Morfologia dentária no Homo habilis e suas implicações para a evolução do Homo inicial
Nature Communications volume 15 , número do artigo: 286 ( 2024 )
Abstrato
A posição filogenética do Homo habilis é central nos debates sobre a origem e evolução inicial do gênero Homo . Uma grande parte do hipodigma da espécie consiste em restos dentários, mas eles só foram estudados na superfície do esmalte, frequentemente desgastada. Investigamos a morfologia da H. habilis junção esmalte-dentina (EDJ) do , que é preservada em casos de desgaste dentário moderado e conhecida por carregar um forte sinal taxonômico. A morfometria geométrica é usada para caracterizar a forma e o tamanho da coroa dentinária em toda a fileira dentária inferior e maxilar, em comparação com uma ampla amostra comparativa ( n = 712). Descobrimos que a morfologia do EDJ no H. habilis é, em sua maior parte, notavelmente primitiva, apoiando a hipótese de que o hipodigma do H. habilis tem mais em comum com o Australopithecus posterior do que com o Homo . Além disso, o espécime cronologicamente mais jovem OH 16 exibe um conjunto de características derivadas; sua inclusão no H. habilis leva a níveis excessivos de variação.
Conteúdo semelhante sendo visualizado por outras pessoas


Introdução
As origens do gênero Homo permanecem indefinidas. Há mais de 60 anos, Leakey et al. 1 propôs a espécie Homo habilis com a descoberta de fósseis no desfiladeiro de Olduvai, na Tanzânia; propuseram que a espécie ocupava uma lacuna morfológica entre o Australopithecus e o Homo erectus , e a colocaram na base do gênero Homo . Embora a nomenclatura desta nova espécie tenha sido inicialmente controversa, as descobertas subsequentes dos primeiros espécimes de Homo em Olduvai, bem como em outros locais no Quênia, na África do Sul e na Etiópia, levaram gradualmente à aceitação geral de H. habilis como uma espécie válida. 2,3,4,5,6,7. Despite this acceptance, a number of key questions about early Homo remain unresolved, and in particular, we have a poor understanding of the phylogenetic relationship between H. habilis and other species. Additionally, recent work has emphasised hominin species diversity in the Pliocene and Pleistocene, including the co-existence of multiple species in close proximity8,9,10,11,12, and thus a once influential account of hominin evolution, according to which Australopithecus evolved into H. habilis, then H. erectus, and ultimately modern humans13,14, is now considered overly simplistic15,16. In the case of the early Homo fossil record; a number of studies have suggested that the variation exceeds that expected of a single species and that a second species, Homo rudolfensis, is also present17 , 18 , 19 , 20 , 21 . Suporte adicional para a existência desta segunda espécie vem de uma reconstrução da mandíbula do espécime-tipo de H. habilis , OH 7, que descobriu que o espécime mostra uma arcada dentária primitiva longa e estreita que é incompatível com vários de H. rudolfensis espécimes 22 . Além disso, a nossa compreensão das origens do género foi impactada por várias descobertas de fósseis de Homo com mais de 2 milhões de anos atrás (Ma). 7 , 23 , 24 , 25 , 26 , 27 , 28 , incluindo uma mandíbula de Ledi Geraru, que representa o mais antigo Homo conhecido com 2,8 Ma 29 . Também é importante notar que alguns sugerem que os espécimes atualmente atribuídos ao H. habilis (particularmente OH 13, OH 24, OH 62 e KNM-ER 1813) podem de fato ser melhor classificados como Australopithecus devido à falta de sinapomorfias com Homo posteriores. 30 , 31 , 32 . Outros sugeriram que todos os hipodigmas de H. habilis e/ou H. rudolfensis deveriam ser transferidos para fora do gênero Homo 10 , 33 , 34 , 35 , 36 , embora várias análises filogenéticas apoiem a monofilia do gênero conforme definido atualmente 37 , com a inclusão do Australopithecus sediba 38 , 39 , ou com a exclusão de qualquer Homo floresiensis 38 ou estrela Homo 39 .
O hipodigma de H. habilis inclui numerosos restos dentários, com OH 7 preservando uma fileira de dentes mandibulares quase completa, OH 13 e OH 16 preservando a maioria dos dentes inferiores e maxilares, e OH 24 preservando a maioria das posições dos dentes maxilares pós-caninos. Várias características da dentição foram sugeridas como derivadas do Australopithecus ; a descrição inicial de H. habilis destacou dentes anteriores relativamente grandes e dentes pós-caninos estreitos bucolingualmente 1 , enquanto trabalhos subsequentes destacaram o pequeno tamanho dos dentes pós-caninos, os talonídeos pré-molares reduzidos e um M 3 reduzido que é semelhante em tamanho ao M 2 5. However, the usefulness of a number of these traits in distinguishing H. habilis from Australopithecus has been questioned22,40,41, and H. habilis teeth are considered by some to be relatively generalised in their morphology19. É importante ressaltar que esses estudos consideraram apenas a morfologia da superfície externa do esmalte (OES) dessas amostras, cuja morfologia original é frequentemente alterada ou removida pelos efeitos do desgaste dentário oclusal. Por exemplo, os primeiros molares inferiores de OH 7, OH 13 e OH 16 estão moderadamente a fortemente desgastados e, consequentemente, as análises da morfologia da coroa concentraram-se em medições 2D da superfície oclusal (por exemplo, dimensões lineares, áreas relativas das cúspides e formato do contorno da coroa). Criticamente, essas medidas não capturam os componentes verticais da altura da copa e da altura da cúspide, que se mostraram úteis para distinguir o Australopithecus do Homo. 42 , 43 .
Neste estudo, superamos as limitações do desgaste dentário variável e das medições 2D através de uma análise da coroa dentinária que está subjacente à capa de esmalte. Doravante referida como junção esmalte-dentina (EDJ), é uma superfície que se estabelece no início do desenvolvimento dentário e preserva a forma da membrana basal sobre a qual o esmalte é depositado. Além de reter os componentes verticais da altura da coroa e da cúspide, é onde a maioria das características que são conhecidas por terem relevância taxonômica são formadas durante a odontogênese. Como a JED não é remodelada ao longo da vida de um indivíduo, ela pode fornecer informações sobre a morfologia dentária que de outra forma não seriam possíveis devido aos efeitos do desgaste dentário. Vários estudos demonstraram que a morfologia do EDJ é taxonomicamente informativa em primatas existentes 42 , 44 , 45 , 46 e hominóides fósseis 43 , 47 , 48 , 49 , 50 , 51 , incluindo um estudo recente em que Zanolli et al. 52 sugeriu que o status Homo de vários fósseis sul-africanos de Sterkfontein, Drimolen e Swartkrans não é suportado com base na morfologia do EDJ. Os fósseis do desfiladeiro de Olduvai são cruciais para as questões que envolvem a sistemática do Homo inicial , mas pouco se sabe sobre sua morfologia dentária interna.
Aqui avaliamos a morfologia EDJ dos principais espécimes de Olduvai (Figs. 1 e 2 ), incluindo o espécime-tipo de H. habilis , OH 7, bem como os primeiros fósseis de Homo de Koobi Fora. Usando morfometria geométrica (GM) baseada em pontos de referência 3D, caracterizamos as principais diferenças de forma do EDJ entre o Australopithecus posterior e o Homo do Pleistoceno Médio (humanos modernos, Neandertais e Homo ; Tabela 1 ). Descobrimos que os principais espécimes de H. habilis , incluindo OH 7, retêm a condição primitiva e que várias características sugeridas anteriormente para caracterizar a dentição de H. habilis não são evidentes no EDJ. Também avaliamos a homogeneidade do hipodigma do H. habilis do ponto de vista odontológico, encontrando altos níveis de variação que em alguns casos excedem nossas expectativas de uma única espécie.
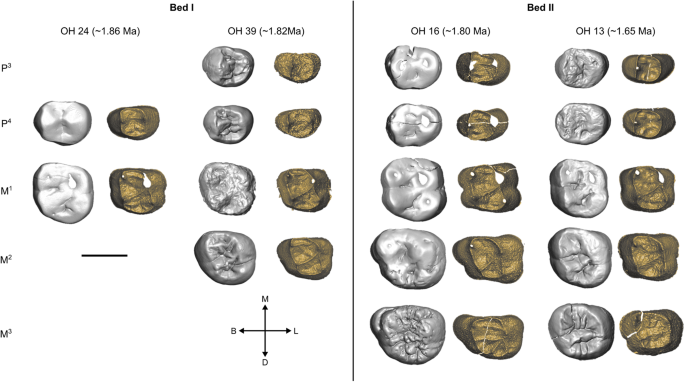
Três fileiras de dentes de H. habilis são mostradas, OH 7, OH 13, OH 16, OH 39. Cada uma é mostrada na vista oclusal na superfície externa do esmalte (esquerda) e na junção esmalte-dentina (direita). Todos os dentes são mostrados como do lado direito. Informações de datação e estratigrafia das refs. 69 , 72 , 73 .
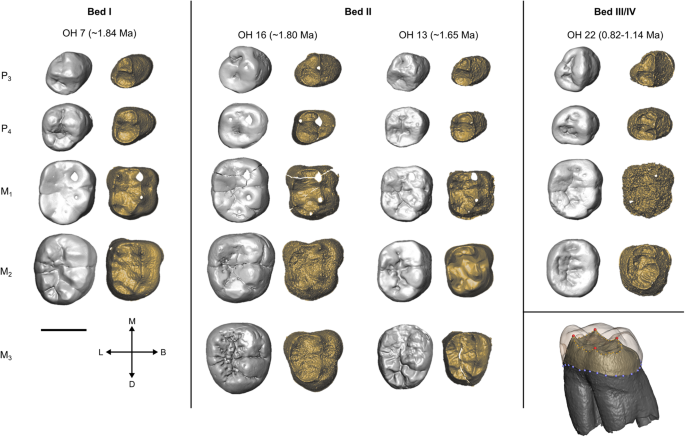
São mostradas três fileiras de dentes de H. habilis , OH 7, OH 13 e OH 16, bem como OH 22 que é atribuído a H. erectus . Cada um é mostrado na vista oclusal na superfície externa do esmalte (esquerda) e na junção esmalte-dentina (direita). Todos os dentes são mostrados como do lado direito. Um exemplo do protocolo de marcação para molares é mostrado no canto inferior direito (Vermelho = marcos fixos EDJ, amarelo = semimarcas da crista EDJ, azul = semimarcas da crista CEJ). Informações de datação e estratigrafia das refs. 69 , 72 , 73 , 87 , 88 .
Resultados
Padrões de forma geral
As análises de componentes principais (PCA) resumem a variação da forma do EDJ para três posições representativas dos dentes; P3 ( Fig. 3a ), P 4 (Fig. 4a ) e M 1 (Fig. 5a ). PCAs para todas as posições dos dentes pós-caninos podem ser encontrados na Figura 7 Complementar (versões html interativas desses gráficos estão disponíveis nos Arquivos de Dados Suplementares 1 – 10 ). Em cada caso, o primeiro componente principal (PC1) separa o Australopithecus ( Australopithecus afarensis e Australopithecus africanus ) da amostra posterior do Homo . Para a maioria das posições dos dentes, e particularmente dos pré-molares, PC1 representa uma alta proporção da variação total da forma presente na amostra. Isto corresponde a uma série de diferenças de forma presentes em toda a dentição pós-canina do grupo Homo posterior que os distingue do Australopithecus .
Essas mudanças de formato estão resumidas na Tabela 2 (ver também Tabelas Suplementares 4 e 5 ) e visualizadas usando wireframes (Figs. 3 b, 4 b, 5b ; Figs. Complementares 8 e 9 ). Uma diferença notável de forma entre o Australopithecus posterior e o Homo é um aumento relativo na altura do corpo da dentina (ver Figura 5 suplementar para uma explicação da terminologia usada para descrever a altura da coroa no EDJ) no último grupo; isso está presente em todas as posições dos dentes pós-caninos, embora seja menos pronunciado nos molares superiores. Outras diferenças de forma posteriormente O Homo , quando comparado com o Australopithecus , inclui uma redução na região da garra/talonídeo nos pré-molares, um protoconídeo relativamente mais alto e colocado mais distalmente nos molares inferiores e uma redução na crista marginal distal nos molares superiores (Tabela 2 ). A diferença na forma do EDJ entre o Homo posterior e o Australopithecus é significativa para todas as posições dos dentes nos testes de permutação (Tabela Suplementar 6 ).
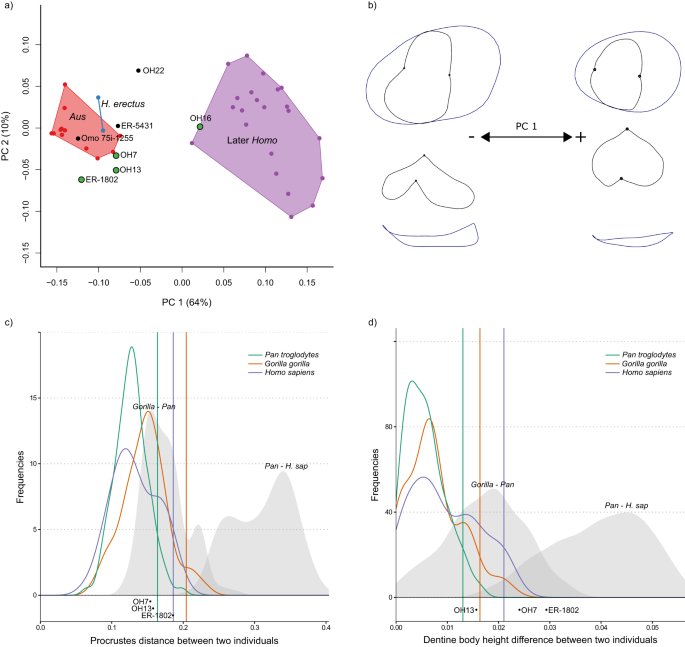
a PCA plot of P3 EDJ shape. Homo habilis specimens are represented by green points. Aus = Australopithecus. PC = principal component. b Wireframe images showing shape changes associated with the first principal component, showing landmark positions associated with extremes of PC1 (±1.5 standard deviations from the mean) in occlusal view (top) and lingual view (bottom). c Frequency plot of Procrustes distances between all possible pairs of individuals within (coloured lines) and between groups (grey fill). Vertical lines show the 95% limits of the within-group distributions. Below is shown the Procrustes distance between OH 16 and other H. habilis or early Homo specimens. Red = Gorilla gorilla, Green = Pan troglodytes, Purple = Homo sapiens. H. sap = H. sapiens. d Same as c for dentine body height. Source data are provided as a Source Data file.
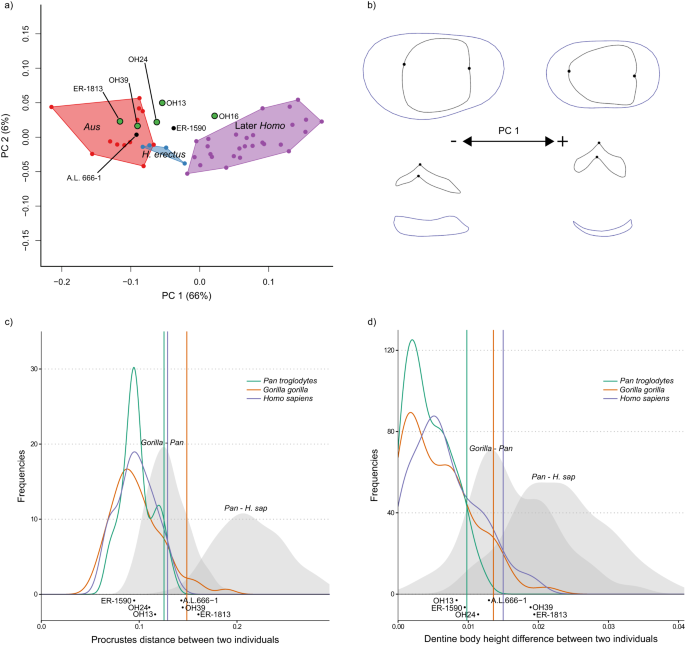
a PCA plot of P4 EDJ shape. Homo habilis specimens are represented by green points. Aus = Australopithecus. PC = principal component. b Wireframe images showing shape changes associated with the first principal component, showing landmark positions associated with extremes of PC1 (±1.5 standard deviations from the mean) in occlusal view (top) and lingual view (bottom). c Frequency plot of Procrustes distances between all possible pairs of individuals within (coloured lines) and between groups (grey fill). Vertical lines show the 95% limits of the within-group distributions. Below is shown the Procrustes distances between OH 16 and other H. habilis or early Homo specimens. Red = Gorilla gorilla, Green = Pan troglodytes, Purple = Homo sapiens. H. sap = H. sapiens. d Same as c for dentine body height. Source data are provided as a Source Data file.
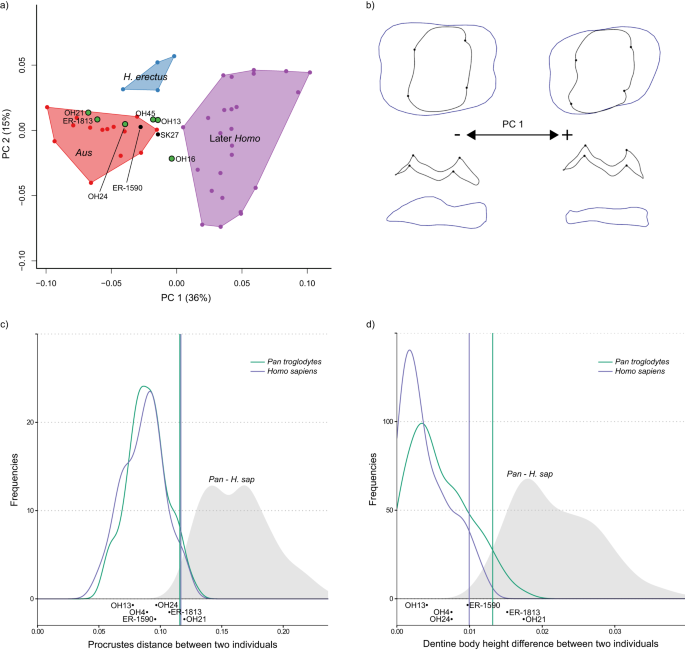
a PCA plot of M1 EDJ shape. Homo habilis specimens are represented by green points. Aus = Australopithecus. PC = principal component. b Wireframe images showing shape changes associated with the first principal component, showing landmark positions associated with extremes of PC1 (±1.5 standard deviations from the mean) in occlusal view (top) and lingual view (bottom). c Frequency plot of Procrustes distances between all possible pairs of individuals within (coloured lines) and between groups (grey fill). Vertical lines show the 95% limits of the within-group distributions. Below is shown the Procrustes distances between OH 16 and other H. habilis or early Homo specimens. Green = Pan troglodytes, Purple = Homo sapiens. H. sap = H. sapiens. d Same as c for dentine body height. Source data are provided as a Source Data file.
In these features that serve to distinguish between Australopithecus and later Homo at the EDJ, a number of H. habilis specimens retain the primitive condition, including the holotype, OH 7. For example, the mandibular premolar talonids are not reduced and the relative dentine body height is either short (M2) or intermediate (P3, P4, M1). Table 2 summarises the condition observed in selected Olduvai H. habilis specimens for these traits, and Supplementary Table 4 for specimens from Koobi Fora and A.L. 666-1. Other Bed I Olduvai specimens OH 4, OH 24, and OH 39 also approximate the Australopithecus condition, as do Koobi Fora H. habilis specimens KNM-ER 1502, KNM-ER 1802 and KNM-ER 1813 from Koobi Fora. This is evident from the GM analysis, in which these specimens plot closely to specimens of Australopithecus. Furthermore, we found only a few significant differences in postcanine tooth shape or size between Australopithecus and H. habilis in permutation tests (Supplementary Table 6). Several tooth positions of KNM-ER 5431 (Hominini gen. et sp. indet.) and A.L. 666-1 from Hadar (Homo aff. H. habilis ) também apresentam morfologias de EDJ semelhantes às do Australopithecus .
Entre os espécimes comumente atribuídos ao H. habilis , a condição mais derivada é vista em OH 16. Isto é particularmente evidente nos pré-molares, que apresentam uma altura relativa alta do corpo da dentina, garras/talonídeos reduzidos (P 3 , P 4), and an interrupted P3 mesial marginal ridge, features that are frequent in later Homo. The molars also show a tall dentine body, as well as taller molar dentine horns, particularly the mandibular molar protoconids and maxillary molar protocones (M2 and M3; Table 2). These features contribute to the position of OH 16 in PCAs, which for some tooth positions is closer to later Homo than Australopithecus. However, it is important to note that there are also a number of primitive aspects of the EDJ morphology in OH 16, such as the shape of the EDJ marginal ridges in the P4, M1, M1, and P3, que são semelhantes aos observados em outros espécimes de H. habilis e Australopithecus . OH 13 apresenta um padrão misto; o P 3 e M 2 são mais semelhantes ao OH 7 e KNM-ER 1802, enquanto o M 2 e M 3 parecem ser mais derivados, semelhantes ao OH 16.
As diferenças gerais de forma (medidas usando distâncias de Procrustes aos pares; Figs. 3 c, 4 c, 5c ; Fig. Complementar 10 ) entre OH 16 e outros espécimes de H. habilis P 4 estão principalmente dentro do limite de 95% dos grupos existentes, embora a forma é distinto do KNM-ER 1802, o formato do M 1 é distinto de OH 21, e o P 4 é distinto do KNM-ER 1813. No entanto, as diferenças mais claras entre OH 16 e outros espécimes de H. habilis estão na altura relativa do corpo da dentina. Quando esta característica é considerada isoladamente, OH 16 é distinto de OH 7 (P 3 -P 4 ), OH 21 (M 1 ), OH 39 (P 3 -P 4 ), KNM-ER 1802 (P 3 -P 4 ), KNM-ER 1813 (P 4 -M 2 ) e KNM-ER 1502 (M1 ) . Nestes casos, a diferença na altura relativa do corpo da dentina entre OH 16 e outros espécimes atribuídos a H. habilis excede o que seria esperado de uma única espécie (Figs. 3 d, 4 d, 5d ; Fig. Complementar 11 ).
Algumas posições dos dentes fornecem melhor distinção entre espécimes de Australopithecus e Homo primitivo do que outras em gráficos de PCA. Por exemplo, embora os M 2 s de H. habilis e H. erectus se sobreponham em grande parte aos do Australopithecus , há uma melhor distinção em outras posições dos dentes, particularmente o M 2 s. 3 (Figura 6 suplementar ), apesar de sua forma geralmente mais variável. Isto é bem ilustrado no exemplo do KNM-ER 1813. O P 4 , M 1 e M 2 desta parcela de espécime com Australopithecus , enquanto o M 3 is clearly distinct. In this case, H. habilis specimens occupy the positive end of PC3, which corresponds to a mesiodistally shortened crown, with an asymmetrical EDJ ridge due to particularly reduced mesiolingual and distobuccal crown corners. The M2 also provides a better distinction between H. habilis and Australopithecus in most cases; only KNM-ER 1813 plots within the Australopithecus range. This is also reflected in permutation tests, which find that the mean shapes of the M2 and M3diferem significativamente entre Australopithecus e H. habilis . Houve também diferença média significativa entre esses dois grupos no P 3 formato ; isso é impulsionado em parte pela condição derivada vista em OH 16, mas outros H. habilis P 3 também têm uma altura de corpo dentinário ligeiramente mais alta do que em Australopithecus , bem como uma base de coroa mais simétrica e uma ponta de corno de dentina protoconídeo colocada internamente (esta característica não pôde ser avaliada no OH 16 devido ao desgaste).
A análise de variáveis canônicas (CVA) foi realizada para investigar melhor as diferenças de forma entre Australopithecus , H. habilis e H. erectus (mais tarde o Homo foi excluído neste caso para fornecer a melhor distinção entre o Homo inicial e o Australopithecus ). Os resultados do CVA são mostrados nas Figuras Suplementares. 12 e 13 , e CVAs com validação cruzada (cvCVA) na Figura 14 Complementar . Os gráficos de CVA mostram a distinção entre Australopithecus e H. habilis em diversas posições dentárias, porém, na maioria dos casos, a separação entre esses grupos é severamente reduzida ou removida em cvCVAs, sugerindo que eles provavelmente representam diferenças espúrias entre grupos. As diferenças entre H. habilis e Australopithecus nos molares superiores são mantidas nos cvCVAs, indicando que estas representam diferenças reais de forma. Neles 3 , os espécimes de H. habilis são diferenciados do Australopithecus ao longo de CV2, que representa várias das mesmas características descritas acima (uma coroa mesiodistalmente curta, assimétrica na crista da JED). Eles 2 s são igualmente reduzidos mesiodistalmente, com uma crista EDJ assimétrica devido a um canto da coroa mesiolingual e distovestibular reduzido, enquanto o M 1 s são distinguidos ao longo de uma combinação de CV1 e CV2, correspondendo a diferenças sutis no formato da crista EDJ ao redor do metacone, na altura da coroa no lado lingual da coroa e no formato da junção cimento-esmalte (CEJ; embora seja importante notar que a diferença de forma entre Australopithecus e H. habilis M 1 s não foram significativos nos testes de permutação).
Vários espécimes de H. erectus incluídos aqui também representam mais de perto o Australopithecus posterior do que o Homo em PCAs, embora isso varie de acordo com o espécime e a posição do dente, e os espécimes de Sangiran são melhor distinguidos do Australopithecus do que aqueles de Koobi Fora. KNM-ER 1507 é o menos derivado; a amostra é plotada dentro da faixa de Australopithecus ao longo dos dois primeiros PCs para P 3 -M 2 e não mostra redução de talonídeos nos pré-molares, altura do corpo da dentina curta ou intermediária e coroa mesiodistalmente alongada em M 1 e M 2 (Tabela Suplementar 5 ). Estas características são mais semelhantes ao Australopithecus ou H. habilis do que a outros H. erectus , particularmente espécimes de Sangiran. KNM-ER 992 é mais derivado e, embora várias posições dos dentes se sobreponham ao Australopithecus ao longo do PC1, há um corpo dentinário moderadamente alto em cada dente, há alguma redução dos talonídeos pré-molares e o M 2 e M 3 mostram EDJ mais arredondado cristas marginais. Outros espécimes de Koobi Fora H. erectus estão localizados na periferia ou fora do Gama de Australopithecus nos primeiros 2 PCs (KNM-ER 3733 P 3 , P 4 , KNM-ER 806 M 1 ) ou são distinguidos ao longo de PC2 (KNM-ER 806 M 2 , M 3 , KNM-ER 807 M 1 , KNM-ER 1808 M 2 ) ou PC3 (KNM-ER 807 M 3 , KNM- ER 1808M 2 ; Figura Complementar 15 ). OH 22 é bem diferenciado de outros espécimes da amostra em PCAs, incluindo aqueles atribuídos a H. erectus . Em particular, a amostra ocupa uma posição positiva ao longo de PC2 para P 3 -M 1 e uma posição positiva ao longo de PC3 para M 2 (Fig. Complementar 15 ). Isto reflete uma série de características em toda a fileira dentária, incluindo a presença de uma crista marginal mesial baixa tanto nos pré-molares quanto nos molares, uma fóvea mesial relativamente alongada (presente em todos os dentes, mas particularmente notável no P3 ) e um metaconídeo curto. . Os espécimes de H. erectus são claramente distintos de Australopithecus e H. habilis em CVAs, e essas diferenças de grupo são mantidas em cvCVAs para a maioria das posições dentárias, exceto para o P 3 onde há sobreposição entre todos os três grupos e o M 2 onde KNM-ER 1808 e Sangiran 4 se sobrepõem ao Australopithecus . Esta distinção corresponde a uma série de características em toda a dentição, incluindo pré-molares com uma JCE (base da coroa) de formato oval mais simétrica, molares com cristas marginais da JED mais arredondadas e um hipocone molar superior reduzido.
Typicality-based classifications for unclassified specimens are presented in Supplementary Table 7. KNM-ER 5431 is mostly classified as Australopithecus. The premolars of KNM-ER 1590 and A.L. 666-1 are also classified as Australopithecus, however, the molars are frequently not classified into any of the groups. KNM-ER 1507 is classified mostly as H. habilis or Australopithecus. L26-1g, L398-573, and SK 847 are most often not classified into any group, while SK 27 is most often classified as Homo erectus, and Omo 75i-1255 is most often classified as H. habilis. OH 22 plots either close to H. erectus, or occupies a more extreme position along the CV that best separates H. erectus from other taxa. In particular, the P4 has an oval-shaped CEJ that is mesiodistally compressed and a clearly reduced talonid, while the M2tem uma crista EDJ arredondada, que é semelhante a outros espécimes de H. erectus , mas mais extrema em sua expressão. Como resultado dessas características, os dentes são classificados como H. erectus ou nenhum dos grupos.
Variation in CEJ shape in each anterior tooth position is summarised in Supplementary Fig. 16. The mandibular incisors provide a good distinction between Australopithecus and later Homo along PC1; this refers to the extent of the apical extension of the enamel on the labial and lingual sides of the tooth, which is more marked in Australopithecus, giving the CEJ a sinusoidal shape. In this respect, OH 7 and OH 16 retain the Australopithecus condition, while two H. erectus specimens (KNM-ER 820 and KNM-WT 15000) show a much flatter CEJ, even more so than in later Homo specimens. This CEJ shape difference is less marked in the other anterior tooth positions, and as a result, there is more overlap between groups in these cases.
Buccolingually narrow postcanines
Na superfície do esmalte, a maioria dos dentes pós-caninos do H. habilis são bucolingualmente mais estreitos que os do Australopithecus , particularmente na dentição mandibular (Tabelas Suplementares 1 e 2 ). No entanto, esse padrão é menos evidente na JED; em vez disso, há variação por espécime, posição do dente e por qual parte da JED está sendo considerada (Figuras Complementares 1 e 2 ). Esta característica é discutida em detalhes na Nota Complementar 1 .
Padrões de tamanho na fileira de dentes
A Figura 6 mostra os padrões de tamanho cervical ao longo da fileira dentária em H. habilis quando comparado com Australopithecus, H. erectus e, posteriormente, Homo (comparações entre H. erectus e outros grupos, incluindo Homo sp., estão disponíveis na Figura 17 Complementar , versões html interativas de cada gráfico de tamanho de centróide estão disponíveis nos arquivos de Dados Suplementares 11 a 20 e todos os dados de tamanho de centróide estão disponíveis em Dados Suplementares 21 ). O tamanho dos dentes mandibulares de H. habilis apresenta um grande grau de sobreposição com os do Australopithecus , e não encontramos diferenças significativas de tamanho entre os dois táxons (Tabela Suplementar 6 ). No OH 7, todas as posições dos dentes preservados estão dentro da faixa do Australopithecus . Uma possível exceção a esse padrão é o OH 13, que é o menor espécime de H. habilis incluído aqui, e para o qual o canino e o P 4 estão fora da do Australopithecus faixa de variação . OH 13 também difere de outros espécimes de H. habilis por ter um P 3 maior que o canino; em OH 7 e OH 16 o P 3 é menor. O P 3 > O padrão C observado em OH 13 é semelhante ao observado em vários espécimes de H. erectus (KNM-ER 820, 992, 1507 e KNM-WT 15000). Há também uma sobreposição no tamanho dos dentes maxilares entre H. habilis e Australopithecus , embora os dentes pós-caninos ocupem a extremidade inferior da faixa do Australopithecus , e alguns M. 2s and M3s are smaller. The teeth of OH 16 are large, similar to other H. habilis and Australopithecus. The anterior teeth in H. habilis are variable; OH 15 (C) and OH 16 (I1) occupy the larger end or are slightly above, the Australopithecus range of variation, while KNM-ER 1813 (I2-C) and OH 39 (I1, C) occupy the lower end or are slightly below the Australopithecus range. H. habilis specimens mostly have maxillary premolars that are similar in size (or in OH 39, a P4 that is slightly smaller), whereas Australopithecus usually has a P4 > P3 pattern. However, there are also some A. afarensis specimens in which the P3 é maior (AL 199-1 e AL 200-1a).
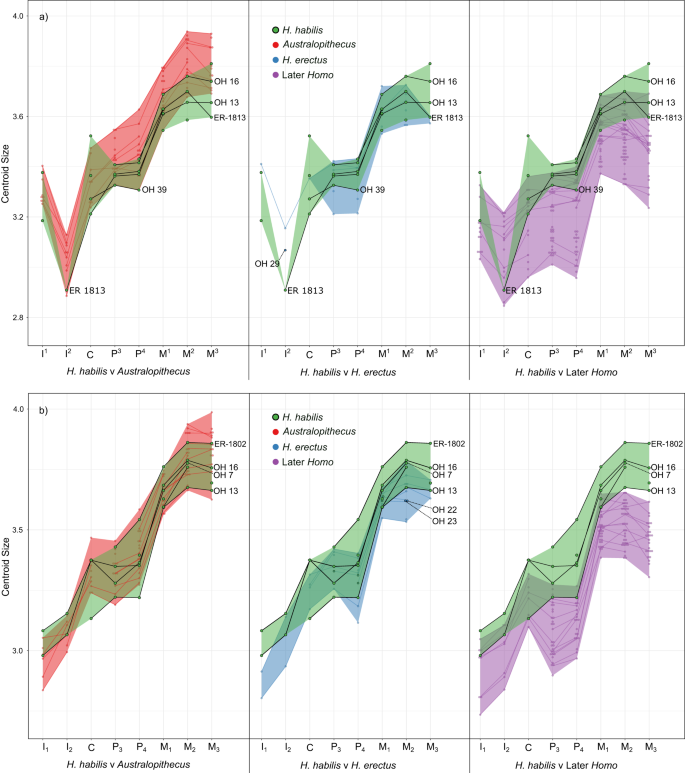
Plots show the natural logarithm of the centroid size of the CEJ ridge landmark set, calculated separately for a the maxillary tooth row, and b the mandibular tooth row. In both plots, Australopithecus is shown in red, later Homo in purple, and Homo habilis in green. Shaded areas represent the range of centroid sizes for each group, and lines connect teeth from the same individual. H. habilis specimens that preserve complete or nearly complete tooth rows are labelled and connected using black lines. Source data are available in Supplementary Data 21.
Os dentes inferiores do H. habilis posterior são geralmente maiores que os do Homo , embora a diferença seja significativa apenas nas posições dos dentes posteriores para as quais existem amostras maiores (Tabela Suplementar 6 ). Os dentes superiores do H. habilis mostram mais sobreposição com os do Homo posterior , mas os pré-molares e molares ainda eram significativamente diferentes. Não houve diferenças significativas entre H. habilis e H. erectus , embora vários espécimes Sangiran de H. erectus e KNM-ER 820 (P 4 -M 2 ) estejam abaixo da faixa de H. habilis . Descobriu-se que a maioria das posições dos dentes pós-caninos em H. erectus eram significativamente diferentes das do Homo posterior (exceto M. 1 e M 3 ) e apenas um pequeno número de posições dentárias (M 3 , M 2 e M 3 ) foram significativamente diferentes do Australopithecus . O tamanho dos dentes de OH 22 e OH 23 é semelhante a outros espécimes de H. erectus (Fig. Complementar 17 ), exceto que ambos apresentam um padrão M 1 > M 2 , enquanto KNM-ER 806, KNM-ER 992, KNM -ER 1507, KNM-WT 15000 e Sangiran 1b mostram o padrão oposto. Os dentes do KNM-ER 1590 são grandes comparados aos do H. habilis , e possui um canino que é semelhante em tamanho de colo ao P. 3 , que é raro no Australopithecus , e não está presente em nenhum dos espécimes de H. habilis que preservam estes dentes (OH39 e KNM-ER 1813), ou KNM-WT 15000, mas é visto em alguns Homo posteriores . No AL 666-1, os pré-molares são ligeiramente maiores que os do H. habilis , mas o M 1 e M 2 estão dentro da faixa H. habilis .
Discussão
A suite of EDJ features distinguishes the postcanine teeth of later Homo from those of Australopithecus (A. afarensis and A. africanus), including the relative height of the dentine body and reduction of the premolar talons and talonids. However, when considering these traits in early Homo, we find that key H. habilis specimens such as OH 7, OH 24, and KNM-ER 1813 largely retain the Australopithecus condition. The EDJ shape and size patterns across the tooth row in these H. habilis specimens accord more closely with Australopithecus than later Homo, and we fail to find statistically significant differences between our H. habilis and Australopithecus samples in size in any tooth position, and only find significant shape differences in three tooth positions (P3, M2, M3) (Tabela Suplementar 6 ).
Na verdade, encontramos o H. habilis M 2 and M3 shapes to be distinct from those of other groups more broadly. Homo habilis M2s (n = 4), and especially M3s (n = 4), have a mesiodistally short, asymmetrical EDJ ridge. In the M2s, this is due to a reduced crown distobuccally, while the M3s are reduced mesiolingually and distobuccally. The M3s also frequently show a metacone dentine horn that is mesially and internally placed, but this is less pronounced in the M2S. Esta combinação de características impulsiona a separação de H. habilis em AVCs para ambas as posições dentárias, e a separação de M. 3 s ao longo do PC3 do PCA (Fig. Complementar 7 ), e os distingue do Australopithecus , mais tarde Homo , e do H. erectus ( H. erectus M 3 s também têm uma crista EDJ que é encurtada mesiodistalmente, mas são menos assimétricas e mais arredondadas na vista oclusal). Esta morfologia é evidente em alguns dos espécimes geralmente mais derivados de H. habilis (OH 13 e OH 16), mas também está presente no M 3 do KNM-ER 1813, que por outro lado é muito semelhante ao Australopithecus . Os molares distais são conhecidos por serem altamente variáveis, enquanto os molares M 1 em particular tem uma morfologia mais estável 53 , 54 . Este nível mais baixo de variação no M 1 pode ser considerado útil para a sistemática dos hominídeos, pois reduz a variação intraespecífica 55 , 56 , 57 , 58 , entretanto, neste caso, parece que as diferenças na morfologia da JED entre nossas amostras de H. habilis e Australopithecus são mais claras nos molares distais (M 2 e M 3 ). Isto poderia sugerir que diferenças morfológicas relativamente pequenas no M 1 s podem ser exagerados ao longo da fileira molar, de modo que sejam mais claros no M 2 e M 3 , embora isso exija uma investigação mais aprofundada. Houve também uma diferença significativa na forma do P3 entre nossas amostras de H. habilis e Australopithecus ; impulsionado por uma altura do corpo da dentina ligeiramente aumentada (mais extrema em OH 16), base da coroa simétrica e ponta do corno de dentina protoconídeo colocada internamente.
Existem características dentárias OES derivadas em H. habilis , incluindo um estreitamento vestíbulo-lingual da dentição pós-canina 1 , 41 , 59 (Tabelas Suplementares 1 e 2 ). No entanto, no EDJ esta característica é menos clara e varia de acordo com a amostra, a posição do dente e por qual parte da coroa do dente contribui para a diferença de forma. É possível que diferenças na distribuição e espessura relativa do esmalte ao longo da coroa contribuam para esta característica, embora isto requeira investigação mais aprofundada. Da mesma forma, não encontramos evidências consistentes de dentes anteriores grandes em H. habilis na base da coroa, quando comparado com Australopithecus ou Homo posterior (Tabela Suplementar 6 ). Para uma discussão mais detalhada de ambas as características, consulte a Nota Complementar 1 . Quam et al. também descobriu que o M 1 of KNM-ER 1813 and OH 21 show derived relative cusp areas compared with Australopithecus and other H. habilis specimens55; this is not clear at the EDJ, where both specimens show a primitive overall M1forma em PCAs, embora a relação entre a morfologia da JED e as áreas relativas das cúspides seja provavelmente complexa, com contribuições de múltiplos aspectos da forma da JED, bem como da espessura e distribuição regional do esmalte. É possível que existam características dentárias adicionais não identificadas aqui que sejam autapomórficas para H. habilis , ou que efetivamente distingam o Homo primitivo do Australopithecus . Em particular, este pode ser o caso de aspectos da morfologia que não são capturados em nossa análise GM (como a bacia oclusal ou as faces laterais da JED), ou características dentárias discretas. No entanto, como mostrado recentemente para cúspides acessórias de molares inferiores, tais características dentárias discretas frequentemente mostram padrões de expressão complexos que podem ser problemáticos em estudos de sistemática de hominídeos 60 .
A falta de características dentárias claramente derivadas de H. habilis no EDJ na maioria das posições dos dentes, combinada com a morfologia geral do EDJ pós-canino primitivo em vários espécimes-chave de H. habilis , sugere uma morfologia dentária endoestrutural semelhante ao Australopithecus nos primeiros membros de o gênero Homo . Alguns dos espécimes que apresentam uma morfologia mais primitiva, como KNM-ER 1813 e OH 24, foram sugeridos por alguns autores como pertencentes ao Australopithecus e não ao Homo. 30 , 31 , 32 . OH 7 é importante nesse aspecto, entretanto, pois combina um padrão dentário e gnático em grande parte semelhante ao do Australopithecus com um volume endocraniano aumentado que é consistente com outros membros iniciais do gênero Homo. 22 , 61 . Um estudo recente de Zanolli et al. 52 analisaram a morfologia EDJ de 23 espécimes de cavernas sul-africanas que foram sugeridas como pertencentes ao gênero Homo , descobrindo que apenas quatro dos espécimes analisados poderiam ser inequivocamente atribuídos ao Homo , enquanto outros mostram um padrão dentário claramente mais semelhante ao Australopithecus ou Parantropo . Ao contrário dos espécimes da África Oriental, como OH 7, OH 24 e KNM-ER 1813, muitos dos espécimes da África Austral analisados são dentes isolados e, portanto, as avaliações do seu estatuto taxonómico dependem necessariamente fortemente da morfologia dentária. Uma vez que os fósseis de hominídeos do Pleistoceno Inferior da África Oriental geralmente preservam mais estruturas anatômicas ao longo do esqueleto, estudos futuros terão que integrar toda a morfologia disponível: dentária, craniana e pós-craniana, ao avaliar as afinidades taxonômicas de espécimes individuais de hominídeos.
Alternativamente, alguns defenderam que todos os hipodigmas de H. habilis e/ou H. rudolfensis fossem transferidos para fora do gênero Homo. 10,33,34,35,36. They suggest that the inclusion of these species in Homo could render the genus paraphyletic, which is supported by some older phylogenetic analyses33,62. However, it is important to mention that several more recent phylogenetic studies have supported the monophyly of Homo even with the inclusion of H. habilis/rudolfensis37,63, although in certain instances, maintaining this monophyly might necessitate reconsideration of the generic status of A. sediba, H. naledi and/or H. floresiensis38,39. It has also been argued that species included in Homo should occupy a coherent adaptive grade33, and that plesiomorphic features across the H. habilis skeleton (e.g., refs. 22,64,65,66) suggest that the adaptive strategy of H. habilis is more similar to Australopithecus than H. sapiens33,36. Others have argued against the requirement for genera to be adaptively unified16 , enfatizando que as adaptações observadas em H. sapiens evoluíram em um padrão escalonado e em mosaico e que a distinção entre Australopithecus e os primeiros membros do gênero Homo é necessariamente pequena 15 , 16 . Os resultados aqui apresentados destacam uma série de características plesiomórficas em toda a dentição do H. habilis que enfatizam a distinção morfológica e adaptativa da espécie em relação aos membros posteriores do gênero Homo , mas se isso justifica a remoção da espécie do Homo depende da definição do gênero.
Ao considerar o H. erectus , há também alguma sobreposição com o Australopithecus em PCAs para várias posições dentárias, no entanto, o H. erectus é melhor distinguido do Australopithecus nas análises de AVC, e há diferenças significativas na forma da JED entre os dois táxons em todos os dentes mandibulares. posições, bem como P 4 e M 2 . Os espécimes de H. erectus são diferenciados do Australopithecus por uma série de características, incluindo uma redução da garra/talonídeo e um formato CEJ mais simétrico nos pré-molares (exceto P 3 ) e cristas EDJ mais arredondadas nos molares. Existem também diferenças significativas no tamanho de alguns molares posteriores (M 3 , M2 , M3 ) . No geral, estas diferenças são mais pronunciadas do que as diferenças entre Australopithecus e H. habilis , e são indicativas de uma morfologia dentária mais derivada em H. erectus , mesmo nos espécimes do Pleistoceno Inferior aqui incluídos. O pequeno tamanho dos molares posteriores é consistente com a descoberta de que o H. erectus deriva de ter M 3 s relativamente reduzidos (em relação ao M 1 ), enquanto os primeiros espécimes do Homo são mais semelhantes ao Australopithecus. 67 (embora a amostra de H. erectus na ref. 67 é em sua maioria mais jovem do que os espécimes do Pleistoceno Inferior incluídos aqui). Descobriu-se que um espécime de H. erectus , KNM-ER 1507, tinha uma morfologia EDJ mais primitiva, com P 3 -M 2 semelhante a H. habilis ou Australopithecus mais do que outros espécimes de H. erectus , o que é apoiado por classificações baseadas em CVA (Tabela Suplementar 7 ). É possível que este espécime fosse melhor atribuído ao H. habilis , no entanto, isto deve ser avaliado posteriormente com referência à morfologia da mandíbula, bem como dos molares decíduos, particularmente porque atribuições anteriores deste espécime ao H. erectus têm confiou nesses aspectos da morfologia 5,68. Análises anteriores deste espécime também delinearam semelhanças com outra mandíbula juvenil, KNM-ER 820, que não pôde ser incluída nas análises de EDJ aqui devido à má distinção de tecidos nas varreduras. OH 22 compartilha algumas semelhanças com espécimes de H. erectus de Koobi Fora e Sangiran, mas também é distinto em características como o alongamento da fóvea mesial (particularmente no P 3 ), e mostra expressão mais extrema de algumas características observadas no início do H. . espécimes erectus , como redução de talonídeos nas cristas marginais P 4 e arredondadas M 2 EDJ. Esta combinação é consistente com a idade mais recente do espécime em comparação com os outros espécimes de H. erectus incluídos aqui, embora a comparação com espécimes mais jovens de H. erectus fosse necessária para avaliar completamente as afinidades taxonômicas do OH 22.
Encontramos um nível considerável de variação dentro do hipodigma dentário do H. habilis que em alguns casos excede o esperado de uma única espécie (Figs. 3 , 4 e 5 ; Figs. Complementares 10 e 11 ). Os espécimes do leito I Olduvai geralmente mostram uma do Australopithecus morfologia mais semelhante à ; OH 7 (~1,84 Ma 69 ) e OH 24 (~1,86 Ma 69 ) estão próximos ou dentro da faixa de variação do Australopithecus para todas as posições dentárias preservadas (Fig. Complementar 6 ), enquanto OH 39 (~ 1,82 Ma 69 ) está dentro da faixa do Australopithecus para ambos os pré-molares, mas tem um M mais derivado 2 , e tem dentes geralmente pequenos, alguns dos quais são menores que os do Australopithecus (I 1 , M 2 ). Dos espécimes Koobi Fora, KNM-ER 1802 é o mais antigo com 1,98-2,09 Ma 70 , e os dentes pós-caninos são consistentemente semelhantes aos do Australopithecus na morfologia geral, além de serem maiores do que qualquer outro de H. habilis espécime . Da mesma forma, os dentes do KNM-ER 1813 (1,78-1,95 Ma 71 ) são em grande parte primitivos em morfologia, embora com um M distinto 3 e geralmente dentes pequenos.
mais bem preservados Os espécimes de H. habilis do leito II de Olduvai são OH 13 e OH 16, ambos preservando fileiras de dentes pós-caninos inferiores e superiores inteiras. OH 13 é do meio do Leito II e é o espécime mais jovem atribuído a H. habilis , com idade sugerida de 1,65 Ma 72 , 73 . OH 16 is older, deriving from the lower section of Bed II. The specimen comes from the above marker Tuff IF, which makes it no older than 1.8 Ma69,72. Some early criticisms of the naming of the species H. habilis suggested that these Bed II specimens represented a different taxa to the older Bed I material74,75, while others suggested the Bed II specimens could be distinguished from each other, with either OH 1376,77 or OH 1678 more similar to H. erectus specimens known at the time. We find that OH 16 is clearly more derived than other H. habilis specimens in several aspects of postcanine EDJ morphology, and the specimen displays a suite of derived traits including an increased relative dentine body height in the postcanine dentition, which is typical of later Homo. In this regard OH 16 differs significantly from OH 7 and KNM-ER 1802 (lower premolars), OH 39 (upper premolars), and KNM-ER 1813 (P3-M2); these differences in dentine body height exceed the level of variation expected within a species, based on extant hominines (Supplementary Fig. 11). However, it is important to note that there are key differences between OH 16 and later Homo, in particular, the specimen retains several features that are clearly primitive (as seen in other H. habilis specimens), particularly in the EDJ marginal ridge shape of the P4, M1, P3, and M1.
As características derivadas presentes na dentição OH 16 poderiam sugerir evolução dentro da linhagem H. habilis ao longo do tempo. No entanto, apesar da sua idade mais jovem, o OH 13 não é tão derivado; O P 4 , M 1 e M 2 são semelhantes ao OH 16, mas para uma série de posições dentárias, como o P 3 , a morfologia é mais semelhante aos espécimes do leito I, como o OH 7. O espécime, portanto, mostra uma morfologia intermediária geral. É possível que as diferenças entre ele e OH 16 representem variação normal dentro da amostra do Leito II; as diferenças entre os dois espécimes estão dentro dos limites esperados para uma espécie baseada na maioria das amostras existentes para todas as posições dos dentes (Figs. Complementares 10 e 11 ). Deve-se notar que outro espécime que se sugere derivar do leito II inferior, OH 21, difere significativamente do M 1 de OH 16 (na distância de Procrustes e na altura do corpo da dentina) e é mais semelhante em morfologia ao KNM-ER 1813. No entanto, há alguma incerteza sobre a proveniência deste espécime, que foi encontrado na superfície de um depósito perturbado 41 . No geral, embora seja claro que há uma variação substancial dentro dos espécimes atribuídos ao H. habilis , provavelmente demais para ser incluída em uma única espécie, não está claro até que ponto a profundidade do tempo influencia essa variação, se é que influencia.
O alto nível de variabilidade no registro fóssil do Homo primitivo levou muitos autores a sugerir que múltiplas espécies estão representadas 17 , 19 , 20 , 21 , 22 , embora haja pouco acordo sobre a disposição dos espécimes em grupos. Infelizmente, o KNM-ER 1470, que é central para este debate, não preserva nenhuma coroa dentária, enquanto a maxila OH 65, sugerida como muito semelhante ao KNM-ER 1470 31 , could not be included in this sample. Equally, several specimens suggested to belong to this second species of early Homo such as KNM-ER 60000, KNM-ER 62000, and KNM-ER 6200321, could not yet be included in this sample as they have not yet been microCT scanned. KNM-ER 1590, which was included here, has been likened to KNM-ER 1470, although this is mostly based on the large size of the vault5,79. We find that the teeth of this specimen are more derived along PC1 than KNM-ER 1813, and the M2 is particularly derived. All teeth are larger in size than those of H. habilis, which is consistent with a recent study of canine size that found that including KNM-ER 1590 in a sample of H. habilis s.l. led to greater variability than seen in modern humans80. A forma EDJ não é tão derivada quanto OH 16 na altura do corpo da dentina, redução dos talonídeos ou redução na altura das cúspides, mas mostra uma morfologia intermediária nessas características. Nos AVCs, os pré-molares do KNM-ER 1590 são semelhantes aos do Australopithecus , e classificam-se com o grupo. Os molares apresentam sinal mais misto, com algumas classificações com Homo , mas mais frequentemente não se classificando com nenhum grupo. Isto seria consistente com o espécime pertencente a uma segunda espécie de Homo primitivo não erectus . No entanto, um estudo adicional incluindo alguns dos espécimes mencionados acima seria necessário para uma avaliação dentária detalhada desta questão. Outro espécime importante é AL 666-1, uma maxila de 2,3 Ma de Hadar. O espécime foi atribuído ao Homo aff. H. habilis 25 , mas mais recentemente descobriu-se que o formato da arcada não correspondia ao OH 7 (e, portanto, ao H. habilis ) e, em vez disso, mostra fileiras de dentes pós-caninos encurtadas, como visto nos primeiros H. erectus e H. sapiens. 22 . Descobrimos que a morfologia dos pré-molares deste espécime é bastante primitiva, com ambos os pré-molares dentro da faixa do Australopithecus , mas também próximos aos do OH 39. Os pré-molares também são classificados como Australopithecus nas análises de CVA. Como em outros espécimes Homo primitivos , o M 2 é mais derivado, mostrando, entre outras características, uma crista EDJ encurtada mesiodistalmente e plotando entre o Australopithecus posteriores e grupos Homo . Eles 2 classifica como Australopithecus , H. habilis ou nenhum dos grupos nas análises de CVA, dependendo de quantos PCs estão incluídos. A morfologia dentária não é semelhante à dos primeiros espécimes de H. erectus incluídos aqui. Isto sugere que este espécime combina uma morfologia dentária primitiva geral, semelhante ao Australopithecus e alguns outros espécimes do Homo primitivo , com uma forma de arcada significativamente mais derivada. Dos restantes não atribuídos ou Homo sp. espécimes, não houve atribuições claras para L26-1g (M 1 ), L398-573 (M 3 ) ou SK 847 (M 3 ); ambos M 3 s ocupam uma posição intermediária nas parcelas de CVA, no entanto, a amostra comparativa de H. erectus aqui é de apenas 2 espécimes, portanto é improvável que esta análise capture toda a gama de variação neste táxon. No entanto, o Omo 75i-1255 (P 3 ) foi considerado semelhante ao H. habilis , enquanto o SK 27 (M 1) foi considerado mais semelhante ao H. erectus .
To conclude, we find that the postcanine EDJ morphology of a number of key specimens of H. habilis, including the type specimen OH 7, is very similar to that of Australopithecus. These results suggest that dental changes associated with later Homo were not present in the earliest members of the genus. The EDJ morphology of H. habilis is for the most part generalised, which is consistent with the primitive nature of the dental arcade and mandibular corpus in the species22, as well as important aspects of the postcrania such as the hand morphology64,65 and relative limb proportions66. As such, features linking specimens such as OH 7 to the genus Homo are largely limited to the neurocranium. Considering the skeleton as a whole, this indicates that H. habilis possessed a number of plesiomorphic features, which is relevant for ongoing discussions over the genus-level designation of H. habilis16,36,81,82. Finally, our results highlight excessive levels of dental variation within the hypodigm as currently defined; Bed II specimen OH 16 shows a suite of derived dental traits that clearly distinguish it from other H. habilis specimens, but do not appear to align the specimen with early African H. erectus.
Métodos
Permissões, divulgação e envolvimento
Os dados de digitalização de espécimes do desfiladeiro de Olduvai foram coletados para este estudo após aceitação de uma licença de pesquisa (#2017-182-NA-2016-304) para a Comissão de Ciência e Tecnologia da Tanzânia (COSTECH) e sob um memorando de entendimento assinado entre MMS e o Museu Nacional da Tanzânia. Todos os dados originais da tomografia computadorizada, uma estação de trabalho de computador de última geração (com instalação do pacote Avizo Software) e impressões 3D derivadas dessas tomografias foram depositados no museu. Um estágio de treinamento na Universidade de Kent, sob a supervisão do MMS, foi fornecido à AG para se tornar proficiente no processamento e análise de tomografias computadorizadas. Além disso, atividades de divulgação foram organizadas por AG no museu para visitas locais de alunos de escolas primárias, usando materiais didáticos personalizados disponíveis para download no Arquivo de Registros Fósseis Humanos ( https://human-fossil-record.org/index.php?/category /6687 ). Isto incluiu folhas de atividades interativas personalizadas, criadas e financiadas pela MMS, focadas na evolução humana e Homo habilis na Tanzânia, especificamente.
Amostra do estudo
A amostra do estudo de hominídeos está resumida na Tabela 1 , e uma lista detalhada de espécimes de hominídeos pode ser encontrada em Dados Suplementares 21 . A amostra consiste em 712 dentes de hominídeos (91 incisivos, 54 caninos, 212 pré-molares e 355 molares. Além disso, uma amostra de pós-caninos de macacos existentes ( Gorilla gorilla e Pan troglodytes , n = 199) foi coletada para comparar os níveis de variação em H. habilis com táxons existentes (juntamente com os Homo sapiens incluídos na amostra principal do estudo) espécimes de ;
A amostra de hominídeos inclui 48 dentes atribuídos a H. habilis de Olduvai Gorge (OH 4, 7, 13, 15, 16, 21, 24, 39, 45; Figs. 1 e 2 ) e 13 de Koobi Fora (KNM-ER 1502, 1802, 1813; Figuras Suplementares 3 e 4 ). Uma lista completa de espécimes que preservam coroas dentárias inteiras que são aqui consideradas pertencentes ao H. habilis está disponível na Tabela Suplementar 3 ; para o pequeno número de espécimes que não puderam ser incluídos no presente estudo, as razões estão descritas nesta tabela.
Também incluímos vários espécimes que foram sugeridos por alguns autores como pertencentes ao H. habilis, mas não possuem atribuições seguras em nível de espécie (SK 27, SK 847, L26-1g, L398-573 e Omo 75i-1255). Esses espécimes são separados da amostra de H. habilis para fins comparativos. Além disso, está incluído KNM-ER 1590, que foi sugerido pertencer a H. rudolfensis através de suas semelhanças cranianas com KNM-ER 1470 5 , 19 . AL 666-1 é uma maxila de 2,3 Ma de Hadar atribuída ao Homo aff. H. habilis 25 , mas é aqui agrupado separadamente como Homo sp. KNM-ER 5431 não recebeu uma designação de gênero, mas foi sugerido que mostra características semelhantes ao Australopithecus afarensis ou ao antigo Homo 5 , 23 , 83 .
A amostra de H. erectus é limitada a espécimes de H. erectus do Pleistoceno Inferior de Koobi Fora (KNM-ER 806, 807, 992, 1507, 1808, 1812, 3733) e Sangiran (Sangiran 1a, 4, 7-3a-c, 7 -10, 7-20, 7-26, 7-27, 7-30, 7-32, 7-53, 7-65, 7-89) já que o H. erectus antigo fornece a comparação mais apropriada para o Homo material inicial . KNM-ER 807 foi alternadamente atribuído ao Homo sp 5 . ou H. erectus 84 , mas está incluído aqui como H. erectus . A morfologia EDJ de KNM-ER 1507 parece diferir de outros espécimes de H. erectus e, portanto, foi excluída dos testes de permutação e projetada em análises de CVA para testar suas afinidades de grupo. OH 22, OH 23 e OH 29 também são atribuídos a H. erectus , mas são separados da amostra principal de H. erectus em números, pois são mais jovens que os espécimes de Koobi Fora e Sangiran (todos são do Camada III ou superior, <1,14 Ma; 72 , 85 , 86 , 87 , 88 ). Avaliamos esse hipodigma dentário do Homo inicial contra uma agrupada de Australopithecus amostra , consistindo de A. afarensis e Australopithecus africanus , e uma amostra agrupada de Homo do Pleistoceno Médio posterior, consistindo de humanos modernos, Neandertais e Homo . Esses grupos são agrupados para nos permitir identificar as mudanças gerais na forma do EDJ que ocorreram durante a evolução do gênero Homo e, subsequentemente, identificar quais dessas características estão presentes no H. habilis .
Terminology
Throughout the manuscript, we refer to the morphology of the EDJ rather than the OES unless otherwise specified. At the EDJ it is clear that the height of the crown can be divided into two components: dentine body height and dentine horn height. Dentine body height refers to the distance between the cervix and the marginal ridges that encircle the occlusal basin, while dentine horn height refers to the distance between the marginal ridges and the tip of the tallest dentine horn (Supplementary Fig. 5). Total crown height is the combination of the two. Unless otherwise specified, these aspects of crown height are described relative to overall crown shape, rather than as an absolute measure.
Microtomografia
As varreduras microtomográficas da amostra foram obtidas usando um SkyScan 1172 ou SkyScan 1173 a 100–130 kV e 90–130 microA, um scanner BIR ACTIS 225/300 a 130 kV e 100–120 microA, ou um Diondo d3 a 100–140 kv e 100–140 microA e reconstruídos como pilhas tif de 16 bits com uma resolução de voxel isométrica variando de 13 a 45 mícrons. Sete dentes de H. erectus (Sangiran 7-3b, 7-3c, 7-20, 7-26, 7-53, 7-65, 7-89) foram fotografados usando microtomografia de nêutrons no centro de imagem ANTARES localizado na e FRM Reator II da Universidade Técnica de Munique, Alemanha (para detalhes técnicos ver ref. 89 ).
Segmentação
Para a maioria das amostras, as pilhas TIFF foram filtradas usando apenas um filtro de média de menor variância (tamanho do kernel um) ou um filtro mediano 3D (tamanho do kernel de três) seguido por um filtro de média de menor variância (tamanho do kernel de três), implementado usando software de código aberto MIA 90 . Pilhas de imagens filtradas foram usadas para segmentar o esmalte da dentina usando um algoritmo de bacia hidrográfica de crescimento de sementes empregado por meio de um plug-in personalizado empregado no Avizo 6.3 (Visualisation Sciences Group), antes de serem verificados manualmente (o plug-in personalizado do Avizo 6.3 não está disponível publicamente, mas versões modernas do Avizo, por exemplo, Avizo 3D 2022.1, possuem módulos de segmentação de bacias hidrográficas integrados de forma semelhante). Em amostras com contraste particularmente fraco entre os tipos de tecido, o esmalte e a dentina foram segmentados usando a ferramenta varinha mágica no Avizo v.8.0 (FEI Visualization Sciences Group) usando interpolação e correções manuais com base nas informações das pilhas de imagens filtradas e não filtradas. Nas regiões onde o esmalte e a dentina não puderam ser demarcados com precisão, ferramentas de interpolação foram utilizadas para estender a partir das regiões vizinhas nas quais havia melhor contraste tecidual. Em todas as segmentações, é dada especial atenção às cristas marginais, onde os pontos de referência são colocados, com verificações em pilhas de imagens não filtradas e filtradas, se necessário. Depois que o esmalte e a dentina foram segmentados, um modelo de superfície baseado em triângulo do EDJ foi produzido usando o parâmetro de suavização irrestrita no Avizo e depois salvo em formato de arquivo poligonal (.ply). Em alguns casos, o desgaste dentário remove as pontas dos cornos da dentina. No caso de amostras com desgaste mínimo, a porção faltante do corno dentinário foi reconstruída seguindo procedimento publicado anteriormente 91. This methodology is similar to correcting for interstitial wear and involves inferring the shape of the dentine horn tip from the preserved anatomy of the dentine horn. This was restricted to cases in which the wear was less than wear level 3 as defined by Molnar92, e o observador estava confiante na posição original do corno dentinário usando sua experiência, conhecimento anatômico e morfologia preservada da JED. O procedimento de reconstrução foi concluído usando as ferramentas de modificação de superfície no Geomagic Studio 2014 ( https://www.geomagic.com ) e ajustando manualmente a posição dos pontos de referência colocados nas pontas dos cornos dentinários no Avizo 6.3. Reconstruções de cornos dentinários em espécimes de H. habilis estão disponíveis na Figura 6 Complementar .
Coleta de marcos e derivação de conjuntos de marcos homólogos
Marcos 3D foram coletados no Avizo 6.3 em três conjuntos distintos: 'EDJ principal', 'EDJ ridge' e 'CEJ ridge' (Fig. 2 ). EDJ principal consiste em pontos de referência fixos colocados nas pontas das cúspides principais (Pré-molares inferiores - 1) protoconídeo 2) metaconídeo; Molares inferiores — 1) protoconídeo, 2) metaconídeo, 3) entoconídeo, 4) hipoconídeo; Pré-molares superiores – 1) protocone, 2) metacone; Molares superiores - 1) protocone, 2) paracone, 3) metacone, 4) hipocone). A crista EDJ consiste em pontos de referência deslizantes colocados ao longo das cristas marginais que circundam o dente, seguindo a mesma ordem dos pontos de referência principais EDJ. Ambos os conjuntos de EDJ foram coletados diretamente nos modelos de superfície de EDJ, mas em amostras com baixo contraste entre os tipos de tecido, os pontos de referência de EDJ foram verificados ou, em alguns casos, colocados diretamente na pilha não filtrada para garantir que estivessem situados corretamente ao longo das cristas marginais . No conjunto do rebordo JCE, os pontos de referência foram colocados ao longo da junção amelo-cementária (JCE), começando no meio da face vestibular do dente (pré-molares e molares superiores) ou no canto mésio-vestibular da coroa, abaixo do protoconídeo (pré-molares molares) e continuando mesialmente. Esses pontos de referência foram colocados em uma renderização de isosuperfície da pilha TIFF não filtrada ou, nos casos em que a JCE não é visível na isosuperfície devido ao acúmulo de cálculo ou à presença de um dente adjacente, os pontos de referência foram colocados diretamente na pilha não filtrada.
Uma função spline cúbica foi usada para ajustar uma curva suave através de ambos os conjuntos de pontos de referência da crista, e os pontos de referência principais do EDJ foram projetados na curva da crista do EDJ para dividi-la em seções (duas seções em pré-molares, quatro em molares). Um número fixo de pontos de referência espaçados equidistantemente foram colocados em cada seção das curvas (nos pré-molares, 20 pontos de referência foram colocados na primeira seção da crista EDJ e 25 na segunda, enquanto nos molares, cada seção da crista EDJ tem 20 pontos de referência. O CEJ conjunto de crista tem 40 pontos de referência em todas as posições dos dentes). Marcos homólogos foram então derivados em R usando os pacotes Morpho 93 and princurve94 using a freely available R-based software routine95, and following previously published protocols for premolars42,43 and molars49,96. Os principais marcos do EDJ foram fixados, enquanto aqueles nos conjuntos de cumeeira EDJ e cumeeira CEJ foram tratados como semimarcos e deixados deslizar ao longo de suas curvas para reduzir a energia de flexão da função de interpolação spline de placa fina calculada entre cada amostra e a média de Procrustes para a amostra 97 . O deslizamento foi realizado duas vezes, com pontos de referência projetados de volta na curva após cada etapa, antes que os pontos de referência fossem considerados geometricamente homólogos e convertidos em coordenadas de forma usando a sobreposição de Procrustes de mínimos quadrados generalizados, que remove informações de escala, localização e orientação das coordenadas 98 , 99 , 100 , 101 .
Para incisivos e caninos, o mesmo processo foi seguido, mas com pontos de referência colocados apenas ao redor da JAC. Os pontos de referência foram colocados começando no ponto médio da face vestibular e continuando mesialmente, e como em outras posições dos dentes, 40 pontos de referência equidistantes da JAC foram derivados, antes do deslizamento.
Análise da forma e tamanho do EDJ
A análise de componentes principais (PCA) foi realizada utilizando as coordenadas de Procrustes de cada amostra no espaço de forma. Isso foi concluído separadamente para cada posição do dente pós-canino inferior e pós-canino superior usando os conjuntos de pontos de referência EDJ e CEJ. Para os dentes anteriores, o mesmo foi feito utilizando apenas os conjuntos de referência CEJ. Em todos os casos, os PCAs foram usados como um passo inicial para explorar a variação na forma do EDJ e para identificar características do EDJ que distinguem entre o Australopithecus posterior e o Homo . Essas mudanças de forma foram visualizadas usando imagens de estrutura de arame da mudança de forma do EDJ nos componentes principais. Os espécimes de H. habilis foram então avaliados de acordo com sua posição nas parcelas de PCA, bem como avaliando manualmente a expressão de características de EDJ que foram identificadas como importantes (por exemplo, aquelas que distinguem o Australopithecus posterior do Homo ).
A análise de variáveis canônicas (CVA) foi realizada nas amostras de Australopithecus , H. habilis e H. erectus , utilizando um pequeno subconjunto de componentes principais (entre 5 e 10) para garantir que o número de espécimes excedesse o número de variáveis. Em cada caso, o número de PCs com a maior precisão de classificação com validação cruzada foi selecionado para uso em figuras, e imagens de wireframe foram produzidas para visualizar a mudança na forma do EDJ entre os CVs. CVAs com validação cruzada (cvCVAs) também foram produzidos para verificar se há separação espúria de grupos. As afinidades de grupo de espécimes sem atribuição específica foram avaliadas projetando-os em CVAs antes do cálculo das probabilidades de tipicidade; isso foi repetido para cada subconjunto de PCs (5-10, resultando em 6 AVCs) para garantir que a escolha dos subconjuntos de PCs não influenciasse os resultados. As afinidades de grupo foram avaliadas utilizando um limite conservador de 0,1; se as probabilidades de tipicidade para todos os três grupos estivessem abaixo deste limite, o espécime era considerado não classificado.
O tamanho dos espécimes foi analisado utilizando o logaritmo natural do tamanho do centróide em duas análises separadas, uma para dentes inferiores e outra para dentes superiores. Apenas os conjuntos de pontos de referência CEJ foram utilizados para estas análises para permitir comparações de padrões de tamanho em toda a fileira dentária.
Duas análises foram realizadas para avaliar os níveis de variação esperados dentro de uma espécie. No primeiro, foram calculadas distâncias de Procrustes entre todos os pares possíveis de indivíduos dentro de cada uma das espécies existentes ( P. troglodytes, G. gorilla, H. sapiens ), bem como entre cada par possível entre grupos para Pan -Gorila e Pan. —H . sapiens . Estas foram plotadas como distribuições de frequência para visualizar a variação inter e intragrupo e foram comparadas com as diferenças observadas entre os espécimes de H. habilis (especificamente, OH 16 vs outro H. habilis ). Isso foi feito separadamente para cada posição dentária pós-canina (exceto para os terceiros molares). Para a segunda análise, é considerado um aspecto específico da forma do EDJ; altura do corpo da dentina. A altura do corpo da dentina refere-se à distância entre o colo do útero e a bacia oclusal do dente (equivalente à altura total da coroa menos a altura dos cornos da dentina, também conhecida como altura cervical 102 ). Nos pré-molares, sabe-se que um corpo de dentina relativamente mais alto distingue o Homo posterior do Australopithecus 42 , 43 , e em humanos modernos está fortemente correlacionado com o tamanho do dente. A altura do corpo da dentina foi quantificada aqui tomando o centróide de um subconjunto de pontos de referência nas cristas marginais mesial e distal de cada dente, bem como o centróide de um subconjunto correspondente de pontos de referência nos lados mesial e distal do colo do útero, calculando então o distância entre os dois. Os dois subconjuntos de pontos de referência foram escolhidos separadamente para cada posição dentária, a fim de capturar a porção mais baixa das cristas marginais, entre os cornos dentinários, nesses espécimes. Nos molares inferiores, o hipoconulídeo frequentemente ocupa uma posição distal na coroa, então, em vez disso, foram utilizados dois subconjuntos distais, um de cada lado do hipoconulídeo. Em cada caso, o mesmo número de pontos de referência foi utilizado nos subconjuntos mesial e distal. Isto foi concluído após a sobreposição de Procrustes e, como tal, descreve a altura do corpo da dentina em relação ao formato geral da coroa, e não como uma medida absoluta. As diferenças dentro e entre grupos na altura do corpo da dentina foram então calculadas e visualizadas da mesma maneira descrita acima para as distâncias de Procrustes.
Também testamos as diferenças de forma e tamanho entre os grupos usando testes de permutação, realizados em R usando 10.000 permutações, separadamente para cada posição do dente pós-canino. Para a forma, isso foi feito usando as coordenadas de Procrustes da análise EDJ completa, e para o tamanho, isso foi feito usando o tamanho do centróide da análise CEJ para permitir a inclusão dos dentes anteriores. O procedimento de Benjamini-Hochberg foi usado para controlar a taxa de falsas descobertas 103 .
Resumo do relatório
Mais informações sobre o desenho da pesquisa estão disponíveis no Nature Portfolio Reporting Summary vinculado a este artigo.
Disponibilidade de dados
Os dados de marcos morfométricos geométricos gerados neste estudo foram depositados na seção Publicações do The Human Fossil Record ( https://human-fossil-record.org/index.php?/category/14230 ). Os dados brutos de digitalização usados neste estudo são curados pelos museus e institutos que fazem a curadoria do material fóssil original. Esses dados foram usados sob um MOU para o estudo atual e, portanto, não estão disponíveis publicamente, mas podem ser acessados por meio de solicitação de pesquisa à instituição curatorial relevante (ver Dados Suplementares 21 ). Os dados de origem para as Figs. 3 , 4 , 5 e Figs. Suplementares. 7 , 10 , 11 , 12 , 13 , 14 , 15 e 16 são fornecidos como um arquivo de dados de origem. Os dados de origem da Figura 6 e da Figura Complementar 17 estão disponíveis em Dados Suplementares 21 . As Tabelas Suplementares 1 e 2 são baseadas em dados publicados anteriormente, que estão disponíveis nas referências listadas na Nota Complementar 1 . Os dados de origem são fornecidos neste artigo.
Code availability
R code used for geometric morphometric analyses is available from: https://zenodo.org/doi/10.5281/zenodo.1025528894.
References
Leakey, L. S. B., Tobias, P. V. & Napier, J. R. A new species of the genus Homo from Olduvai Gorge. Nature 202, 7–9 (1964).
Boaz, NT & Howell, FC Um crânio de hominídeo grácil do membro superior G da Formação Shungura, Etiópia. Sou. J. Física. Antropol. 46 , 93–108 (1977).
Hughes, AR & Tobias, PV Um crânio fóssil provavelmente do gênero Homo de Sterkfontein, Transvaal. Natureza 265 , 310–312 (1977).
Coppens, Y. em Current Argument on Early Man, Relatório de um simpósio Nobel 207–225 (Pergamon Press, 1980).
Wood, BA Projeto de Pesquisa Koobi Fora: Volume 4. Restos Cranianos de Hominídeos . Projeto de Pesquisa Koobi Fora (Clarendon Press, 1991).
Grine, FE, Demes, B., Jungers, WL & Cole, TM III Afinidade taxonômica do crânio Homo inicial de Swartkrans, África do Sul. Sou. J. Física. Antropol. 92 , 411–426 (1993).
Suwa, G., White, T. D. & Howell, F. C. Mandibular postcanine dentition from the Shungura Formation, Ethiopia: crown morphology, taxonomic allocations, and Plio-Pleistocene hominid evolution. Am. J. Phys. Anthropol. 101, 247–282 (1996).
Wood, B. & Boyle, E. K. Hominin taxic diversity: fact or fantasy? Am. J. Phys. Anthropol. 159, 37–78 (2016).
Haile-Selassie, Y., Melillo, SM & Su, DF O enigma da diversidade dos hominídeos do Plioceno: Mais fósseis significam menos clareza? Processo. Acad. Nacional. Ciência. EUA 113 , 6364–6371 (2016).
Leakey, MG et al. Novo gênero de hominídeos da África Oriental mostra diversas linhagens do Plioceno Médio. Natureza 410 , 433–440 (2001).
Haile-Selassie, Y. et al. Novas espécies da Etiópia expandem ainda mais a diversidade de hominídeos do Plioceno Médio. Natureza 521 , 483–488 (2015).
Brunet, M. et al. Australopithecus bahrelghazali , uma nova espécie de antigo hominídeo da região de Koro Toro (Chade). Relatórios de. a Academia de. Ciência. Série 2. Ciência. de. a terra e. do. planetas 322, 907–913 (Comptes Rendus de. l'Académie des. Sci., 1996).
Tobias, P. V. The distinctiveness of Homo habilis. Nature 209, 953–957 (1966).
Tobias, P. V. Early man in East Africa: recent excavations in Olduvai Gorge, Tanzania, have laid bare a new chapter in human evolution. Science 149, 22–33 (1965).
Antón, S. C. & Middleton, E. R. Making meaning from fragmentary fossils: early Homo in the Early to early Middle Pleistocene. J. Hum. Evol. 179, 103307 (2023).
Kimbel, WH & Villmoare, B. Do Australopithecus ao Homo : a transição que não existiu. Filós. Trad. R.Soc. B: Biol. Ciência. 371 , 20150248 (2016).
Lieberman, DE, Pilbeam, DR & Wood, BA Uma abordagem probabilística para o problema do dimorfismo sexual no Homo habilis : uma comparação de KNM-ER 1470 e KNM-ER 1813. J. Hum. Evol. 17 , 503–511 (1988).
Groves, CP Uma Teoria da Evolução Humana e Primata . (Clarendon Press, 1991).
Wood, B. Origem e evolução do gênero Homo . Natureza 355 , 783–790 (1992).
Rightmire, GP Variação entre os primeiros Homo crania do desfiladeiro de Olduvai e da região de Koobi Fora. Sou. J. Física. Antropol. 90 , 1–33 (1993).
Leakey, MG et al. Novos fósseis de Koobi Fora, no norte do Quênia, confirmam a diversidade taxonômica precocemente. Homo 488 , 201 (2012).
Spoor, F. et al. tipo OH 7 reconstruído O Homo habilis sugere uma diversidade de espécies profundamente enraizada no Homo primitivo. Natureza 519 , 83 (2015).
Suwa, G. Uma análise comparativa de restos dentais de hominídeos das formações Shungura e Usno . Omo Valley, Etiópia Tese de doutorado (1990).
Schrenk, F., Bromage, TG, Betzler, CG, Ring, U. & Juwayeyi, YM Homo mais antigo e biogeografia do Plioceno da fenda do Malawi. Natureza 365 , 833–836 (1993).
Kimbel, WH, Johanson, DC & Rak, Y. Avaliação sistemática de uma maxila de Homo de Hadar, Etiópia. Sou. J. Física. Antropol. 103 , 235–262 (1997).
Prat, S. et al. Primeira ocorrência do Homo inicial na formação Nachukui (West Turkana, Quênia) em 2,3-2,4 milhões de anos. J. Hum. Evol. 49 , 230–240 (2005).
Grine, FE et al. Dentição mandibular permanente completa do Homo inicial do Membro Burgi superior da Formação Koobi Fora, Ileret, Quênia. J. Hum. Evol. 131 , 152–175 (2019).
Herries, AI et al. Contemporaneidade do Australopithecus , Paranthropus e dos primeiros Homo erectus na África do Sul. Ciência 368 , eaaw7293 (2020).
Villmoare, B. et al. Early Homo at 2.8 Ma from Ledi-Geraru, Afar, Ethiopia. Science 347, 1352–1355 (2015).
Blumenschine, R. J. et al. Late Pliocene Homo and hominid land use from western Olduvai Gorge, Tanzania. Science 299, 1217–1221 (2003).
Clarke, RJ Um Homo habilis maxilla e outros fósseis de hominídeos recém-descobertos no desfiladeiro de Olduvai, na Tanzânia. J. Hum. Evol. 63 , 418–428 (2012).
Clarke, RJ e Kuman, K. O crânio de StW 573, um Australopithecus prometheus esqueleto de de 3,67 ma das cavernas de Sterkfontein, África do Sul. J. Hum. Evol. 134 , 102634 (2019).
Wood, B. & Collard, M. O gênero humano. Ciência 284 , 65–71 (1999).
Lieberman, DE Outro rosto em nossa árvore genealógica. Natureza 410 , 419–420 (2001).
Cameron, D. W. & Groves, C. P. Bones, Stones and Molecules:” Out of Africa” and human origins. (Academic Press, 2004).
Collard, M. & Wood, B. Defining the Genus Homo. Handbook of Paleoanthropology. 2107–2144 (Springer Science & Business Media, 2015).
Mongle, C. S., Strait, D. S. & Grine, F. E. An updated analysis of hominin phylogeny with an emphasis on re-evaluating the phylogenetic relationships of Australopithecus sediba. J. Hum. Evol. 175, 103311 (2023).
Dembo, M. et al. As relações evolutivas e a idade do Homo naledi : uma avaliação usando métodos filogenéticos bayesianos datados. J. Hum. Evol. 97 , 17–26 (2016).
Caparros, M. & Prat, S. Uma perspectiva de redes filogenéticas na evolução humana reticulada. Ciência 24 , 102359 (2021).
White, TD, Johanson, DC & Kimbel, WH em Novas Interpretações de Macacos e Ancestrais Humanos (eds. Ciochon, RL & Corruccini, RS) 721–780 (Springer, 1983).
Tobias, PV Os Crânios, Endocasts e Dentes do Homo habilis : pt. 1-4 . (Cambridge University Press, 1991).
Davies, TW, Delezene, LK, Gunz, P., Hublin, JJ & Skinner, MM Morfologia endoestrutural em terceiros pré-molares inferiores hominóides: análise morfométrica geométrica do formato da coroa dentinária. J. Hum. Evol. 133 , 198–213 (2019).
Davies, T. W. et al. Distinct mandibular premolar crown morphology in Homo naledi and its implications for the evolution of Homo species in southern Africa. Sci. Rep. 10, 1–13 (2020).
Olejniczak, AJ, Martin, LB & Ulhaas, L. Quantificação da forma da dentina em primatas antropóides. Ana. Anat. Anatomista Anz. 186 , 479–485 (2004).
Olejniczak, AJ et al. Morfologia da junção esmalte-dentina em cortes de molares superiores de primatas antropóides. J. Hum. Evol. 53 , 292–301 (2007).
Skinner, MM, Gunz, P., Wood, BA, Boesch, C. & Hublin, J.-J. Discriminação de existentes de Pan espécies e subespécies usando a morfologia da junção esmalte-dentina dos molares inferiores. Sou. J. Física. Antropol. 140 , 234–243 (2009).
Skinner, MM, Gunz, P., Wood, BA e Hublin, J.-J. A morfologia da junção esmalte-dentina (EDJ) distingue os molares inferiores de Australopithecus africanus e Paranthropus robustus . J. Hum. Evol. 55 , 979–988 (2008).
Pan, L. et al. Outras evidências morfológicas sobre a dentição pós-canina inferior do Homo mais antigo da África do Sul : espessura do esmalte e junção esmalte-dentina. J. Hum. Evol. 96 , 82–96 (2016).
Martin, R. M., Hublin, J.-J., Gunz, P. & Skinner, M. M. The morphology of the enamel–dentine junction in Neanderthal molars: Gross morphology, non-metric traits, and temporal trends. J. Hum. Evol. 103, 20–44 (2017).
Zanolli, C. et al. Evidence for increased hominid diversity in the Early to Middle Pleistocene of Indonesia. Nat. Ecol. Evol. 3, 755–764 (2019).
Pan, L. et al. Morfometria geométrica da interface esmalte-dentina de quartos pré-molares inferiores de Lufengpithecus hudienensis . Acta Antropol. Pecado. 39 , 555 (2020).
Zanolli, C. et al. Os dados dentários desafiam a presença onipresente do Homo no Berço da Humanidade. Processo. Acad. Nacional. Ciência. EUA 119 , e2111212119 (2022).
Dahlberg, AA A mudança na dentição do homem. Geléia. Dente. Assoc. 32 , 676–690 (1945).
Butler, PM Morfologia dentária e evolução dos primatas. Antropol Odontológico. 1963 , 1–13 (1963).
Quam, R., Bailey, S. & Wood, B. Evolução do tamanho da coroa M1 e proporções das cúspides no gênero Homo . J. Anat. 214 , 655–670 (2009).
Bailey, S. E. A morphometric analysis of maxillary molar crowns of Middle-Late Pleistocene hominins. J. Hum. Evol. 47, 183–198 (2004).
Gómez-Robles, A., Bermúdez de Castro, J. M., Martinón-Torres, M., Prado-Simón, L. & Arsuaga, J. L. A geometric morphometric analysis of hominin upper second and third molars, with particular emphasis on European Pleistocene populations. J. Hum. Evol. 63, 512–526 (2012).
Gómez-Robles, A. et al. Uma análise morfométrica geométrica da forma do primeiro molar superior dos hominídeos. J. Hum. Evol. 53 , 272–285 (2007).
Wood, B. em Espécies, Conceitos de Espécies e Evolução de Primatas. (eds. Kimbel, WH & Martin, LB) 485–522 (Springer, 1993).
Davies, TW et al. Expressão de cúspide acessória na junção esmalte-dentina de molares inferiores hominídeos. PeerJ 9 , e11415 (2021).
Bruner, E. & Beaudet, A. O cérebro do Homo habilis : Três décadas de paleoneurologia. J. Hum. Evol. 174 , 103281 (2023).
Lieberman, D. E., Wood, B. A. & Pilbeam, D. R. Homoplasy and early Homo: an analysis of the evolutionary relationships of H. habilis sensu stricto and H. rudolfensis. J. Hum. Evol. 30, 97–120 (1996).
Irish, J. D., Guatelli-Steinberg, D., Legge, S. S., de Ruiter, D. J. & Berger, L. R. Dental morphology and the phylogenetic “place” of Australopithecus sediba. Science 340, 1233062 (2013).
Napier, J. Fossil hand bones from Olduvai Gorge. Nature 196, 409–411 (1962).
Susman, RL & Creel, N. Afinidades funcionais e morfológicas da mão subadulta (OH 7) de Olduvai Gorge. Sou. J. Física. Antropol. 51 , 311–331 (1979).
Johanson, DC et al. Novo esqueleto parcial do Homo habilis do desfiladeiro de Olduvai, Tanzânia. Natureza 327 , 205–209 (1987).
Brasil, MF, Monson, TA, Schmitt, CA & Hlusko, LJ Um genótipo: abordagem fenotípica para testar hipóteses taxonômicas em hominídeos. Ciência. Nat. 107 , 1–16 (2020).
Grine, FE A dentição decídua do Kalahari San, do negro sul-africano e dos hominídeos sul-africanos do Plio-Pleistoceno. (Dissertação de doutorado, Universidade de Witwatersrand. 1984).
Deino, AL 40Ar/39Ar datação do leito I, desfiladeiro de Olduvai, Tanzânia, e a cronologia das mudanças climáticas do início do Pleistoceno. J. Hum. Evol. 63 , 251–273 (2012).
Joordens, JC et al. Melhor controle de idade em fósseis de Homo primitivos do membro Burgi superior em Koobi Fora, Quênia. J. Hum. Evol. 65 , 731–745 (2013).
Feibel, CS, Lepre, CJ & Quinn, RL Estratigrafia, correlação e estimativas de idade para fósseis da Área 123, Koobi Fora. J. Hum. Evol. 57 , 112–122 (2009).
Leakey, MD Olduvai Gorge: Volume 3, Escavações nos Leitos I e II, 1960-1963 . 3 (Cambridge University Press, 1971).
Spoor, F. et al. Implicações de novos fósseis de Homo primitivos de Ileret, a leste do Lago Turkana, Quênia. Natureza 448 , 688–691 (2007).
Robinson, JT Homo 'habilis' e os Australopitecos. Natureza 205 , 121–124 (1965).
Simons, EL, Pilbeam, D. & Ettel, PC Taxonomia controversa de hominídeos fósseis. Ciência. 166 , 258–259 (1969).
Tobias, P. V. & von Koenigswald, G. H. R. A comparison between the Olduvai hominines and those of Java and some implications for hominid phylogeny. Curr. Anthropol. 6, 427–431 (1965).
Tobias, PV et al. Novas descobertas em Tanganica: sua influência na evolução dos hominídeos [e comentários e respostas]. Curr. Antropol . 6 , 391–411 (1965).
Leakey, LS Homo habilis , Homo erectus e os Australopitecos. Natureza 209 , 1279–1281 (1966).
Antón, SC Early Homo : quem, quando e onde. Curr. Antropol. 53 , S278–S298 (2012).
Jiménez-Arenas, JM Variabilidade do tamanho canino em táxons de hominídeos extintos, com ênfase especial no Homo habilis : implicações paleobiológicas e taxonômicas. Período. J. Paleontol. 27 , 1–14 (2020).
Prat, S. Surgimento do gênero Homo : Do conceito à taxonomia. L'Anthropologie 126 , 103068 (2022).
Schwartz, JH & Tattersall, I. Definindo o gênero Homo . Ciência 349 , 931–932 (2015).
Leonard, WR & Hegmon, M. Evolução da morfologia P3 em Australopithecus afarensis . Sou. J. Física. Antropol. 73 , 41–63 (1987).
Suwa, G. et al. Early Pleistocene Homo erectus fossils from Konso, southern Ethiopia. Anthropol. Sci. 115, 133–151 (2007).
Rightmire, G. P. Middle Pleistocene hominids from Olduvai Gorge, northern Tanzania. Am. J. Phys. Anthropol. 53, 225–241 (1980).
Leakey, MD Descobertas recentes de restos de hominídeos em Olduvai Gorge, Tanzânia. Natureza 223 , 756–756 (1969).
Deino, AL et al. Cronoestratigrafia e modelagem de idade de testemunhos de perfuração do Pleistoceno da bacia de Olduvai, Tanzânia (projeto de testemunho Olduvai Gorge). Paleogeogr. Paleoclimatol. Paleoecol. 571 , 109990 (2021).
Leakey, M. & Roe, D. Olduvai Gorge: Volume 5, Escavações nos leitos III, IV e nos leitos Masek . 5 (Cambridge University Press, 1995).
Zanolli, C. et al. Quando os raios X não funcionam. caracterizando a estrutura interna de restos dentognáticos de hominídeos fósseis usando imagens microtomográficas de nêutrons de alta resolução. Frente. Eco. Evol. 8 , 42 (2020).
Wollny, G. et al. MIA-A free and open source software for gray scale medical image analysis. Source Code Biol. Med. 8, 20 (2013).
Skinner, M. Enamel-Dentine Junction Morphology of Extant Hominoid and Fossil Hominin Lower Molars. (The George Washington University, 2008).
Molnar, S. Human tooth wear, tooth function and cultural variability. Am. J. Phys. Anthropol. 34, 175–189 (1971).
Schlager, S. in Statistical Shape and Deformation analysis. 217–256 (Elsevier, 2017).
Cannoodt, R. & Bengtsson, H. rcannood/princurve: princurve 2.1.4. https://doi.org/10.5281/zenodo.3351282 (2019).
Davies, TW tomdavies15/EDJ_GM: Análise morfométrica geométrica da morfologia da junção esmalte-dentina. https://doi.org/10.5281/zenodo.10255289 (2023).
Skinner, MM et al. Uma perspectiva dentária sobre a afinidade taxonômica da mandíbula Balanica (BH-1). J. Hum. Evol. 93 , 63–81 (2016).
Gunz, P. & Mitteroecker, P. Semilandmarks: um método para quantificar curvas e superfícies. Hystrix-Ital. J. Mamífero. 24 , 103–109 (2013).
Gower, JC Análise generalizada de procustos. Psicometria 40 , 33–51 (1975).
Rohlf, FJ & Slice, D. Extensões do método Procrustes para a sobreposição ideal de pontos de referência. Sist. Biol. 39 , 40–59 (1990).
Goodall, C. Procrustes methods in the statistical-analysis of shape. J. R. Stat. Soc. Ser. B-Methodol. 53, 285–339 (1991).
Dryden, IL e Mardia, KV Análise Estatística de Forma (Wiley, 1998).
Monson, TA, Fecker, D. & Scherrer, M. Evolução neutra da morfologia da junção esmalte-dentina humana. Processo. Acad. Nacional. Ciência. EUA 117 , 26183–26189 (2020).
Benjamini, Y. & Hochberg, Y. Controlando a taxa de falsas descobertas: uma abordagem prática e poderosa para testes múltiplos. Estado JR. Soc. Ser. Método B. 57 , 289–300 (1995).
Reconhecimentos
Pelo acesso às amostras e microtomografias, gostaríamos de agradecer a Emma Mbua e Samuel Muteti (Museus Nacionais do Quénia), Audax Mabulla (Museu Nacional da Tanzânia), a Comissão de Ciência e Tecnologia da Tanzânia, Metasebia Endalemaw, Yared Assefa (Autoridade Etíope para Pesquisa e Conservação do Patrimônio Cultural), Bernhard Zipfel, Sifelani Jira (Instituto de Estudos Evolutivos, Universidade de Witwatersrand), Miriam Tawane (Museu Ditsong), Davorka Radovčić (Museu Croata de História Natural), Michel Toussaint (ASBL Archéologie Andennaise, Royal Belgian Instituto de Ciências Naturais), Jean-Jacques Cleyet-Merle (Musée National de Préhistoire des Eyzies-de-Tayac), Véronique Merlin-Langlade (Musée d'Art et d'Archéologie du Périgord), Christine Feja (Institut für Anatomie der Universität Leipzig), Antonio Rosas (Museu Nacional de Ciências Naturais), Flora Gröning (Instituto de Ciências Médicas, Universidade de Aberdeen), Dejana Brajković (Academia Croata de Ciências e Artes), Jean-François Tournepiche (Musée d'Angoulême), Patrick Périn (Musée d'Archéologie nationale de Saint-Germain-en-Laye), Paul Tafforeau (Instalação Europeia de Radiação Síncrotron), David Hunt (Museu Nacional de História Natural), Ullrich Glasmacher (Instituto für Geowissenschaften, Universität Heidelberg), Mirjana Roksandic (Universidade de Winnipeg), Dušan Mihailović, Bojana Mihailović (Departamento de Arqueologia, Universidade de Belgrado e Museu Nacional), Eleni Panagopoulou (Eforato de Paleoantropologia e Espeleologia do Sul da Grécia), Katarina Harvati (Universidade de Tübingen), Martin Fischer (Museu Phyletisches Jena) , Irina Ruf, Friedemann Schrenk e Christine Hemm (Senckenberg Forschungsinstitut und Naturmuseum, Frankfurt), Frieder Mayer e Christiane Funk (Museum für Naturkunde — Instituto Leibniz para a Evolução e Ciência da Biodiversidade), Emmanuel Gilissen (Museu Real da África Central, Tervuren), Christophe Boesch e Roman Wittig (Projeto Taï Chimpanzee), e Senckenberg, Forschungsstation für Quartärpaläontologie. Expressamos nossa gratidão à Fundação Werner Reimers em Bad Homburg (Alemanha), que fornece a coleção Gustav Heinrich Ralph von Koenigswald como empréstimo permanente para pesquisa científica ao Instituto de Pesquisa Senckenberg e ao Museu de História Natural de Frankfurt. Pela varredura µCT e assistência técnica, agradecemos a Kudakwashe Jakata, David Plotzki e Heiko Temming. Agradecemos à instalação de imagem ANTARES localizada no reator FRM II da Universidade Técnica de Munique, Alemanha, e a Burkhard Schillinger pela possibilidade de varredura de nêutrons das amostras de Sangiran. Pela assistência na segmentação, agradecemos a Lia Schurtenberger, Rhianna Drummond-Clarke e Mykolas Imbrasas. Este projeto recebeu financiamento da Sociedade Max Planck (TWD, PG, J.-JH), do Conselho Europeu de Pesquisa (acordo de subvenção nº 819960; MMS) e da Fundação Calleva (Reino Unido; FS).
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.