Um provável coala do Oligoceno da Austrália central fornece informações sobre a evolução inicial dos diprotodontianos
Relatórios Científicos volume 13 , número do artigo: 14521 ( 2023 )
Abstrato
Os diprotodontianos são a ordem de marsupiais mais diversa morfológica e ecologicamente. No entanto, uma lacuna de aproximadamente 30 milhões de anos no registro fóssil de vertebrados terrestres australianos significa que a primeira metade da evolução diprotodontiana é desconhecida. Os táxons fósseis de ambos os lados desta lacuna são, portanto, críticos para reconstruir a evolução inicial da ordem. Aqui relatamos os prováveis parentes de coalas mais antigos conhecidos (Phascolarctidae), da Fauna Local Pwerte Marnte Marnte do Oligoceno tardio (Austrália central). Estes incluem espécies coevas de Madakoala e Nimiokoala , bem como um novo provável coala (?Phascolarctidae). O novo táxon, Lumakoala blackae gen. e sp. nov., era comparável em tamanho aos menores fascolarctídeos conhecidos, com estimativas de massa corporal de 2,2–2,6 kg. Seus molares superiores bunoselenodontes mantêm a condição metatérica primitiva de um centrocrista contínuo e cúspides estilares distintas B e D que não possuíam oclusão com o hipoconídeo. Este arranjo estrutural: (1) sugere uma morfoclina dentro de Phascolarctidae de bunoselenodontia a selenodontia; e (2) esclarece melhor as transições evolutivas entre morfologias molares dentro da Vombatomorphia. Nossa hipótese é que a forma molar de Lumakoala blackae se aproxima da condição ancestral da subordem Vombatiformes. Além disso, fornece uma ligação plausível entre os diprotodontianos e os supostos polidolopimórficos Chulpasia jimthorselli e Thylacotinga bartholomaii da Fauna Local de Tingamarra do início do Eoceno (leste da Austrália), que inferimos como tendo morfologias molares consistentes com os diprotodontianos do caule.
Conteúdo semelhante sendo visualizado por outras pessoas


Introdução
A Austrália é conhecida pela singularidade da sua biota, especialmente dos seus marsupiais. Este endemismo notável deve-se principalmente à sua ascendência Gondwana 1 , 2 , e longo isolamento após a separação do continente da Antártida há cerca de 45 milhões de anos (mya) 3 , 4 . Grande parte da diversidade morfológica e ecológica dos marsupiais australianos se manifesta dentro da ordem Diprotodontia, que compreende duas subordens de táxons principalmente herbívoros - Vombatiformes (wombats, coalas e parentes extintos) e Phalangerida (gambás e cangurus) - bem como Thylacoleonidae (marsupiais). leões'; Diprotodontia incertae sedis 5 ) (Figura 1 ). Análises de relógio molecular sugerem que Diprotodontia divergiu de Agreodontia (Dasyuromorphia + Peramelemorphia + Notoryctemorphia) em algum momento entre o último Cretáceo e o início do Eoceno médio 5 , 6 , 7 , 8 , 9 , 10 , 11 , 12 , 13 , 14 . Isto é importante do ponto de vista biogeográfico porque implica que os diprotodontianos divergiram antes da separação final da Austrália e da Antártida. No entanto, nenhum fóssil definitivo de diprotodontianos anteriores ao Oligoceno tardio é conhecido, com a possível exceção de tarsos semelhantes a diprotodontianos do início ou médio do Eoceno do sul da Argentina. 15 .
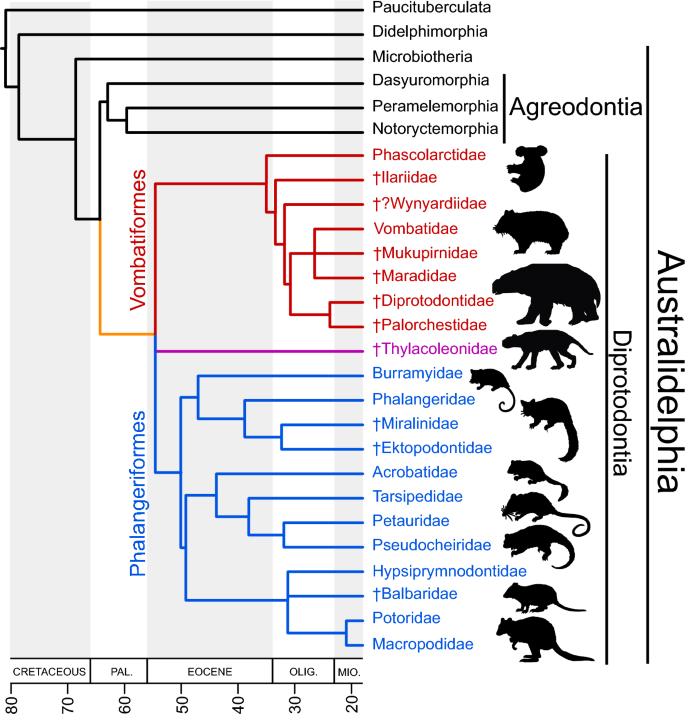
Reconstrução cladística das inter-relações diprotodontianas. Estimativas de relacionamentos e divergências para linhagens existentes seguem Duchêne et al. 6 . Aqueles de linhagens extintas seguem Beck et al. 5 , Travouillon et al. 102 e Crichton et al. 38 , 39 . As estimativas de divergência para vombatiformes e falangeróides extintos são especulativas.
Apenas dois conjuntos de fósseis australianos datados entre 110 e 25 milhões de anos produziram mamíferos 16 , 17 , deixando uma lacuna enorme que abrange praticamente toda a janela das origens dos marsupiais australianos 18 , 19 . Nesta janela, as descrições de táxons foram publicadas apenas para metatherianos (marsupiais e parentes do caule) do início do Eoceno (55 milhões de anos atrás) da Fauna Local de Tingamarra (LF) do sudeste de Queensland, todos com base em registros dentários, perióticos e tarsais isolados. espécimes 17 , 20 , 21 , 22 , 23 , 24 , 25 , 26 , 27 . Esses elementos foram interpretados como carecendo de sinapomorfias inequívocas ligando-os a táxons marsupiais de nível ordinal conhecidos no final do Oligoceno australiano. 1 , 9 , 17 , 19 , 21 . Destes táxons do início do Eoceno, apenas Djarthia murgonensis foi referido como Australidelphia (marsupiais australianos + Microbiotheria da América do Sul) 22 . Argumenta-se que outros marsupiais Tingamarra LF têm maior afinidade com polidolopimorfianos bunodontes da América do Sul 23 , 25 , 27 .
No momento em que o registro de mamíferos terrestres australianos recomeça, perto do final do Oligoceno (c. 25 milhões de anos atrás), a maioria das famílias diprotodontianas modernas estão representadas 21 , 28 , 29 . A distinção de suas morfologias dentárias e esqueléticas, bem como a disparidade molecular entre os representantes modernos, sugerem longos intervalos de divergência pré-Mioceno 5,21,30,31. Each family is characterized by a specialized molar dentition that occupies a unique region in a morphospace that includes selenodont (multiple crescents), bunodont (bulbous cusps) and bilophodont (two transverse crests) forms. Identifying homologies between these molar architectures, and reconstructing the transformations that led to their acquisition, has long been considered important to resolving the broader problem of diprotodontian interrelationships5,30,32,33,34,35,36,37,38,39,40. A radiação vombatiforme tem sido de particular interesse; os representantes existentes (coalas e wombats) capturam apenas uma pequena parte da diversidade morfológica total que se sabe ter existido.
Os coalas (Phascolarctidae) possuem molares selenodontes, caracterizados por cristas crescentes em série que funcionam como cortadores de folhas durante a mastigação (por exemplo, 41,42,43). Winge44 considered the molar form of the modern koala to be the least specialized among known diprotodontians, being only slightly modified from that of the archetypal metatherian condition. This view was accepted by Archer32, que sugeriram que a selenodontia em diprotodontistas poderia ter surgido diretamente de uma morfologia do tipo peramelemorfa (bandicoot). Isso foi baseado em grande parte na observação de que os molares modernos de coala e bandicoot não possuem uma crista contínua (centrocrista) conectando o paracone e o metacone, com o pós-paracrista e o pré-metacrista ligados a uma cúspide nas posições da cúspide estilar C e D, respectivamente. 32 . Desde então, tornou-se evidente que esta morfologia é derivada de Peramelemorphia 45 , 46 , 47 ; no entanto, a visão de que os vombatiformes fizeram a transição da selenodontia para a lofodontia foi amplamente apoiada 30 , 33 , 34 , 35 .
Este estudo relata fósseis recém-descobertos de coalas, e prováveis coalas, do depósito Pwerte Marnte Marnte, sul do Território do Norte, o provavelmente mais antigo local com mamíferos terrestres do Oligoceno no continente 29 , 48 (Figura 2 ). Descrevemos um novo gênero e espécie caracterizado por uma morfologia molar distinta que ajuda a esclarecer os padrões evolutivos dentro da Diprotodontia, e pode lançar luz sobre as afinidades filogenéticas dos enigmáticos metatherianos do início do Eoceno da Austrália. Também relatamos a presença de outras duas espécies de coalas no local.
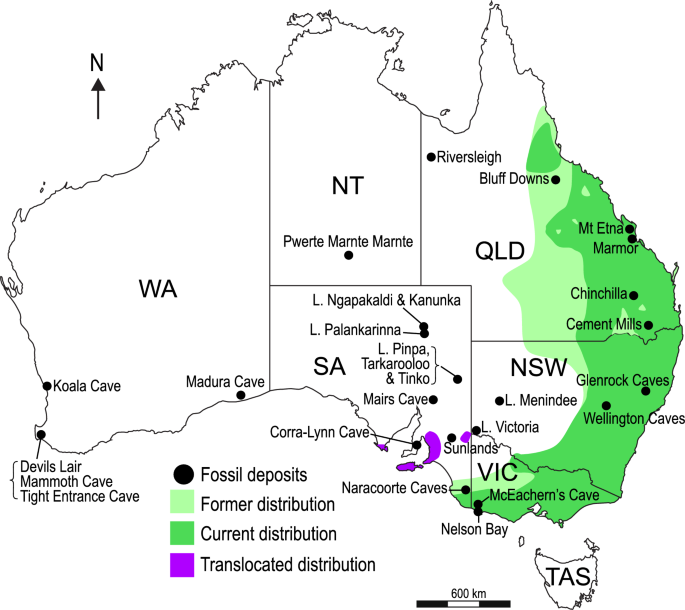
Mapa da Austrália representando ocorrências de fascolarctídeos, modificado de Price 59 e Black et al. 54 . Os sítios fósseis que produziram restos de fascolarctídeos são indicados com círculos pretos. A antiga distribuição histórica de Phascolarctos cinereus é indicada em verde claro; distribuição atual em verde escuro; e distribuição translocada em roxo.
Paleontologia sistemática
Infraclasse Marsupialia Illiger, 1811
Ordem Diprotodontia Owen, 1877
Subordem Vombatiformes Woodburne, 1984
Infraordem ?Phascolarctomorphia Aplin & Archer, 1987
Family ?Phascolarctidae Owen, 1839
Lumakoala blackae gen. et sp. nov. (Fig. 3)
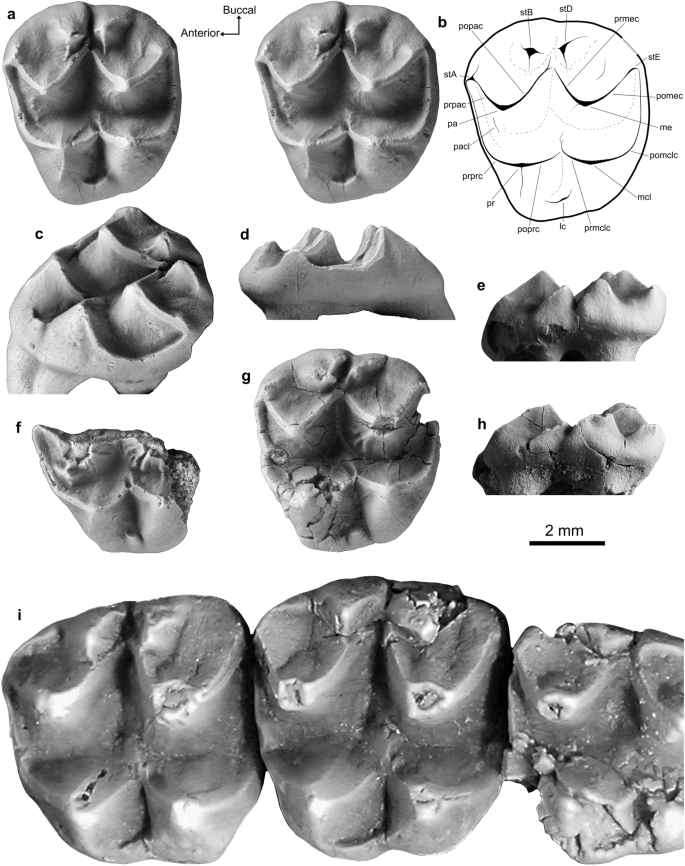
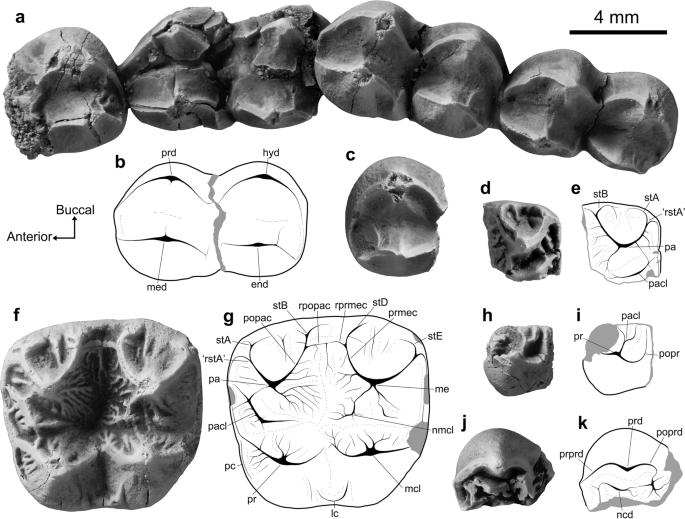
Lumakoala blackae gen. e sp. novembro. molares superiores. Esquerda M 2? ou 3 (Holotype, NTM P12012): ( a ) vista estereoclusal, com ( b ) desenho de linha anotado; ( c ) visão oclusal com deslocamento posterolingual; ( d ) vista anterior; e ( e ) vista bucal. Metade anterolingual de LM1 (Paratype, NTM P12014) em ( f ) vista oclusal. Esquerda M ?2 ou 3 (Paratype, NTM P12013) nas vistas ( g ) oclusal e ( h ) bucal. Priscakoala lucyturnbullae deixou M1 – M3 (Holotype, QM F20203), adaptado de Black et al. 58. As setas de orientação (Anterior, Bucal) referem-se apenas às amostras em vista de oclusão. lc cingulum lingual, me metacone, mcl metaconule, pa paracone, pacl paraconule, pomclc postmetaconulecrista, pomec postmetacrista, popac postparacrista, poprc postprotocrista, pr protocone, prmclc premetaconulecrista, prmec premetacrista, prpac preparacrista, prprc preprotocrista, stA stylar cúspide A, stB stylar cúspide B, cúspide estilar stD D, cúspide estilar stE E.
Holotype. NTM P12012, left M2 or M3 (Fig. 3a–e).
Parátipos . NTM P12013, M2 ou M3 esquerdo (Fig. 3 g – h); NTM P12014, metade anterolingual de M1 esquerdo (Fig. 3 f).
Localidade . Sítio fóssil Pwerte Marnte Marnte (24°21′ S 133°43′ E), derivado de uma formação geológica sem nome no flanco sul da Cordilheira James, Território do Norte, Austrália.
Fauna Local e Idade . Avaliações biocronológicas iniciais sugeriram que o Pwerte Marnte Marnte LF tem idade do final do Oligoceno 38 , 39 , 48 , e pode ser anterior às mais antigas assembleias do Oligoceno tardio das Formações Etadunna e Namba do sul da Bacia do Lago Eyre, talvez correspondendo a uma idade de mamíferos terrestres ainda sem nome imediatamente anterior ao Etadunnan 29 , 48 .
Etimologia do gênero . Luma significa 'espinho' em latim, em referência à morfologia das cúspides estilares B e D, e sua distinção da pós-paracrista e da pré-metacrista, respectivamente. O gênero do gênero é feminino.
Etimologia das Espécies . Nomeado em homenagem a Karen Black, cuja pesquisa ampliou enormemente nossa compreensão dos fósseis de fascolarctídeos e outros vombatiformes.
Diagnóstico do gênero . Quanto à espécie até que outra seja reconhecida.
Specific Diagnosis. A new species that appears most similar in upper molar morphology to members of the Phascolarctidae, with: a deep, U-shaped longitudinal valley between the primary buccal and lingual cusps, rather than being shallow and V-shaped as in selenodont vombatomorphians; and a paraconule on M1 (Fig. 3a–h).
Lumakoala blackae is relatively small compared to known phascolarctids, with upper molars that fall within the lower end of the size range of the smallest species of Litokoala. The taxon differs from all phascolarctids in having an M2 with: stylar cusps B and D that are somewhat conical and more structurally independent from the postparacrista and premetacrista, respectively; a postparacrista that continues beyond its junction with the premetacrista towards stylar cusp D; a wider stylar shelf, wherein the junction between the premetacrista and postparacrista is roughly mid-width between the buccal margin and the apices of the paracone and metacone; and more rounded buccal and posterior margins in occlusal view, leading to a generally rounder outline.
Lumakoala blackae differs from all other phascolarctids, unless otherwise noted, in having: proportionately wider molars relative to length (except Litokoala garyjohnstoni); more buccally situated protocone and metaconule apices, with a steeper buccal face of the protocone and metaconule compared with the lingual face (except L. garyjohnstoni); stylar cusp B taller than stylar cusp A on M2 (except Priscakoala lucyturnbullae); a small cuspate paraconule that extends up the anterolingual face of the paracone on M1 (except species of Phascolarctos); lacking a neometaconule (except Priscakoala lucyturnbullae); and lacking a hypertrophied anterobuccal terminus of the preprotocrista on M1 (except Priscakoala lucyturnbullae, species of Madakoala and Perikoala).
Lumakoala blackae é mais semelhante a Priscakoala lucyturnbullae, compartilhando: ornamentação simples da superfície do esmalte sem neometacônulo; cúspide estilar B mais alta que cúspide estilar A; cúspides estilares cúspides B e D; e uma ampla prateleira estilosa. Além das diferenças identificadas em relação a todos os fascolarctídeos, L. blackae difere de Priscakoala lucyturnbullae por apresentar: crenulações finas nas faces linguais do paracone e metacone; um paraconule em M1; uma cúspide estilar A que é consideravelmente maior, sendo semelhante em altura à cúspide estilar D; uma crista proeminente que desce do ápice do metacone pela face anterolingual; um cíngulo lingual curto e às vezes cúspide em vez de alongado; e um metacônulo reduzido em relação ao protocone em M2, resultando em uma porção anterior de largura marcadamente maior que a porção posterior.
Descrição . Os molares superiores de Lumakoala blackae (Fig. 3 a – h) são de coroa baixa, com crenulações finas nas faces linguais do paracone e metacone. Os molares possuem morfologia de coroa bunoselenodonte composta por quatro cúspides principais, sendo as duas cúspides anteriores representando o paracone (vestibular) e o protocone (lingual), e as duas posteriores representando o metacone (vestibular) e o metacônulo (lingual). Duas cúspides adicionais, um tanto cônicas, estão presentes, vestibulares à pós-paracrista e à pré-metacrista, que são chamadas de cúspides estilares B e D, respectivamente (ver Suplemento 1 para discussão sobre homologias de cúspides estilares). A cúspide no canto ântero-vestibular do dente, no término do preparacrista, é chamada de cúspide estilar A.
As amostras NTM P12012 e NTM P12013 são consideradas representativas de M2, ou possivelmente M3, porque: a largura do dente diminui anteroposteriormente; e há uma depressão proeminente vestíbulo-lingual extensa abrangendo as faces anterior e posterior, decorrente do contato com os molares superiores vizinhos. Com base na comparação com os molares de Priscakoala lucyturnbullae , acredita-se que NTM P12012 e NTM P12013 representem provavelmente M2, em vez de M3 porque, neste último, as cúspides estilares B e D são marcadamente reduzidas. Os espécimes atribuídos à posição M2 são trapezoidais em vista oclusal com cantos arredondados dos dentes, decorrentes de uma margem vestibular fortemente convexa e uma margem lingual que se curva póstero-vestibularmente. As proporções molares diminuem de anterior para posterior e de vestibular para lingual (Fig. 3 ). A largura anterior do NTM P12012 é maior que a largura posterior em 7% e maior que o comprimento em 12% ( Tabela S1 ). O corpo de prova NTM P12013 está completamente fraturado e, consequentemente, possui dimensões ligeiramente infladas ( Tabela S1 ). Na NTM P12013, também há danos na pós-protocrista, no ápice do metacone e no canto póstero-vestibular do dente. O maior desgaste está presente ao longo das bordas das cristas. A descrição da morfologia M2 baseia-se principalmente no holótipo NTM P12012, devido à sua melhor preservação.
O metacone é a maior cúspide e o metacônulo é o menor (Fig. 3 ). As bacias bucais do paracone e do metacone são apenas ligeiramente côncavas à medida que se inclinam dos ápices para encontrar os limites das cúspides estilares. A margem vestibular é aberta entre cada uma das (1) cúspides estilares A e B, e (2) cúspide estilar D e o término da pós-metacrista (Fig. 3 ). O ápice do paracone está posicionado ligeiramente anterior em relação ao ápice do protocone e ligeiramente vestibular em relação ao ápice do metacone. Uma preparacrista linear desce ântero-vestibularmente até uma estrutura cúspide ligeiramente elevada no canto ântero-vestibular do dente, representando a cúspide estilar A. Duas cristas descem da cúspide estilar A: uma póstero-vestibular, terminando anterior à cúspide estilar B; e o outro anterolingualmente, que é então contínuo com o pré-protocrista. Este último não é hipertrofiado anteriormente em uma estrutura cúspide (referido como parastilo em Black et al. 49 ). Uma pós-paracrista linear desce póstero-bucalmente, encontrando a pré-metacrista próxima à extremidade lingual das cúspides estilares B e D. A pós-paracrista continua fracamente além de sua junção com a pré-metacrista em direção à cúspide estilar D antes de terminar. Na NTM P12013, a pós-paracrista é contínua com uma crista descendo anterolingualmente da cúspide estilar D, que, juntas, fecham a cúspide estilar B. A pós-paracrista e a pré-metacrista são estruturalmente independentes das cúspides estilares B e D, respectivamente.
As cúspides estilares B e D são um tanto cônicas; a cúspide estilar B é a mais alta, sendo apenas c. 20% mais curto que o paracone (Fig. 3 e – h). Várias cristas curtas descem dos ápices das cúspides estilares B e D, expressas diferencialmente entre NTM P12012 e NTM P12013 (Fig. 3 ). Na NTM P12012, a crista mais proeminente que desce do ápice da cúspide estilar B é orientada anterolingualmente, fracamente paralela à pós-paracrista. Da cúspide estilar D, uma crista é orientada posterolingualmente, paralela à pré-metacrista, e uma segunda é orientada posteriormente. Em comparação, no NTM P12013, a crista mais proeminente que desce da cúspide estilar B é orientada póstero-vestibularmente; e da cúspide estilar D, uma é orientada anterolingualmente e outra póstero-vestibular, formando juntas uma crista. Nenhuma estrutura liga as cúspides estilares. No NTM P12012, uma crista tênue desce pela bacia bucal do metacônio anterior ao pós-metacrista. O pós-metacrista desce póstero-vestibularmente do ápice do metacone, em direção ao canto póstero-vestibular do dente, onde então se arqueia ventralmente, tornando-se contínuo com o pós-metaconulecrista. Há um ligeiro inchaço no terminal póstero-vestibular da pós-metacrista, que representa a cúspide estilar E. O paracone e o metacone têm perfil um tanto cônico, em que as faces lingual e bucal são igualmente inclinadas. Uma leve crista desce do ápice do metacone pela face anterolingual, terminando antes da bacia longitudinal. No NTM P12013, a crista é mais proeminente e se bifurca no meio da face anterolingual do metacone.
O protocone está posicionado um pouco mais posteriormente que o paracone e um pouco mais lingual que o metacônulo (Fig. 3 ). Um pré-protocrista crescente desce ântero-vestibularmente do ápice do protocone, curvando-se à medida que atravessa a margem anterior do dente. Nos espécimes NTM P12012 e NTM P12013, uma crista tênue ascende pela face anterolingual do paracone, representando um paraconule incipiente. Na NTM P12013, há também uma crista curta posterolingual ao ápice do paracone no vale longitudinal que forma um leve inchaço ao interceptar a pré-protocrista (Fig. 3 ). Esta estrutura é considerada um protostilo, cuja ausência na NTM P12012 pode ser uma variação intraespecífica. Uma pós-protocrista linear desce posteriormente, interceptando a pré-metaconulecrista no vale transversal. O pré-metaconulecrista continua além de sua junção com o pós-protocrista antes de terminar. O pós-protocrista e o premetaconulecrista são, cada um, apenas ligeiramente inclinados para vestibular, formando juntos um ângulo lingual relativamente amplo de 115 graus, em comparação com o ângulo de 75 graus formado pelo pós-paracrista e pelo pré-metacrista. O ápice do metacônulo é ligeiramente posterior ao ápice do metacônio. Um pós-metaconulecrista crescente curva-se póstero-vestibularmente ao longo da margem posterior do dente. As faces vestibulares do protocone e metacônulo são mais íngremes que as faces linguais, com esta última representando aproximadamente um terço da largura do dente na vista oclusal. Um curto cíngulo lingual fecha o vale transversal na margem lingual entre o protocone e o metacônulo. No NTM P12013, o cíngulo lingual é cúspide.
A metade anterolingual de um molar superior (NTM P12014) tem morfologia geralmente semelhante à NTM P12012 e NTM P12013. Considera-se que este fragmento molar representa um M1 com base no seguinte: a porção posterior parece ter sido mais larga que a porção anterior; o paraconule é relativamente mais proeminente; a depressão na face anterior, derivada do ponto de contato com (presumivelmente) a face posterior de P3, é pequena e circular (em vez de extensa bucolingualmente). O paraconule é cúspide, posicionado na face anterolingual do paracone. Em comum com o espécime NTM P12014, duas cristas proeminentes, alinhadas paralelamente uma à outra, descem pela face anterolingual do metacone (Fig. 3 ). A colocação dessas duas cristas em relação ao metacone reflete a do paraconule em relação ao paracone.
Madakoala sp. cf. M. devisi. Espécimes molares inferiores do maior fascolarctídeo no Pwerte Marnte Marnte LF comparam-se estreitamente aos de Madakoala devisi em tamanho ( Tabela S2, Suplemento 2 ) e morfologia (Fig. 4 a – c; e Suplemento 3 para descrições). As espécies de Madakoala são conhecidas nas formações Namba e Etadunna do Oligoceno tardio, no sul da Bacia do Lago Eyre 50 , 51 . Notamos que os espécimes de molares inferiores (NTM P12015 e NTM P12016) são mais semelhantes em morfologia a M. devisi do que os espécimes de molares superiores (Fig. 4 d – g; NTM P14005; NTM P12017), sendo estes últimos também semelhantes às espécies de Perikoala (ver Suplemento 3 ). É possível que esses espécimes não derivem do mesmo táxon. A ocorrência de representantes de ambos os gêneros em uma única assembleia foi relatada em Ditjimanka LF da Formação Etadunna 50 , 51 . Nesta fase, consideramos mais parcimonioso tratar estes espécimes como pertencentes a uma única espécie aliada a M. devisi .
Espécimes de Madakoala sp. cf. M. devisi e Nimiokoala sp. indet. do site Pwerte Marnte Marnte, com desenhos anotados. Madakoala sp. cf. M. devisi : ( a , b ) fileira parcial do molar inferior direito preservando m1–m4 (NTM P12015); ( c ) talonídeo m3 esquerdo (NTM P12016); ( d , e ) paracone da direita M 2? (NTM P12017); ( f , g ) esquerda M1 (NTM P14005). Nimiokoala sp. indet.: ( h , i ) protocone esquerdo (NTM P12018); ( j , k ) protoconídeo direito (NTM P12019). As setas de orientação (Anterior, Bucal) referem-se aos espécimes do molar inferior direito ( a , b , j , k ) e do molar superior esquerdo ( f , g , h , i ). final entoconídeo hyd , hipoconídeo lc , cíngulo lingual , metacônulo mcl , me metacone, med metaconídeo ncd , cúspide neomórfica nmcl , neometaconule , pa paracone, pacl paraconule, pc precingulum, popac postparacrista, popr postprotocrista, poprd postprotocristid, pr protocone, prd protoconid, prmec premetacrista, prprd preprotocristid, rpopac remanescente pósparacrista, rprmec remanescente postmetacrista, rstA remanescente cúspide estilar A, stA cúspide estilar A, s tB denomina cúspide B, stD denomina cúspide D, stE denomina cúspide E.
Nimiokoala sp. indet. Two molar fragments (NTM P12017, NTM P12018: Fig. 4h–k) compare best with species of Nimiokoala, though both are too fragmentary for systematic assessment beyond generic referral (Supplementary 4). The only other late Oligocene record of the genus, represented by a partial dentary (SAMA P19952) from the Namba Formation52, is not yet referred to a species. The type species, Nimiokoala greystanesi, is known from the Early to Middle Miocene of Riversleigh, north-west Queensland52,53,54,55,56.
Results of phylogenetic analyses. Parsimony analyses with all taxa included, and ordering of states where morphoclines were inferred, generated a strict consensus of 58 most parsimonious trees, each of 498 steps (Fig. S4, Supplementary 7). The strict consensus tree had a consistency index of 0.39 and a retention index of 0.77. Our maximum parsimony (strict consensus) and undated Bayesian analyses (majority rule consensus topology of the post-burnin trees) recovered similar topologies to the equivalent analyses of Crichton et al.39 (Figs. S2–S5, Supplementary 7). This reflects that relatively few modifications were made to the morphological characters of Crichton et al.39. In our unconstrained Bayesian analysis, Lumakoala blackae was weakly supported (BPP = 0.61) as the sister taxon to a moderately supported Phascolarctidae + Vombatomorphia (BPP = 0.82); whereas, under maximum parsimony, it formed a polytomy (BS = 0.88) with Cercartetus lepidus, Thylacoleonidae, and a weakly supported Phascolarctidae + Vombatomorphia (BS = 14%). The placement of L. blackae outside of Phascolarctidae + Vombatomorphia was supported by two character-transformations: stC not linked to paracone on M2 (C.I. = 0.33: char. 41); and metacone not linked to stD on M2 (C.I. = 0.50: char. 47). Thylacotinga bartholomaii consistently formed the sister taxon to Diprotodontia (BBP = 0.97; BS = 0.87).
A hipótese alternativa para a colocação de Lumakoala blackae , testada restringindo-a a um clado com fascolarctídeos, rendeu 52 árvores mais parcimoniosas, cada uma com comprimento de 499 passos, diferindo daquela da topologia irrestrita em apenas um passo. Isto foi associado a uma diferença não significativa (P = > 0,05) na distribuição do estado de caráter, conforme indicado pelo teste unilateral de Templeton (P = 0,28–0,43). Como tal, a hipótese de que Lumakoala blackae é um fascolarctídeo não pode ser rejeitada. O consenso estrito das árvores mais parcimoniosas da análise restrita recuperou Phascolarctidae como uma politomia composta por Lumakoala blackae , Priscakoala lucyturnbullae e um clado compreendendo os demais táxons de coala ( Fig. S5, Suplementar 7 ).
Discussão
The discovery of phascolarctids in the Pwerte Marnte Marnte fossil site provides the first record of koalas from the Northern Territory in both the historical and fossil records. Assuming Lumakoala blackae is indeed a phascolarctid, the site preserves three koala species, representing the equal highest number recorded from a single deposit, along with Camel Sputum Site from the early Miocene (Faunal Zone B) of Riversleigh57,58. Surprisingly, the fossil sites from the late Oligocene (Faunal Zone A) of Riversleigh have not yielded any phascolarctids55,56. From the late Oligocene Namba and Etadunna Formations of central Australia, phascolarctids are represented in most local faunas, with a single taxon in most, and two in some50. It has been suggested that the high early Miocene koala diversity from Riversleigh may correlate with greater plant diversity in these presumably rainforest communities54. A elevada diversidade de fascolarcídeos no Pwerte Marnte Marnte LF pode, portanto, também indicar a presença de uma elevada diversidade de plantas num paleoambiente arborizado.
Se Lumakoala blackae gen. e sp. novembro. é um fascolarctídeo, é o décimo gênero e está entre os geologicamente mais antigos conhecidos. Sua descoberta enfatiza ainda mais o bem conhecido 54 , 59 rotatividade de fascolarctídeos do final do Oligoceno ( Lumakoala, Madakoala , Perikoala , Nimiokoala e Litokoala ) ao início do Mioceno ( Nimiokoala e Litokoala , Priscakoala ). Lumakoala blackae era relativamente pequena, produzindo estimativas de massa corporal de 2,2–2,6 kg ( Tabela S3, Suplementar 8 ) com base em equações de regressão 60 de comprimento M2 e M3. Isso se enquadra na faixa de tamanho do gambá moderno Trichosurus vulpecula (1,5–4,5 kg 61 ), e é marcadamente menor do que outros fascolarctídeos alegadamente plesiomórficos (por exemplo, espécies de Priscakoala , Madakoala , Perikoala , Koobor e Invictokoala ), todos os quais se enquadram na faixa de tamanho das espécies modernas de coala (ver 54 : 4,1–13,5kg 62 ). De fato, L. blackae está entre os menores fascolarctídeos conhecidos, juntamente com espécies de Litokoala (medidas na Tabela S1, Suplementar 2 ).
Unlike the selenodont molars of unequivocal phascolarctids, those of L. blackae have a wide stylar shelf with large, conical stylar cusps B and D, that are unconnected from the postparacrista and premetacrista, respectively (Fig. 5). From a functional standpoint, this crown morphology, and the associated wear pattern, suggest no occlusion between the stylar cusps and the hypoconid during the transverse (shearing) phase of the power stroke. Instead, stylar cusps B and D would have overhung the buccal face of the lower molars, presumably abetting puncture-crushing during the initial vertical phase. Direct occlusion between the stylar cusps and the hypoconid may, nonetheless, have been possible as an artifact of high tooth wear later in life history. More generally, the low crown height and poorly developed enamel crenulations are consistent with adaptation for relatively soft foods. We hypothesize that L. blackae had a primarily plant-based diet, although it may have opportunistically consumed insects like many extant possums62.
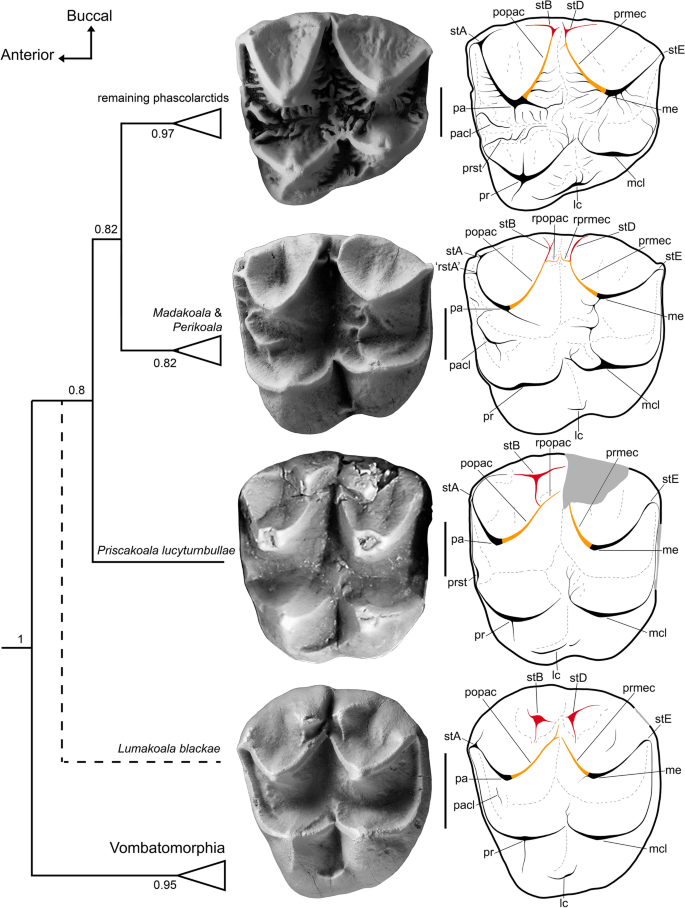
Concatenated phascolarctid interrelationships, contextualized by upper molars from representative taxa in occlusal view, with annotated line drawings. From top to bottom: Phascolarctos cinereus (M2, FUR 095); Madakoala devisi (M2, FUR unregistered; field number BC 20–112); Priscakoala lucyturnbullae (M2, QM F20203) adapted from Black et al.58; and the probable phascolarctid Lumakoala blackae, gen. et sp. nov. (Left M2 or 3, NTM P12012). Topology based on undated Bayesian analysis, presented as a majority rule consensus, with numbers at nodes representing Bayesian posterior probabilities (Fig. S3, Supplementary 7 for complete tree). Lumakoala blackae was excluded from this analysis; its hypothesized placement within Phascolarctidae is indicated by a dashed line. lc lingual cingulum, me metacone, mcl metaconule, pa paracone, pacl paraconule, popac postparacrista, pr protocone, prmec postmetacrista, prst protostyle, rpopac remnant postparacrista, rprmec remnant postmetacrista, rstA remnant stylar cusp A, stA stylar cusp A, stB stylar cusp B, stD stylar cusp D, stE stylar cusp E. Scale bar equals 2 mm.
A presença desta morfologia molar bunoselenodonte em um marsupial diprotodontiano é filogeneticamente significativa. Isso ocorre porque a independência estrutural das cúspides estilares B e D, e um centrocrista contínuo (postparacrista + premetacrista), são simplesiomorfias inequívocas das outras ordens australidelfianas (Microbiotheria, Peramelemorphia, Dasyuromorphia e Notoryctemorphia), bem como em Didelphimorphia e muitas outras ordens metatherianas. clados (por exemplo, veja 5 ).
Nossas análises filogenéticas bayesianas irrestritas implicam L. que blackae é um vombatiforme de caule, irmão de Phascolarctidae + Vombatomorphia, embora com suporte fraco (BBP = 0,61; para árvores completas, ver Fig. S2, Suplemento 7). A inclusão de L. blackae nas análises também resultou em menor suporte para a monofilia de Phascolarctidae e Vombatiformes ( Figs. S2 e S3, Suplementar 7 ), refletindo que a morfologia conhecida do novo táxon confunde os limites desses clados. Esta má resolução pode não ser surpreendente, dado que L. blackae só pôde ser pontuada para 18% dos caracteres morfológicos, uma vez que é conhecida apenas pelos molares superiores. Além disso, não há sinapomorfias óbvias nos molares superiores de Phascolarctidae, com a possível exceção de um paracônulo em M1, que está presente em todos os fascolarctídeos, exceto Priscakoala lucyturnbullae 58 . Com base em um teste de Templeton, o comprimento da árvore da topologia em que L. blackae é restrito para formar um clado com fascolarctídeos definitivos não é significativamente pior do que a topologia irrestrita em que L. blackae é um vombatiforme de caule ( Figs. S4 e S5, Suplementar 7 ). Nesta fase, consideramos mais apropriado considerar L. blackae um provável fascolarctídeo.
Dentro de Phascolarctidae, a morfologia dos molares superiores de L. blackae é mais semelhante à de P. lucyturnbullae . Ambos são de coroa baixa, possuem cúspides estilares proeminentes, ornamentação simples da superfície do esmalte e não possuem neometacônulo (Fig. 5 ). Priscakoala lucyturnbullae já foi visualizada 58como o fascolarctídeo mais plesiomórfico conhecido até o momento porque: não possui paraconule e neometaconule em M1; tem um protostilídeo fracamente expresso em m1; e possui molares selenodontes simples, não crenulados. Nesta base, os molares de L. blackae poderiam ser considerados ligeiramente mais derivados do que os de P. lucyturnbullae porque um paraconule está presente em M1 e também (embora apenas fracamente desenvolvido) em M2. No entanto, argumentamos que a morfologia molar superior de L. blackae é, na verdade, mais plesiomórfica do que a de P. lucyturnbullae , porque estende o morfoespaço molar dos fascolarctídeos de variações em uma forma estrita de selenodonte, até a bunoselenodontia. Da extremidade mais plesiomórfica para a mais derivada desta proposta de morfoclina de fascolarctídeos, observamos: uma redução da plataforma estilar; integração das cúspides estilares B e D na pós-paracrista e pré-metacrista, respectivamente; redução da cúspide estilar B em relação às cúspides estilares A e D; e aumento da obliquidade do ângulo formado na junção pós-protocrista-premetaconulecrista (Fig. 5 ).
A morfologia molar de Lumakoala blackae também é intermediária entre aquela dos selenodontes vombatomorfos basais e a condição metatheriana arquetípica (Fig. 6 ). Em particular, confirma que alguns selenodontes vombatomorfos retêm restos de uma centrocrista contínua, lingual às grandes cúspides estilares cúspides B e D em M1; estes incluem os ilariídeos Kuterintja ngama (ver fig. 2 63 ) e Ilaria illumidens (Fig. 6 ), bem como o ?wynyardiid Muramura pinpensis . A atrofia dessas estruturas ocorre de forma gradual, de posterior para anterior ao longo da fileira molar. O pós-paracrista e o pré-metacrista são perdidos em vombatiformes mais derivados (Fig. 6 ).
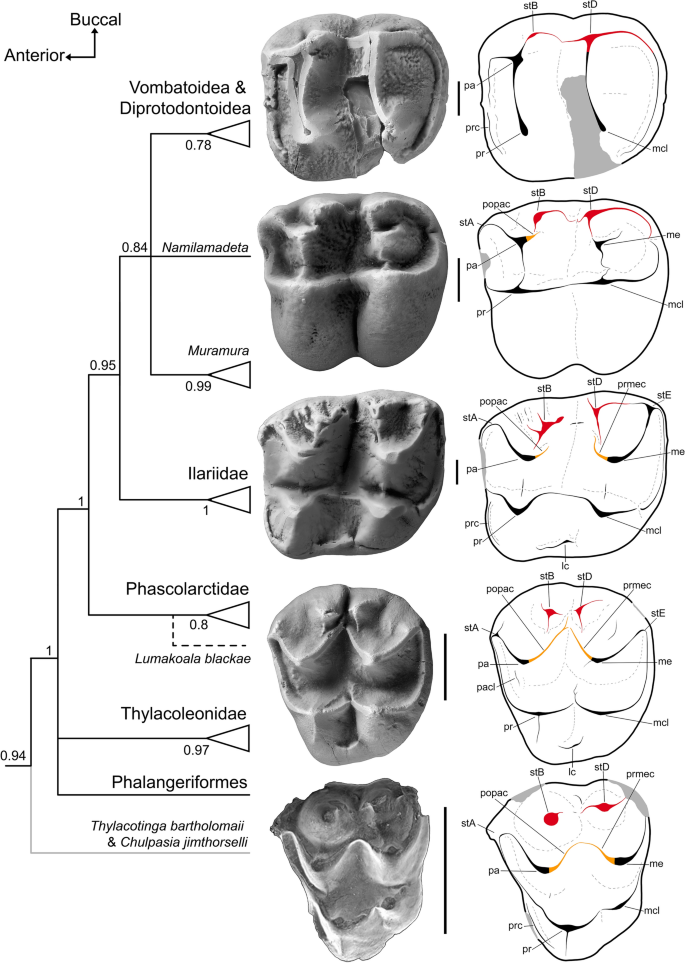
Inter-relações vombatiformes, contextualizadas por molares superiores de táxons representativos em vista oclusal, com desenhos de linha anotados. De cima para baixo: o diprotodontóide, Raemeotherium yatkolai (RM1, SAMA P43060: fotografia espelhada); ?wynyardiid, Namilamadeta crassirostrum (LM1, elenco de AR 9745); ilariídeo, Ilaria illumidens (M1, SAMA P43055); e o provável fascolarctídeo, Lumakoala blackae , gen. e sp. nov., (isolado esquerdo M2 ou 3, NTM P12012). O suposto polidolopimórfico Chulpasia jimthorselli é representado na parte inferior por um molar superior (M1 ou 2 esquerdo, QM F50411). Topologia concatenada baseada na análise bayesiana sem data, apresentada como um consenso de regra majoritária, com números nos nós representando probabilidades posteriores bayesianas ( Fig. S3, Suplemento 7 para árvore completa). Lumakoala blackae foi excluída desta análise; sua colocação hipotética dentro de Phascolarctidae é indicada por uma linha tracejada. lc cíngulo lingual, mcl metaconule, me metacone, pa paracone, pacl paraconule, popac postparacrista, pr protocone, prc precingulum, prmec premetacrista, prst protostyle, stA cúspide estilar A, stB cúspide estilar B, stD cúspide estilar D, stE cúspide estilar E. A barra de escala é igual a 2 mm.
É menos claro como as homologias das cúspides do estilo metatheriano se traduzem nos molares um tanto bunolofodontes dos tilacoleonídeos e na maioria dos primeiros falangeriformes (Fig. 7 ; ver 5,36,38,64 and references therein). Thylacoleonids have been interpreted, by some, as retaining the symplesiomorphic condition of a continuous centrocrista and a distinct stylar shelf38,65,66. It has also been suggested that that the small buccal-most structures on the M1 of thylacoleonids are actually neomorphic, and that the primary buccal cusps, traditionally referred to as the paracone and metacone, are actually stylar in origin5. Many early macropodoids and some phalangeroids bear a small cuspate ridge on the posterobuccal face of the paracone on M138, which has traditionally been identified as stylar cusp C36,67. No entanto, muitos falangeriformes iniciais também apresentam cúspides centrais em seus molares superiores, mais comumente na porção posterior de M1. A última configuração levou vários autores a sinalizar a possibilidade de que as cúspides tradicionalmente chamadas de paracone e metacone possam, em vez disso, representar as cúspides estilares B e D, respectivamente 68 , 69 , e que quaisquer estruturas subsidiárias vestibulares a eles são neomórficas 5 .
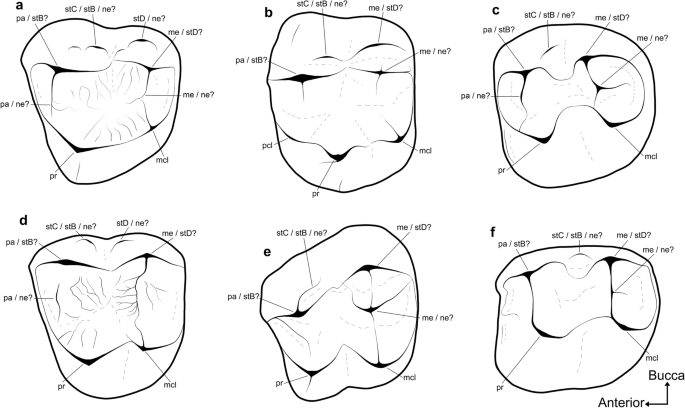
Desenhos anotados de molares superiores de diprotodontianos não vombatiformes representativos, ilustrando a ambigüidade em relação à homologia de suas estruturas bucais. O tilacoleonídeo ( a ) Lekaneleo roskellyae (RM1, espelhado; QM F23442); falangeróides ( b ) Burramys wakefieldi (LM1; SAMA P40931) e ( e ) Durudawiri inusitatus (RM1, espelhado; QM F31468); macropodoides ( c ) Priscus Palaeopotorous (RM1 ou M2, espelhado; NMV P157547) e ( f ) Nambaroo tarrinyeri (LM1 ou M2, 157528); e miminipossumida? ( d ) gen. e sp. indet. (LM1 ou M2, Fauna Local de Ditjimanka; SAMA P19856). mcl metaconule, me metacone, ne estrutura neomórfica, pa paracone, pcl protoconule, pr protocone, stB cúspide estilor B, stC cúspide estilor C, stD cúspide estilor D. Fora de escala.
À luz da morfoclina gradual observada entre as topografias molares dos gêneros de fascolarctídeos (Fig. 5 ), e mais geralmente entre as famílias vombatiformes (Fig. 6 ), postulamos que a morfologia molar do bunoselenodonte de L. blackae está próxima da condição ancestral para Vombatiformes; isso é congruente com a posição vombatiforme do caule para este táxon na análise bayesiana irrestrita. É digno de nota que as estimativas de massa corporal para Lumakoala blackae (2,2–2,6 kg: Tabela S3, Suplementar 8 ) também estão dentro dos limites previstos para o ancestral dos Vombatiformes (1–5,5 kg) 30 . Com exceção da proeminência relativa do metacônulo, esta morfologia provavelmente não difere marcadamente da condição ancestral da Diprotodontia. Essas observações implicam que o clado da coroa Diprotodontia (Vombatiformes + Phalangeriformes + Thylacoleonidae) manteve a condição australidelfiana simplesiomórfica de cúspides estilares B e D estruturalmente independentes e uma centrocrista contínua. Se correto, a redução e/ou integração das cúspides estilares B e D nas estruturas proximais ocorreu de forma convergente em várias linhagens diprotodontianas à medida que evoluíram dentições distintas e especializadas.
A maioria das estimativas de divergência molecular implica que os diprotodontianos estavam presentes quando os sedimentos do início do Eoceno perto de Murgon foram depositados (por exemplo, 6 , 7 , 8 , 9 , 10 , 11 , 12 ), mas nenhum foi identificado no Tingamarra LF associado 21 . A forma molar plesiomórfica de L. blackae fornece uma ligação inesperada entre as morfologias molares dos táxons Tingamarran Chulpasia jimthorselli e Thylacotinga bartholomaii e os primeiros diprotodontianos indiscutíveis (Fig. 6 ). Correspondentemente, esses táxons de Tingamarran são diprotodontianos antecedentes plausíveis. Os principais atributos molares possuídos por esses táxons de Tingamarran incluem: cúspides estilares B e D proeminentes e estruturalmente independentes; um centrocrista contínuo; um metacônulo pequeno, mas distinto; um canto póstero-vestibular que forma um ângulo > 60°; e um paracone e protocone que são de tamanho subigual em M2 (ver Suplemento 9 para comparações adicionais).
Thylacotinga bartholomaii and Chulpasia jimthorselli have been reconstructed as omnivores23,25, with body mass estimates of 3.2 kg and 0.2 kg, respectively27; the former being considerably larger than other mammals described to date from the Tingamarra LF. It has been suggested that these chulpasiines became extinct during the Palaeogene23. When included in our primarily vombatiform phylogenetic analyses, Thylacotinga bartholomaii was consistently recovered with strong support as the sister taxon to Diprotodontia (Fig. 6), to the exclusion of the representative agreodontians (for complete trees, Figs. S2–S5 Supplementary 7). A link between these Tingamarran taxa and definitive diprotodontians is attractive in its simplicity, supporting continuity between the early Eocene and late Oligocene Australian marsupial faunas, but we recognize that this may be an artifact of the very poor early Paleogene fossil record from Australia.
Thylacotinga bartholomaii e Chulpasia jimthorselli , juntamente com seu congênere C. mattaueri do final do Paleoceno ou início do Eoceno do Peru, foram colocados na subfamília Chulpasiinae e referidos à ordem principalmente sul-americana Polydolopimorphia 23 . No entanto, permanece a controvérsia sobre a relação de C. jimthorselli com C. mattaueri e, portanto, as inferências biogeográficas que podem ser tiradas 70 , 71 . Isto ocorre porque todos os três táxons são conhecidos de molares isolados 23 , 25 , 72 , e foram agrupados com base em características bunodontas generalizadas que são conhecidas por serem altamente homoplásticas dentro de Metatheria 2 , 73 , 74 . Ainda não foi publicada uma análise filogenética formal que teste suas relações entre si e com outros marsupialiformes bunodontes e não bunodontes. Como tal, uma afinidade diprotodontiana para Thylacotinga bartholomaii e Chulpasia jimthorselli pode não necessitar de uma distribuição pan-Gondwanana para a ordem australidelfiana (como estaria implícito se estes dois táxons estivessem intimamente relacionados aos táxons sul-americanos). Contudo, vale reconhecer que Lorente et al. 15 descreveu tarsos marsupiais australianos isolados, da localidade de La Barda (que agora parece ter ~ 43,5 milhões de anos ou menos 75 ) no sul da Argentina, que se enquadraram na coroa Diprotodontia em sua análise filogenética. Supostas chulpasiínas não foram descritas em La Barda ou em um segundo local com uma fauna de mamíferos taxonomicamente semelhante, Laguna Fria 76 , e não está claro se os tarsos de La Barda pertencem a algum dos táxons do local conhecidos a partir de restos dentários 2 , 15 .
Também foi levantada a hipótese, embora controversa, de que a Diprotodontia pode ser aliada à Polydolopimorphia (a ordem à qual Sige et al. 23 , referido Chulpasiinae) juntos derivando de um ancestral semelhante ao microbioteriano 15,77,78,79,80,81,82,83. However, the phylogenetic affinities of polydolopimorphians are quite controversial, both with respect to their ordinal relationships and their monophyly as a single clade2,74. Furthermore, phylogenomic and retroposon insertion data provides compelling evidence that Microbiotheria is sister to the modern Australian marsupial radiation as whole6,14,84 em vez de apenas Diprotodontia especificamente.
Independentemente de Thylacotinga e Chulpasia serem ou não de fato diprotodontianos iniciais, levantamos a hipótese de que sua morfologia molar bunodonta - bem adequada para uma dieta onívora - é provavelmente muito próxima da condição diprotodontiana ancestral. Ao contrário dos metatherianos tribosfênicos, o golpe de força na maioria dos diprotodontianos existentes é dividido em uma fase vertical e uma fase transversal, com a última realizada puxando os molares inferiores do lado oclusal medialmente através dos molares superiores. 85. Interestingly, the upper molar specimens of T. bartholomaii show distinct wear facets on the crests linking stB and stD, presumably due to occlusion with the hypoconid, indicating a strong transverse component to the masticatory stroke. Indeed, much of the required change in molar morphology from the form of Thylacotinga and Chulpasia, to that of Lumakoala and other selenodont diprotodontians, can be explained as adaptations for facilitating a stronger transverse component for specialized herbivory. The formation of continuous transverse crests between the paracone and metacone, and stylar cusps B and D, respectively, increases shearing blade length; whereas loss of the link between the postparacrista and premetacrista allows the hypoconid to move further buccally, forming a larger crushing basin. These changes, alongside cranial adaptions enabling a larger transverse masticatory stroke, probably represented crucial steps in the transition from insectivorous to herbivorous diets in diprotodontian evolution.
Métodos
Os espécimes de fascolarctídeos relatados neste estudo foram recuperados de ~ 2 toneladas de calcário extraído dos leitos fósseis de Pwerte Marnte Marnte em expedições em 2014 e 2020, lideradas por A. Couzens e A. Crichton, respectivamente. A rocha fossilífera foi processada durante 2020–2022 no Laboratório de Paleontologia da Universidade Flinders usando uma combinação de ataque com ácido acético (5–10%) e métodos mecânicos, por exemplo, serras de rocha e ferramentas pneumáticas de micro-macaco 38 . Para amostras comparativas utilizadas, consulte o Suplemento 2 .
Terminology. Higher-level systematic nomenclature follows Aplin and Archer86, with the exception of: the use of the suborder Phalangeriformes Szalay87, sensu Woodburne88; the superfamily Vombatoidea for the clade that includes Vombatidae + Mukupirnidae + Maradidae following Beck et al.30 and Crichton et al.39; and the subordinal placement of Thylacoleonidae as Diprotodontia incertae sedis following Beck et al.5. We also use ?Wynyardiidae to refer to species within the genera Namilamadeta and Muramura, because their inclusion within the family Wynyardiidae has never been robustly demonstrated33,89,90,91. Biostratigraphic nomenclature follows Woodburne et al.28, Archer et al.92 and Travouillon et al.93. The ages of vertebrate bearing localities from the Namba and Etadunna Formations follows Woodburne et al.28 and Megirian et al.29, and those of Riversleigh World Heritage Area follow Archer et al.92 and Travouillon et al.93 sensu Woodhead et al.53.
A homologia da posição molar segue Luckett 94. Molar cusp nomenclature follows Rich et al.95 with the exception of: the structure reported therein as the hypocone, which is here referred to as the metaconule following Tedford and Woodburne96; and the buccal-most cusps on the upper molars in diprotodontians, which are discussed in Supplementary 1.
Abreviaturas da instituição . Coleção AR Archer, Universidade de Nova Gales do Sul, Sydney, Nova Gales do Sul; Departamento de Paleontologia de Vertebrados do AMNH , Museu Americano de História Natural, Nova York, EUA; Departamento de Paleontologia FUR , Faculdade de Ciências e Engenharia, Universidade Flinders, Bedford Park, Adelaide, Sul da Austrália; Paleontologia NMV P , Museu Victoria, Melbourne; Museu NTM P da Austrália Central, Museu e Galeria de Arte do Território do Norte, Alice Springs; QM F , Coleção de Fósseis do Museu de Queensland, Brisbane; S SAMA P Paleontologia, Museu da Austrália do Sul, Adelaide, Sul da Austrália.
Análises filogenéticas. Para avaliar as relações filogenéticas de Lumakoala blackae , adotamos o conjunto de dados morfológicos de Crichton et al. 39 , baseado no de Beck et al. 30 , compreendendo 124 caracteres craniodentais e 20 pós-cranianos, que foram pontuados para 46 táxons. Incluímos mais 13 táxons, totalizando 59, a saber: Lumakoala blackae gen. e sp. nov., Koobor notabilis, Stelakoala riversleighensis e Invictokoala monticola (Phascolarctidae); Thylacotinga bartholomaii (suposto polidolopimórfico); Djarthia murgonensis (Australidelfia); Naraboryctes philcreaseri (Notoryctemorphia); ' Bulungu spp.' (Peramelemorfia); Nimbacinus dicksoni , Muribacinus gadiyuli e Badjcinus turnbulli (Thylacinidae); bem como Keeuna woodburnei e Ankotarinja tirarensis (supostos dasyuromorfos). Para acomodar agreodontianos adicionais do grupo externo e alterações na homologia da cúspide estilar B em Diprotodontia, minimizando a pseudoreplicação: três caracteres foram excluídos (char. 27, Lophodonty; 37, Preparacrista; 49, Colocação relativa de stC em M1); 19 foram modificados; e 17 foram adicionados. A matriz de caracteres resultante compreendeu 138 caracteres craniodentais e 20 caracteres pós-cranianos (ver Suplementar 5 e 6 ).
Undated Bayesian analysis of the morphological dataset was carried out in MrBayes 3.2.7a 97, using the Markov Chain Monte Carlo (MCMC) approach, with gamma rate variability implemented for morphological data maintaining the assumption that only variable characters were scored. The Bayesian analyses were run for 15 million generations, using four independent runs of four chains (one cold and three heated chains, with the temperature of the heated chains set to the default value of 0.2), sampling trees every 1000 generations and a burn-in fraction of 25%. The post-burn-in trees were summarised using a majority rule consensus of all compatible groups, with Bayesian posterior probabilities as support values.
Análises de máxima parcimônia também foram realizadas no conjunto de dados morfológicos, no TNT versão 1.5 98. The tree search involved an initial “new technology” search with sectorial search, ratchet, drift, and tree fusing that was run until the same minimum tree length was found 1000 times. From these saved trees a “traditional” search was applied using the tree bisection resection (TBR) swapping algorithm, with the resulting most parsimonious trees combined into a strict consensus tree. Support values for branch nodes were calculated using 2000 standard bootstrap replicates, implemented using a “traditional” search, which results in output as absolute frequencies.
Análises adicionais de parcimônia foram implementadas no PAUP 4.0a169 99 , para testar se havia diferenças significativas no comprimento da árvore e na distribuição do estado de caráter de uma hipótese topológica alternativa, que incluía uma restrição positiva que impunha Lumakoala blackae dentro de Phascolarctidae. Para comparar estatisticamente essas hipóteses filogenéticas, uma única árvore mais parcimoniosa foi escolhida aleatoriamente a partir das análises irrestritas e comparada com 50 árvores aleatórias mais parcimoniosas das análises restritas usando um teste de Templeton unilateral. 100 . Isso difere de um teste de Templeton pareado porque o valor P é reduzido pela metade 101 .
Disponibilidade de dados
Os dados gerados e analisados durante este estudo estão incluídos neste artigo publicado e em seu arquivo de informações suplementares (disponível em https://doi.orgTBA ).
Referências
Beck, R. M. D. In Handbook of Australasian Biogeography, Ch. 13 (ed. Ebach, M. C.) 329–366 (CRC Press, 2017).
Beck, R. M. D. In American and Australasian marsupials: An Evolutionary, Biogeographical, and Ecological Approach, Ch. 1 (eds. Cáceres, N. C. & Dickman, C. R.) 23–87 (Springer, 2023).
White, L. T., Gibson, G. M. & Lister, G. S. A reassessment of paleogeographic reconstructions of eastern Gondwana: Bringing geology back into the equation. Gondwana Res. 24, 984–998 (2013).
Stotz, IL, Tassara, A. & Iaffaldano, G. O fluxo de Poiseuille impulsionado pela pressão, herdado da circulação do manto mesozóico, levou à separação do Eoceno entre a Austrália e a Antártica. J. Geofísica. Res. Sólido 126 , 9945. https://doi.org/10.1029/2020JB019945 (2021).
Beck, RMD, Voss, RS & Jansa, SA Morfologia craniodental e filogenia de marsupiais. Touro. Sou. Mus. Nat. História. 457 , 1–352 (2022).
Duchêne, DA et al. A análise do espaço da árvore filogenômica resolve as relações entre as famílias de marsupiais. Sist. Biol. 67 , 400–412 (2018).
Mitchell, KJ et al. Filogenia molecular, biogeografia e evolução de preferência de habitat de marsupiais. Mol. Biol. Evol. 31 , 2322–2330 (2014).
Upham, N. S., Esselstyn, J. A. & Jetz, W. Inferring the mammal tree: Species-level sets of phylogenies for questions in ecology, evolution, and conservation. PLoS Biol. 17, e3000494 (2019).
Beck, R. M. D. A dated phylogeny of marsupials using a molecular supermatrix and multiple fossil constraints. J. Mammal. 89, 175–189 (2008).
Meredith, RW, Krajewski, C., Westerman, M. & Springer, MS Relacionamentos e tempos de divergência entre as ordens e famílias de marsupialia. Mus. Norte. Arizona. 65 , 383–406 (2009).
Meredith, RW, Westerman, M. & Springer, MS Uma filogenia de Diprotodontia (Marsupialia) baseada em sequências de cinco genes nucleares. Mol. Filógeno. Evol. 51 , 554–571 (2009).
Meredith, RW et al. Impactos da Revolução Terrestre do Cretáceo e da extinção do KPg na diversificação dos mamíferos. Ciência 334 , 521–524 (2011).
Álvarez-Carretero, S. et al. Uma linha do tempo da evolução dos mamíferos em nível de espécie, integrando dados filogenômicos. Natureza 602 , 263–267 (2022).
Feng, S. et al. Classificação incompleta de linhagem e evolução fenotípica em marsupiais. Célula 185 , 1646–1660 (2022).
Lorente, M., Chornogubsky, L. & Goin, F. J. On the existence of non-microbiotherian Australidelphian marsupials (Diprotodontia) in the Eocene of Patagonia. Palaeontology 59, 533–547 (2016).
Hocknull, S. A. et al. Time is of the essence: three case studies from Queensland, Australia, demonstrating responses to fossil site loss through mining and development over differing time scales. J. Vertebr. Paleontol., Program and Abstracts 2019, 118.
Godthelp, H., Archer, M., Cifelli, R., Hand, SJ & Gilkeson, CF A mais antiga fauna de mamíferos terciários australianos conhecida. Natureza 356 , 514–516 (1992).
Long, JA, Archer, M., Flannery, TF & Hand, S. Mamíferos Pré-históricos da Austrália e Nova Guiné: Cem Milhões de Anos de Evolução (Univ. of New South Wales Press, Sydney, e Johns Hopkins Univ. Press, 2002 ).
Archer, M. et al. A história evolutiva e a diversidade dos mamíferos australianos. Austrália. Mamífero. 21 , 1–45 (1999).
Vickers-Rich, P. Em Paleontologia de Vertebrados da Australásia (eds. Vickers-Rich, P. et al .) 721–808 (Pioneer Design Studio e Monash University Publications Committee, 1991).
Black, KH, Archer, M., Hand, SJ & Godthelp, H. Na Terra e na Vida: Biodiversidade Global, Intervalos de Extinção e Perturbações Biogeográficas ao Longo do Tempo (ed. Talent, JA) 983–1078 (Springer, 2012).
Beck, R. M. D., Godthelp, H., Weisbecker, V., Archer, M. & Hand, S. J. Australia’s oldest marsupial fossils and their biogeographical implications. PLoS ONE 3, e1858 (2008).
Sigé, B. et al. Chulpasia and Thylacotinga, late Paleocene-earliest Eocene trans-Antarctic Gondwanan bunodont marsupials: New data from Australia. Geobios 42, 813–823 (2009).
Beck, RMD Um marsupial 'ameridelfiano' do início do Eoceno da Austrália apóia um modelo complexo de biogeografia marsupial do Hemisfério Sul. Naturwissenschaften 99 , 715–729 (2012).
Archer, M., Godthelp, H. & Hand, SJ Marsupial do início do Eoceno da Austrália. Kaupia 3 , 193–200 (1993).
Godthelp, H., Wroe, S. & Archer, M. Um novo marsupial do início do Eoceno Tingamarra Fauna local de Murgon, sudeste de Queensland: um marsupial australiano prototípico?. J. Mãe. Evol. 6 , 289–313 (1999).
Beck, RMD Um metatheriano faunívoro peculiar do início do Eoceno da Austrália. Acta Paleontol. Pol. 60 , 123–129 (2013).
Woodburne, M. O. et al. Land mammal biostratigraphy and magnetostratigraphy of the Etadunna Formation (Late Oligocene) of South Australia. J. Vert. Paleontol. 13, 483–515 (1994).
Megirian, D., Prideaux, G. J., Murray, P. F. & Smit, N. An Australian land mammal age biochronological scheme. Paleobiology 36, 658–671 (2010).
Beck, RMD et ai. Uma nova família de marsupiais diprotodontianos do último Oligoceno da Austrália e a evolução de wombats, coalas e seus parentes (Vombatiformes). Ciência. Rep. 10 , 1–13 (2020).
Black, K. Maradidae: Uma nova família de marsupiais vombatomorfos do final do Oligoceno de Riversleigh, noroeste de Queensland. Alcheringa 31 , 17–32 (2007).
Archer, M. Origens dos Phascolarctidas e o potencial do molar selenodonte na evolução dos marsupiais diprotodontes. Mem. Rainha. Mus. 17 , 367–371 (1976).
Tedford, RH et al. A descoberta de vertebrados do Mioceno, área do Lago Frome, Sul da Austrália. BMR J. Aust. Geol. Geofísica. 2 , 53–57 (1977).
Black, K. Descrição de novo material para Propalorchestes novaculacephalus (Marsupialia: Palorchestidae) do Mioceno médio de Riversleigh, noroeste de Queensland. Alcheringa 30 , 351–361 (2006).
Murray, P. Primitive marsupial tapirs (Propalorchestes novaculacephalus Murray and P. ponticulus sp. nov.) from the mid-Miocene of north Australia (Marsupialia: Palorchestidae). Beagle Records Museums Art Galleries Northern Territory 7, 39–51 (1990).
Passeio, WDL Jackmahoneya gen. novembro. e a gênese do molar macropodiforme. Mem. Assoc. Austral. Paleontol. 15 , 441–459 (1993).
Archer, M. Em Zoogeografia e Evolução de Vertebrados na Australásia (eds. Archer, M. & Clayton, G.) 585–631 (Hesperian Press, 1984).
Crichton, AI, Worthy, TH, Camens, A. & Prideaux, GJ Um novo gambá ektopodontídeo (Diprotodontia, Ektopodontidae) do Oligoceno da Austrália central e suas implicações para as inter-relações falangeróides. J. Vert. Paleontol. 2023 , e2171299. https://doi.org/10.1080/02724634.2023.2171299 (2023).
Crichton, AI et al. Uma nova espécie de Mukupirna (Diprotodontia, Mukupirnidae) do Oligoceno da Austrália central lança luz sobre as inter-relações vombatóides basais. Alcheringa https://doi.org/10.1080/03115518.2023.2181397 (2023).
Ride, WDL Sobre a evidência fóssil da evolução dos Macropodidae. Austrália. Zoológico. 13 , 6–16 (1971).
Lanyon, J. M. & Sanson, G. D. Koala (Phascolarctos cinereus) dentition and nutrition. I. Morphology and occlusion of cheekteeth. J. Zool. 209, 155–168 (1986).
Young, W. G., McGowan, M. & Daley, T. J. Tooth enamel structure in the koala, Phascolarctos cinereus: Some functional interpretations. Scann. Microsc. 1, 1925–1934 (1987).
Crompton, AW, Owerkowicz, T. & Skinner, J. Padrão motor mastigatório no coala ( Phascolarctos cinereus ): Uma comparação dos movimentos da mandíbula em herbívoros marsupiais e placentários. J. Exp. Zoológico. R: Ecol. Integr. Fisiol. 313 , 564–578 (2010).
Winge, H. As inter-relações dos gêneros mamíferos: Vol. 1 Monotremata, Marsupialia, Insectivora, Chiroptera, Edentata 1–418 (CA Reitzels Forlag, 1941).
Muirhead, J. & Filan, SL Yarala burchfieldi , um bandicoot plesiomórfico (Marsupialia, Peramelemorphia) de depósitos do Oligo-Mioceno de Riversleigh, noroeste de Queensland. J. Paleontol. 69 , 127–134 (1995).
Travouillon, KJ, Beck, RMD & Case, JA Peramelemorfos do Oligoceno Superior – Mioceno Médio Inferior das formações Etadunna, Namba e Wipajiri do Sul da Austrália. Alcheringa 45 , 109–125 (2021).
Travouillon, K. J., Gurovich, Y., Archer, M., Hand, S. J. & Muirhead, J. The genus Galadi: Three new bandicoots (Marsupialia, Peramelemorphia) from Riversleigh’s Miocene deposits, northwestern Queensland, Australia. J. Vert. Paleontol. 33, 153–168 (2013).
Murray, P. F. & Megirian, D. The Pwerte Marnte Marnte Local Fauna: A new vertebrate assemblage of presumed Oligocene age from the Northern Territory of Australia. Alcheringa 30, 211–228 (2006).
Black, KH, Louys, J. & Price, GJ Compreender a variação morfológica no coala existente como uma estrutura para identificação de limites de espécies em coalas extintos (Phascolarctidae; Marsupialia). J.Sist. Paleontol. 12 , 237–264 (2013).
Rico, TH et al. Em Paleontologia de Vertebrados da Australásia (eds. Vickers-Rich, P. et al .) 1005–1058 (Pioneer Design Studio em cooperação com o Monash University Publications Committee, 1991).
Woodburne, M., Tedford, R., Archer, M. & Pledge, N. Em Possums and Opossums: Studies in Evolution (ed. Archer, M.) 293–317 (Surrey Beatty & Sons Pty Ltd, 1987).
Black, K. & Archer, M. Nimiokoala gen. novembro. (Marsupialia, Phascolarctidae) de Riversleigh, noroeste de Queensland, com uma revisão de Litokoala. Mem. Rainha. Mus. 41 , 209–228 (1997).
Woodhead, J. et al. Desenvolvendo uma sequência cronológica datada radiometricamente para a mudança biótica neogênica na Austrália, na área do Patrimônio Mundial de Riversleigh, em Queensland. Gondwana Res. 29 , 153–167 (2016).
Black, K. H., Price, G. J., Archer, M. & Hand, S. J. Bearing up well? Understanding the past, present and future of Australia’s koalas. Gondwana Res. 25, 1186–1201 (2014).
Black, K. H. Middle Miocene origins for tough-browse dietary specialisations in the koala (Marsupialia, Phascolarctidae) evolutionary tree: Description of a new genus and species from the Riversleigh World Heritage Area. Mem. Mus. Vic. 74, 255–262 (2016).
Archer, M. et al. Situação atual da representação em nível de espécie em faunas de localidades fósseis selecionadas na Área do Patrimônio Mundial de Riversleigh, noroeste de Queensland. Alcheringa 30 , 1–17 (2006).
Travouillon, KJ, Escarguel, G., Legendre, S., Archer, M. & Hand, SJ O uso de MSR (Minimum Sample Richness) para comparações de assembleias de amostras. Paleobiologia 37 , 696–709 (2011).
Black, KH, Archer, M. & Hand, SJ Novo coala terciário (Marsupialia, Phascolarctidae) de Riversleigh, Austrália, com uma revisão da filogenética, paleoecologia e paleobiodiversidade dos fascolarctídeos. J. Vert. Paleontol. 32 , 125–138 (2012).
Price, GJ Em Paleontologia em Ecologia e Conservação , cap. 9 (ed. Louys, J.) 171–192 (Springer, 2012).
Myers, TJ Predição da massa corporal marsupial. Austrália. J. Zool. 49 , 99–118 (2001).
Moore, BD, Wallis, IR, Marsh, KJ & Foley, WJ Na Conservação da Fauna Florestal da Austrália (ed. Lunney, D.) 549–575 (Royal Zoological Society of New South Wales, 2004).
Van Dyck, S. & Strahan, R. Mammals of Australia 3rd edn. (New Holland Publishers, 2008).
Myers, T. & Archer, M. Kutjerintja ngama (Marsupialia, Ilariidae): Uma análise sistemática revisada com base em material do final do Oligoceno de Riversleigh, noroeste de Queensland. Mem. Rainha. Mus. 41 , 379–392 (1997).
Gillespie, AK Dois novos táxons de leões marsupiais (Marsupialia, Thylacoleonidae) do Mioceno Inferior e Médio da Austrália. Alcheringa , 1–16. https://doi.org/10.1080/03115518.2022.2152096 (2023).
Murray, P., Wells, R. & Plane, M. Em gambás e gambás. Estudos em Evolução (ed. Archer, M.) 433–466 (Surrey Beatty & Sons Pty Ltd, 1987).
Gillespie, A. Priscileo roskellyae sp. novembro. (Thylacoleonidae, Marsupialia) de Riversleigh, noroeste de Queensland. Mem. Rainha. Mus. 41 , 321–328 (1997).
Flannery, T. & Rich, THV Macropodóides da Formação Namba do Mioceno Médio, Sul da Austrália, e a homologia de algumas estruturas dentárias em cangurus. J. Paleontol. 60 , 418–447 (1986).
Woodburne, M. O., Pledge, N. & Archer, M. In Possums and Opossums: Studies in Evolution (ed. Archer, M.) 581–602 (Surrey Beatty & Sons Pty Ltd, 1987).
Crosby, K. & Archer, M. Durudawirines, a new group of phalangeroid marsupials from the Miocene of Riversleigh, northwestern Queensland. J. Paleontol. 74, 327–335 (2000).
Wilf, P., Cuneo, N. R., Escapa, I. H., Pol, D. & Woodburne, M. Splendid and seldom isolated: The paleobiogeography of Patagonia. Annu. Rev. Earth Planet. Sci. 41, 561–603 (2013).
Goin, FJ, Woodburne, MO, Zimicz, AN, Martin, GM & Chornogubsky, L. Em Uma Breve História dos Metatherianos Sul-Americanos Ch. 3, 77–124 (Springer, 2016).
Crochet, J.-Y. & Sigé, B. Os mamíferos de Chulpas (Formação Umayo, transição Cretáceo-Terciário, Peru): Dados preliminares. Documento. Laboratório. Geol. Lyon. 125 , 97–107 (1993).
Beck, RMD et ai. Uma nova família bizarra de Marsupialia (incertae sedis) do Plioceno Inferior do nordeste da Austrália: Implicações para a filogenia dos marsupiais bunodontes. J. Paleontol. 82 , 749–762 (2008).
Beck, RMD O crânio de Epidolops ameghinoi do Eoceno Inferior Fauna de Itaboraí, sudeste do Brasil, e as afinidades da extinta ordem marsupialiforme Polydolopimorphia. J. Mãe. Evol. 24 , 373–414 (2017).
Gosses, J. et al. Facies interpretation and geochronology of diverse Eocene floras and faunas, northwest Chubut Province, Patagonia, Argentina. Geol. Soc. Am. Bull. 133, 740–752. https://doi.org/10.1130/b35611.1 (2021).
Tejedor, M. F. et al. New early Eocene mammalian fauna from western Patagonia, Argentina. Am. Mus. Novit. 2009, 1–43 (2009).
Chornogubsky, L. & Goin, F. J. A review of the molar morphology and phylogenetic affinities of Sillustania quechuense (Metatheria, Polydolopimorphia, Sillustaniidae), from the early Paleogene of Laguna Umayo, southeastern Peru. J. Vert. Paleontol. 35, e983238 (2015).
Goin, F. J., Candela, A. M., Bond, M., Pascual, R. & Escribano, V. Una Nueva “Comadreja” (Mammalia, Marsupialia, ?Polydolopimorphia) del Paleoceno de Patagonia. Argentina. Publ. Electron. Assoc. Paleontol. Argent. 5, 71–78 (1998).
Goin, FJ, Candela, AM, Abello, MA & Oliveira, EV Os primeiros paucituberculatanos sul-americanos e seu significado na compreensão das radiações marsupiais 'pseudodiprotodontes'. Zoológico. J. Linn. Soc. 155 , 867–884 (2009).
Goin, FJ Em Predadores com bolsas: a biologia dos marsupiais carnívoros (eds. Jones, M. et al .) Ch. 3, 30–42 (CSIRO (Organização de Pesquisa Científica e Industrial da Commonwealth), 2003).
Goin, FJ & Candela, AM Novos marsupiais Paleógenos da Bacia Amazônica do leste do Peru. Nat. História. Mus. Los. Anjo. Cidade. Ciência. Ser. 40 , 15–60 (2004).
Oliveira, E. V. & Goin, F. J. In Os Marsupiais do Brasil: Biología, Ecología e Evolução (eds. Cáceres, N. C. & Monteiro Filho, E. L. A.) 299–320 (Springer, 2006).
Oliveira, É. V. & Goin, FJ Uma reavaliação dos metatherianos bunodontes do Paleógeno de Itaboraí (Brasil): Sistemática e idade da SALMA Itaboraiana. Rev. Brás. Paleontol. 14 , 105–136 (2011).
Gallus, S., Janke, A., Kumar, V. & Nilsson, M. A. Disentangling the relationship of the Australian marsupial orders using retrotransposon and evolutionary network analyses. Genome Biol. Evol. 7, 985–992 (2015).
Crompton, A. W. Masticatory motor programs in Australian herbivorous mammals: Diprotodontia. Integr. Comp. Biol. 51, 271–281 (2011).
Aplin, KP & Archer, M. Em Gambás e Gambás: Estudos em Evolução (ed. Archer, M.) XV – lxxii (Surrey Beatty & Sons Pty Ltd, 1987).
Szalay, FS Relações filogenéticas dos marsupiais. Geobios 15 , 177–190 (1982).
Woodburne, MO Em Mamíferos: Notas para um Curso Curto Vol. 8 (ed. Broadhead, TW) 48–71 (Knoxville, 1984).
Rich, THV & Archer, M. Namilamadeta snideri , um novo diprotodontano (Marsupialia, Vombatoidea) do Mioceno medial do Sul da Austrália. Alcheringa 3 , 197–208 (1979).
Pledge, NS Em Gambás e Gambás: Estudos em Evolução (ed. Archer, M.) 393–400 (Surrey Beatty & Sons Pty Ltd, 1987).
Megirian, D., Murray, P., Schwartz, L. & Von Der Borch, C. Poço Canguru do Oligoceno Superior Fauna Local do Calcário Ulta (novo nome) e clima da oscilação do Mioceno na Austrália central. Austrália. J. Ciência da Terra. 51 , 701–741 (2004).
Archer, M., Hand, SJ, Godthelp, H. & Creaser, P. Correlação dos sedimentos Cainozóicos da propriedade fóssil do Patrimônio Mundial de Riversleigh, Queensland, Austrália. Mem. Trav. EPHE Inst. Montpellier 21 , 131–152 (1997).
Travouillon, K. J., Archer, M., Hand, S. J. & Godthelp, H. Multivariate analyses of Cenozoic mammalian faunas from Riversleigh, northwestern Queensland. Alcheringa 30, 323–349 (2006).
Luckett, WP Em Filogenia de Mamíferos: Diferenciação Mesozóica, Multituberculados, Monotremados, Primeiros Therians e Marsupiais (eds Szalay FS, Novacek MJ e McKenna MC) 182–204 (Springer, 1993).
Rich, TH, Archer, M. & Tedford, RH Raemeotherium yatkolai , gen. e sp. nov., um diprotodontídeo primitivo do Mioceno medial do Sul da Austrália. Mem. Mus. Vic. 39 , 85–91 (1978).
Tedford, RH & Woodburne, MO Em Gambás e Gambás: Estudos em Evolução (ed. Archer, M.) 401–418 (Surrey Beatty & Sons Pty, 1987).
Ronquist, F. et al. MrBayes 3.2: Inferência filogenética bayesiana eficiente e escolha de modelo em um grande espaço de modelo. Sist. Biol. 61 , 539–542 (2012).
Goloboff, P. A., Farris, J. S. & Nixon, K. C. TNT, a free program for phylogenetic analysis. Cladistics 24, 774–786 (2008).
Swofford, D. L. PAUP*: Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0b10. (Sinauer Associates, 2002).
Templeton, AR Inferência filogenética a partir de mapas de locais de clivagem de endonucleases de restrição com referência particular à evolução de humanos e macacos. Evolução 37 , 221–244 (1983).
Goldman, N., Anderson, JP & Rodrigo, AG Testes de topologias baseados em probabilidade em filogenética. Sist. Biol. 49 , 652–670 (2000).
Travouillon, KJ, Butler, K., Archer, M. & Hand, SJ Duas novas espécies do gênero Gumardee (Marsupialia, Macropodiformes) revelam a evolução repetida da bilofodontia em cangurus. Alcheringa 46 , 105–128 (2022).
Reconhecimentos
Agradecemos a M.-A. Binnie e D. Stemmer, respectivamente, por fornecerem acesso ao Museu de Paleontologia e Coleções de Mamíferos do Sul da Austrália. Somos gratos a W. Klein da Estação Orange Creek por nos permitir acessar e extrair rochas fossilíferas do sítio fóssil Pwerte Marnte Marnte. Agradecemos a C. Burke, S. Arman, W. Handley e G. Gully pela organização e/ou assistência nas viagens de campo de 2014 e 2020 ao local. Reconhecemos o Povo Arrernte do Sul, conhecido como Corporação Aborígine Pwerte Marnte Marnte, pela custódia das terras onde a localidade fóssil está situada. Agradecemos também a G. Price e a um revisor anônimo, por seus comentários e sugestões atenciosos que melhoraram esta pesquisa.
Financiamento
A AIC foi apoiada pela bolsa de estudos do Programa de Treinamento em Pesquisa do Governo Australiano. O apoio para a viagem de campo de 2014 ao local foi fornecido por um Patterson Memorial Grant da Sociedade de Paleontologia de Vertebrados para AMCC.
Informação sobre o autor
Autores e Afiliações
Contribuições
A AIC contribuiu com reagentes/materiais/ferramentas de análise, extraiu calcário fossilífero, encontrou e preparou os espécimes fósseis, concebeu o artigo, analisou os dados, escreveu o artigo e preparou figuras e tabelas. THW revisou os rascunhos do artigo. ABC revisou os rascunhos do artigo. AMCC extraiu calcário fossilífero, revisou rascunhos do artigo e escreveu partes do artigo. O RMDB revisou os rascunhos do documento e escreveu partes do documento. GJP contribuiu com reagentes/materiais/ferramentas de análise, revisou rascunhos do artigo e escreveu partes do artigo. Todos os autores contribuíram para o desenvolvimento das hipóteses apresentadas no manuscrito.
autor correspondente
Declarações éticas
Interesses competitivos
Os autores declaram não haver interesses conflitantes.
Informações adicionais
Nota do editor
A Springer Nature permanece neutra em relação a reivindicações jurisdicionais em mapas publicados e afiliações institucionais.
Informação suplementar
Direitos e permissões
Acesso aberto Este artigo está licenciado sob uma Licença Creative Commons Atribuição 4.0 Internacional, que permite o uso, compartilhamento, adaptação, distribuição e reprodução em qualquer meio ou formato, desde que você dê o devido crédito ao(s) autor(es) original(is) e à fonte, forneça um link para a licença Creative Commons e indique se foram feitas alterações. As imagens ou outros materiais de terceiros neste artigo estão incluídos na licença Creative Commons do artigo, salvo indicação em contrário na linha de crédito do material. Se o material não estiver incluído na licença Creative Commons do artigo e o uso pretendido não for permitido por regulamentação legal ou exceder o uso permitido, você precisará obter permissão diretamente do detentor dos direitos autorais. Para visualizar uma cópia desta licença, visite http://creativecommons.org/licenses/by/4.0/ .
Sobre este artigo

Cite este artigo
Crichton, AI, Beck, RMD, Couzens, AMC et al. Um provável coala do Oligoceno da Austrália central fornece informações sobre a evolução inicial dos diprotodontianos. Sci Rep 13 , 14521 (2023). https://doi.org/10.1038/s41598-023-41471-0
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.