130188 (2013).
Rivas, J. A História de Vida da Anaconda Verde (Eunectes murinus), com Ênfase em Sua Biologia Reprodutiva . Dissertação, Univ. Tenessi (2000).
Shukla, A. et al. Temperaturas terrestres equatoriais frias e monções do sul da Ásia no início do Eoceno: evidências da mina Gurha, Rajastão, Índia. Paleogeogr. Paleoclimatol. Paleoecol. 412 , 187–198 (2014).
Scotese, C. R. PALEOMAP PaleoAtlas for GPlates and the PaleoData Plotter Program, PALEOMAP Project. http://www.earthbyte.org/paleomap--‐paleoatlas--‐for--‐gplates/, https://doi.org/10.13140/RG2.2.34367.00166 (2016).
Makarieva, A. M., Gorshkov, V. G. & Li, B.-L. Gigantism, temperature and metabolic rate in terrestrial poikilotherms. Proc. R. Soc. Lond. B 272, 2325–2328 (2005).
Makarieva, AM, Gorshkov, VG & Li, BL Recalibrando o paleotermômetro de cobra. Natureza 460 , E2–E3 (2009).
Pearson, PN et al. Temperaturas quentes da superfície do mar tropical nas épocas do Cretáceo Superior e Eoceno. Natureza 413 , 481–487 (2001).
Pearson, PN et al. Clima tropical quente estável durante a Época Eocena. Geologia 35 , 211–214 (2007).
Cramwinckel, MJ et al. Evolução síncrona da temperatura tropical e polar no Eoceno. Natureza 559 , 382–386 (2018).
Evans, D. et al. Clima de estufa do Eoceno revelado por termometria de isótopos aglomerados acoplados-Mg/Ca. Processo. Nacional. Acad. Ciência. EUA 115 , 1174–1179 (2018).
Inglis, GN et al. Descida em direção à Casa de Gelo: resfriamento da superfície do mar no Eoceno inferido a partir de distribuições GDGT. Paleoceanografia 30 , 1000–1020 (2015).
Zachos, J. et al. Tendências, ritmos e aberrações no clima global de 65 milhões de anos até o presente. Ciência 292 , 686–693 (2001).
Clementz, M. et al. Early Eocene warming events and the timing of terrestrial faunal exchange between India and Asia. Geology 39, 15–18 (2011).
Samanta, A. et al. Do the large carbon isotopic excursions in terrestrial organic matter across Paleocene–Eocene boundary in India indicate intensification of tropical precipitation? Palaeogeogr. Palaeoclimatol. Palaeoecol. 387, 91–103 (2013).
Mitra, A., Dutta, R. & Halder, K. Um estudo sobre moluscos bentônicos e isótopos estáveis de Kutch, oeste da Índia, revela hipertérmicas do início do Eoceno e transgressão pronunciada durante eventos ETM2 e H2. Suíço J. Paleontol. 141 , 1–22 (2022).
Saraswati, PK, Ramesh, R. & Navada, SV Temperaturas isotópicas Palaeogene do oeste da Índia. Lethaia 26 , 89–98 (1993).
Khanolkar, S. & Saraswati, PK Biofácies foraminíferas do Eoceno na Bacia de Kutch (Índia) no contexto de paleoclima e paleoecologia. J. Paleogeogr. 8 , 1–16 (2019).
Prothero, DR As extinções do final do Eoceno-oligoceno. Anu. Rev. Planeta Terra. Ciência. 22 , 145–165 (1994).
Rage, J. C. Mesozoic and Cenozoic squamates of Europe. Paleobiodivers. Paleoenviron. 93, 517–534 (2013).
Cyriac, V. P. & Kodandaramaiah, U. Paleoclimate determines diversification patterns in the fossorial snake family Uropeltidae Cuvier, 1829. Mol. Phylogenet. Evol. 116, 97–107 (2017).
Delcourt, R. & Grillo, O. N. Tyrannosauroids from the Southern Hemisphere: Implications for biogeography, evolution, and taxonomy. Palaeogeogr. Palaeoclimatol. Palaeoecol. 511, 379–387 (2018).
Whitlock, JA e Mantilla, JAW O dinossauro saurópode do Jurássico Superior ' Morosaurus' agilis Marsh, 1889 reexaminado e reinterpretado como um dicraeossaurídeo. J. Vertebr. Paleontol. 40 , e1780600 (2020).
Bajpai, S. et al. Fósseis do dinossauro diplodocóide mais antigo sugerem que a Índia era um importante centro de radiação neossauropode. Ciência. Rep. 13 , 12680 (2023).
Krause, DW, Sampson, SD, Carrano, MT & O'Connor, PM Visão geral da história da descoberta, taxonomia, filogenia e biogeografia de Majungasaurus crenatissimus (Theropoda: Abelisauridae) do Cretáceo Superior de Madagascar. J. Vertebr. Paleontol. 27 , 1–20 (2007).
Gallina, PA, Apesteguia, S., Haluza, A. & Canale, JI Um saurópode diplodocídeo sobrevivente do Cretáceo Inferior da América do Sul. PLoS ONE 9 , e97128 (2014).
Mohabey, DM, Samant, B., Vélez-Rosado, KI & Wilson Mantilla, JA Uma revisão de dinossauros terópodes de corpo pequeno do Cretáceo Superior da Índia, com descrição de novos restos cranianos de um noasaurídeo (Theropoda: Abelisauria). J. Vertebr. Paleontol. 43 , e2288088 (2023).
Ali, J. R. & Krause, D. W. Late Cretaceous bioconnections between Indo-Madagascar and Antarctica: Refutation of the Gunnerus Ridge causeway hypothesis. J. Biogeogr. 38, 1855–1872 (2011).
Krause, DW et al. A história biogeográfica mesozóica dos vertebrados terrestres de Gondwana: percepções do registro fóssil de Madagascar. Anu. Rev. Planeta Terra. Ciência. 47 , 519–553 (2019).
Wilf, P. et al. Esplêndido e raramente isolado: A paleobiogeografia da Patagônia. Anu. Rev. Planeta Terra. Ciência. 41 , 561–603 (2013).
Head, JJ, Howard, AF & Muller, J. A origem e a história evolutiva inicial das cobras (Cambridge University Press, 2022).
Rage, J.-C. Os continentes periatlânticos no Cretáceo Superior: migrações de faunas continentais e problemas paleogeográficos. Cretaco. Res. 2 , 65–84 (1981).
Chatterjee, S., Scotese, CR & Bajpai, S. A inquieta placa indiana e sua viagem épica de Gondwana à Ásia: sua evolução tectônica, paleoclimática e paleobiogeográfica. Geol. Soc. Sou. 529 , 1–147 (2017).
Borneman, NL et al. Idade e estrutura da sutura Shyok na região de Ladakh, no noroeste da Índia: implicações para o deslizamento no sistema de falhas de Karakoram. Tectônica 34 , 2011–2033 (2015).
Andjić, G., Zhou, R., Jonell, TN & Aitchison, JC Um único arco Dras – Kohistan – Ladakh revelado por registros vulcaniclásticos. Geoquímica. Geofísica. Geossist. 23 , e2021GC010042 (2022).
Martin, CR et al. A latitude paleocena do arco Kohistan-Ladakh indica colisão em vários estágios Índia-Eurásia. Processo. Nacional. Acad. Ciência. EUA 117 , 29487–29494 (2020).
Jin, S. et al. Uma grande Índia menor e uma colisão do Eoceno Médio-Início com a Ásia. Geofísica. Res. Vamos. 50 , e2022GL101372 (2023).
Reeves, C. A posição de Madagascar dentro de Gondwana e seus movimentos durante a dispersão de Gondwana. J. Afr. Ciências da Terra. 94 , 45–57 (2014).
Yadav, R., Bajpai, S., Maurya, A. S. & Čerňanský, A. The first potential cordyliform (Squamata, Scincoidea) from India (uppermost Cretaceous–lowermost Paleocene): An African lizard clade brings possible implications for Indo-Madagascar biogeographic links. Cretac. Res. 150, 105606 (2023).
Goloboff, P. A. & Morales, M. E. TNT version 1.6, with a graphical interface for MacOS and Linux, including new routines in parallel. Cladistics 39, 144–153 (2023).
Bedford, GS & Christian, KA Taxa metabólica padrão e temperaturas corporais preferidas em algumas pítons australianas. Austrália. J. Zool. 46 , 317–328 (1998).
Chappell, MA & Ellis, TM Taxas metabólicas de repouso em cobras boid: relações alométricas e efeitos de temperatura. J. Comp. Fisiol. B 157 , 227–235 (1987).
Reconhecimentos
Os autores agradecem os comentários úteis, sugestões e uma crítica construtiva do manuscrito pelos revisores e pelo Editor, Scientific Reports. Os autores também agradecem a Ritu Sharma, Debasis Das, Vivesh Vir Kapur, N. Saravanan, Lisa Cooper, Lauren Stevens e Hans Thewissen pela ajuda durante o trabalho de campo e a Aatreyee Saha, Abhay Rautela e Poonam Verma pela ajuda e discussões. O Conselho de Pesquisa em Ciência e Engenharia (SERB) (Bolsa nº PDF/2021/00468 como Bolsa Nacional de Pós-Doutorado para DD) e o Departamento de Ciência e Tecnologia (Projeto nº SR/S4/ES-222/2006 para SB) , O governo da Índia é reconhecido pelo apoio financeiro. DD gostaria de agradecer ao IIT Roorkee por fornecer instalações de infraestrutura. SB gostaria de agradecer o apoio obtido do IIT Roorkee como parte de sua cátedra no Instituto.
Informação sobre o autor
Autores e Afiliações
Contribuições
SB e DD conceberam o problema. SB coletou os fósseis. DD e SB analisaram e interpretaram os dados e redigiram o manuscrito. SB e DD estiveram envolvidos em revisões posteriores.
Autores correspondentes
Declarações éticas
Interesses competitivos
Os autores declaram não haver interesses conflitantes.
Informações adicionais
Nota do editor
A Springer Nature permanece neutra em relação a reivindicações jurisdicionais em mapas publicados e afiliações institucionais.
Direitos e permissões
Acesso aberto Este artigo está licenciado sob uma Licença Creative Commons Atribuição 4.0 Internacional, que permite o uso, compartilhamento, adaptação, distribuição e reprodução em qualquer meio ou formato, desde que você dê o devido crédito ao(s) autor(es) original(is) e à fonte, forneça um link para a licença Creative Commons e indique se foram feitas alterações. As imagens ou outros materiais de terceiros neste artigo estão incluídos na licença Creative Commons do artigo, salvo indicação em contrário na linha de crédito do material. Se o material não estiver incluído na licença Creative Commons do artigo e o uso pretendido não for permitido por regulamentação legal ou exceder o uso permitido, você precisará obter permissão diretamente do detentor dos direitos autorais. Para visualizar uma cópia desta licença, visite http://creativecommons.org/licenses/by/4.0/ .
Sobre este artigo

Cite este artigo
Datta, D., Bajpai, S. A maior cobra madtsoiid conhecida do período quente do Eoceno da Índia sugere dispersão intercontinental de Gondwana. Sci Rep 14 , 8054 (2024). https://doi.org/10.1038/s41598-024-58377-0
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.