- 1
McColl, H. et al. O povoamento pré-histórico do Sudeste Asiático. Science 361 , 88–92 (2018).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 2
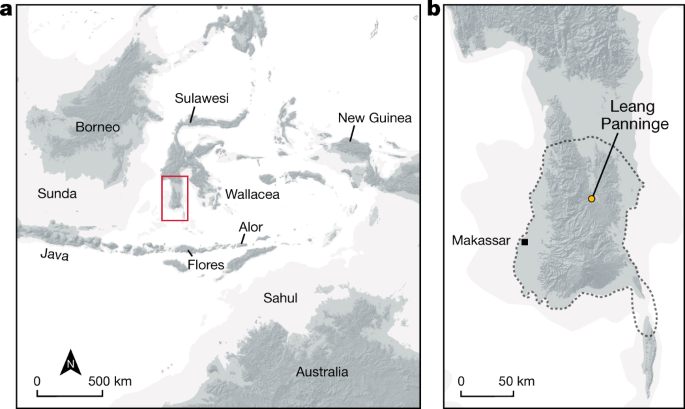
Hasanuddin. Gua Panningnge di Mallawa, Maros: kajian tentang gua hunian berdasarkan artefak batu dan sisa fauna. Naditira Widya 11 , 81–96 (2017).
Artigo Google Scholar
- 3
Bulbeck, D., Pasqua, M. & Di Lello, A. História da cultura de Toalean de South Sulawesi, Indonésia. Perspectiva asiática . 39 , 71–108 (2000).
Artigo Google Scholar
- 4
Perston, YL et al. Um esquema de classificação padronizado para os artefatos Toalean do Holoceno médio de Sulawesi do Sul, Indonésia. PLoS ONE 16 , e0251138 (2021).
CAS PubMed PubMed Central Artigo Google Scholar
- 5
Malaspinas, A.-S. et al. Uma história genômica da Austrália aborígine. Nature 538 , 207–214 (2016).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 6
Pugach,
I., Delfin, F., Gunnarsdóttir, E., Kayser, M. & Stoneking, M. Dados
de todo o genoma substanciam o fluxo gênico do Holoceno da Índia para a
Austrália. Proc. Natl Acad. Sci. USA 110 , 1803–1808 (2013).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 7
Rasmussen, M. et al. Um genoma aborígine australiano revela dispersões humanas separadas na Ásia. Science 334 , 94-98 (2011).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 8
Teixeira, JC & Cooper, A. Using hominin introgression to trace modern human dispersals. Proc. Natl Acad. Sci. USA 116 , 15327–15332 (2019).
CAS PubMed PubMed Central Artigo Google Scholar
- 9
O'Connell, JF et al. Quando o Homo sapiens alcançou o sudeste da Ásia e Sahul pela primeira vez? Proc. Natl Acad. Sci. USA 115 , 8482–8490 (2018).
PubMed PubMed Central Artigo CAS Google Scholar
- 10
Clarkson, C. et al. Ocupação humana do norte da Austrália por 65.000 anos atrás. Nature 547 , 306–310 (2017).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 11
Brumm, A. et al. A arte rupestre mais antiga encontrada em Sulawesi. Sci. Adv . 7 , eabd4648 (2021).
PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 12
Sutikna, T. et al. A
distribuição espaço-temporal de achados arqueológicos e faunísticos em
Liang Bua (Flores, Indonésia) à luz da cronologia revisada do Homo floresiensis . J. Hum. Evol . 124 , 52–74 (2018).
PubMed Artigo Google Scholar
- 13
Samper Carro, SC et al. Em
algum lugar além do mar: restos cranianos humanos das ilhas Lesser
Sunda (Ilha de Alor, Indonésia) fornecem informações sobre o povoamento
do Pleistoceno Superior da Ilha do Sudeste Asiático. J. Hum. Evol . 134 , 102638 (2019).
PubMed Artigo Google Scholar
- 14
Bird, MI et al. O assentamento humano inicial de Sahul não foi um acidente. Sci. Rep . 9 , 8220 (2019).
PubMed PubMed Central Artigo PUBLICIDADES CAS Google Scholar
- 15
Browning,
SR, Browning, BL, Zhou, Y., Tucci, S. & Akey, JM A análise de dados
de sequência humana revela dois pulsos de mistura Denisovan arcaica. Cell 173 , 53-61.e9 (2018).
CAS PubMed PubMed Central Artigo Google Scholar
- 16
Consórcio GenomeAsia100K. O Projeto GenomeAsia 100K permite descobertas genéticas em toda a Ásia. Nature 576 , 106-111 (2019).
Artigo PUBLICIDADES CAS Google Scholar
- 17
Jacobs, GS et al. Múltiplas linhagens denisovanas profundamente divergentes em papuas. Cell 177 , 1010–1021.e32 (2019).
CAS PubMed Artigo Google Scholar
- 18
Ascendência de Qin, P. & Stoneking, M. Denisovan nas populações da Eurásia oriental e dos nativos americanos. Mol. Biol. Evol . 32 , 2665–2674 (2015).
CAS PubMed Artigo Google Scholar
- 19
Reich, D. et al. História genética de um grupo de hominíneos arcaico da caverna Denisova na Sibéria. Nature 468 , 1053–1060 (2010).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 20
Reich, D. et al. Adição de Denisova e as primeiras dispersões humanas modernas no Sudeste Asiático e na Oceania. Sou. J. Hum. Genet . 89 , 516-528 (2011).
CAS PubMed PubMed Central Artigo Google Scholar
- 21
Vernot, B. et al. Escavando DNA de Neandertal e Denisovan de genomas de indivíduos da Melanésia. Science 352 , 235–239 (2016).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 22
Choin, J. et al. Informações genômicas sobre a história da população e adaptação biológica na Oceania. Nature 592 , 583–589 (2021).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 23
Teixeira, JC et al. Ascendência
denisovana difundida na Ilha do Sudeste Asiático, mas nenhuma evidência
de uma mistura substancial de hominídeos superarcaicos. Nat. Ecol. Evol . 5 , 616–624 (2021).
PubMed Artigo Google Scholar
- 24
Mondal,
M., Bertranpetit, J. & Lao, O. Computação Bayesiana aproximada com
aprendizado profundo suporta uma terceira introgressão arcaica na Ásia e
Oceania. Nat. Comun . 10 , 246 (2019).
PubMed PubMed Central Artigo PUBLICIDADES CAS Google Scholar
- 25
Mondal, M. et al. A análise genômica de Andamanese fornece insights sobre a antiga migração humana para a Ásia e sua adaptação. Nat. Genet . 48 , 1066–1070 (2016).
CAS PubMed Artigo Google Scholar
- 26
Lipson, M. et al. Reconstruindo a história da população austronésica na Ilha do Sudeste Asiático. Nat. Comun . 5 , 4689 (2014).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 27
Suryatman, et al. Artefak batu preneolitik situs Leang Jarie: bukti teknologi Maros Point tertua de Kawasan budaya Toalean, Sulawesi Selatan. AMERTA 37 , 1-17 (2019).
Artigo Google Scholar
- 28
Anggraeni, A., Simanjuntak, T., Bellwood, P. & Piper, P. Fundações neolíticas no vale de Karama, West Sulawesi, Indonésia. Antiquity 88 , 740-756 (2014).
Artigo Google Scholar
- 29
Pawlik,
A., Croxier, R., Fuentes, R., Wood, R. & Piper, P. Tradições
funerárias no início do meio-Holoceno Ilha Sudeste Asiático: novas
evidências de Bubog-1, Ilha Ilin, Mindoro Occidental. Antiquity 93 , 901–918 (2019).
Artigo Google Scholar
- 30
Fu, Q. et al. Um ser humano moderno da Romênia com um ancestral Neandertal recente. Nature 524 , 216–219 (2015).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 31
Fu, Q. et al. Uma escala de tempo revisada para a evolução humana com base em genomas mitocondriais antigos. Curr. Biol . 23 , 553–559 (2013).
CAS PubMed PubMed Central Artigo Google Scholar
- 32
Lazaridis, I. et al. Genomas humanos antigos sugerem três populações ancestrais para os europeus atuais. Nature 513 , 409-413 (2014).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 33
Patterson, N. et al. Mistura antiga na história humana. Genetics 192 , 1065–1093 (2012).
PubMed PubMed Central Artigo Google Scholar
- 34
Skoglund, P. et al. Informações genômicas sobre o povoamento do sudoeste do Pacífico. Nature 538 , 510-513 (2016).
PubMed PubMed Central Artigo PUBLICIDADES CAS Google Scholar
- 35
Kanzawa-Kiriyama, H. et al. Genoma nuclear parcial dos Jomons que viveram há 3.000 anos em Fukushima, Japão. J. Hum. Genet . 62 , 213–221 (2017).
CAS PubMed Artigo Google Scholar
- 36
Sikora, M. et al. A história da população do nordeste da Sibéria desde o Pleistoceno. Nature 570 , 182-188 (2019).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 37
Yang, MA et al. O DNA antigo indica mudanças e mistura da população humana no norte e no sul da China. Science 369 , 282-288 (2020).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 38
Yang, MA et al. Um indivíduo de 40.000 anos da Ásia fornece uma visão sobre a estrutura da população inicial na Eurásia. Curr. Biol . 27 , 3202–3208.e9 (2017).
CAS PubMed PubMed Central Artigo Google Scholar
- 39
Meyer, M. et al. Uma sequência de genoma de alta cobertura de um indivíduo Denisovan arcaico. Science 338 , 222-226 (2012).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 40
Bergström, A. et al. Insights sobre a variação genética humana e história populacional de 929 genomas diversos. Science 367 , eaay5012 (2020).
PubMed PubMed Central Artigo CAS Google Scholar
- 41
Peter, BM 100.000 anos de fluxo gênico entre neandertais e denisovanos nas montanhas Altai. Pré-impressão em https://doi.org/10.1101/2020.03.13.990523 (2020).
- 42
Pickrell,
JK & Pritchard, JK Inferência de divisões e misturas de populações a
partir de dados de frequência de alelos em todo o genoma. PLoS Genet . 8 , e1002967 (2012).
CAS PubMed PubMed Central Artigo Google Scholar
- 43
Sikora, M. et al. Genomas antigos mostram o comportamento social e reprodutivo de forrageadoras do início do Paleolítico Superior. Science 358 , 659–662 (2017).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 44
Aubert, M. et al. Arte em cavernas do Pleistoceno de Sulawesi, Indonésia. Nature 514 , 223-227 (2014).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 45
Aubert, M. et al. A cena de caça mais antiga da arte pré-histórica. Nature 576 , 442–445 (2019).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 46
van den Bergh, GD et al. Primeira ocupação hominínea de Sulawesi, Indonésia. Nature 529 , 208-211 (2016).
PubMed Artigo PUBLICIDADES CAS Google Scholar
- 47
Wang, C.-C. et al. Informações genômicas sobre a formação de populações humanas no Leste Asiático. Nature 591 , 413–419 (2021).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 48
Bellwood, P. First Islanders: Prehistory and Human Migration in Island Southeast Asia (Wiley Blackwell, 2017).
- 49.
Mallick, S. et al. The Simons Genome Diversity Project: 300 genomas de 142 populações diversas. Nature 538 , 201–206 (2016).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 50
Fu, Q. et al. A história genética da Idade do Gelo na Europa. Nature 534 , 200–205 (2016).
- 51
Scott, GR & Turner, CG II. A antropologia dos dentes humanos modernos: morfologia dentária e sua variação nas populações humanas recentes (Cambridge Univ. Press, 1997).
- 52
Hillson, S. Dental Anthropology (Cambridge Univ. Press, 1996).
- 53
Pinhasi, R. et al. O DNA antigo ideal é obtido da parte do ouvido interno do osso petroso humano. PLoS ONE 10 , e0129102 (2015).
PubMed PubMed Central Artigo CAS Google Scholar
- 54
Dabney, J. et al. Sequência
completa do genoma mitocondrial de um urso da caverna do Pleistoceno
Médio reconstruída a partir de fragmentos de DNA ultracurtos. Proc. Natl Acad. Sci. USA 110 , 15758–15763 (2013).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 55
Rohland,
N., Harney, E., Mallick, S., Nordenfelt, S. & Reich, D. Partial
uracil-DNA-glycosylase treatment for screening of Ancient DNA. Phil. Trans. R. Soc. Lond. B 370 , 20130624 (2015).
Artigo CAS Google Scholar
- 56
Gansauge,
M.-T., Aximu-Petri, A., Nagel, S. & Meyer, M. Preparação manual e
automatizada de bibliotecas de DNA de fita simples para o sequenciamento
de DNA de restos biológicos antigos e outras fontes de DNA altamente
degradado . Nat. Protoc . 15 , 2279-2300 (2020).
CAS PubMed Artigo Google Scholar
- 57
Kircher,
M., Sawyer, S. & Meyer, M. A indexação dupla supera imprecisões no
sequenciamento multiplex na plataforma Illumina. Nucleic Acids Res . 40 , e3 (2012).
CAS PubMed Artigo Google Scholar
- 58
Peltzer, A. et al. EAGER: eficiente reconstrução do genoma antigo. Genome Biol . 17 , 60 (2016).
PubMed PubMed Central Artigo CAS Google Scholar
- 59.
Li, H. & Durbin, R. Alinhamento de leitura curta rápido e preciso com transformada de Burrows-Wheeler. Bioinformatics 25 , 1754–1760 (2009).
CAS PubMed PubMed Central Artigo Google Scholar
- 60
Peyrégne, S. & Peter, BM AuthentiCT: um modelo de dano de DNA antigo para estimar a proporção de contaminação de DNA atual. Genome Biol . 21 , 246 (2020).
PubMed PubMed Central Artigo CAS Google Scholar
- 61
Renaud,
G., Slon, V., Duggan, AT & Kelso, J. Schmutzi: estimativa de
contaminação e consenso mitocondrial endógeno pedindo DNA antigo. Genome Biol . 16 , 224 (2015).
PubMed PubMed Central Artigo Google Scholar
- 62
Vianello, D. et al. HAPLOFIND: um novo método para atribuição de haplogrupo de mtDNA de alto rendimento. Zumbir. Mutat . 34 , 1189–1194 (2013).
PubMed Artigo Google Scholar
- 63
Katoh,
K., Misawa, K., Kuma, K. & Miyata, T. MAFFT: um novo método para
alinhamento de sequência múltipla rápido com base na transformada rápida
de Fourier. Nucleic Acids Res . 30 , 3059–3066 (2002).
CAS PubMed PubMed Central Artigo Google Scholar
- 64
Kumar,
S., Stecher, G., Li, M., Knyaz, C. & Tamura, K. MEGA X: análise
genética evolutiva molecular em plataformas de computação. Mol. Biol. Evol . 35 , 1547–1549 (2018).
CAS PubMed PubMed Central Artigo Google Scholar
- 65
Skoglund, P. et al. Separando o DNA endógeno antigo da contaminação moderna em um Neandertal siberiano. Proc. Natl Acad. Sci. USA 111 , 2229–2234 (2014).
CAS PubMed PubMed Central Artigo PUBLICIDADES Google Scholar
- 66
Patterson,
N., Richter, DJ, Gnerre, S., Lander, ES & Reich, D. Evidência
genética para especiação complexa de humanos e chimpanzés. Nature 441 , 1103-1108 (2006).
CAS PubMed Artigo PUBLICIDADES Google Scholar
- 67
Chang, CC et al. PLINK de segunda geração: enfrentando o desafio de conjuntos de dados maiores e mais ricos. Gigascience 4 , 7 (2015).
PubMed PubMed Central Artigo CAS Google Scholar
- 68
Narasimhan, VM et al. A formação de populações humanas na Ásia do Sul e Central. Science 365 , eaat7487 (2019).
CAS PubMed PubMed Central Artigo Google Scholar