Tempo e “Idades dos Mamíferos Terrestres”
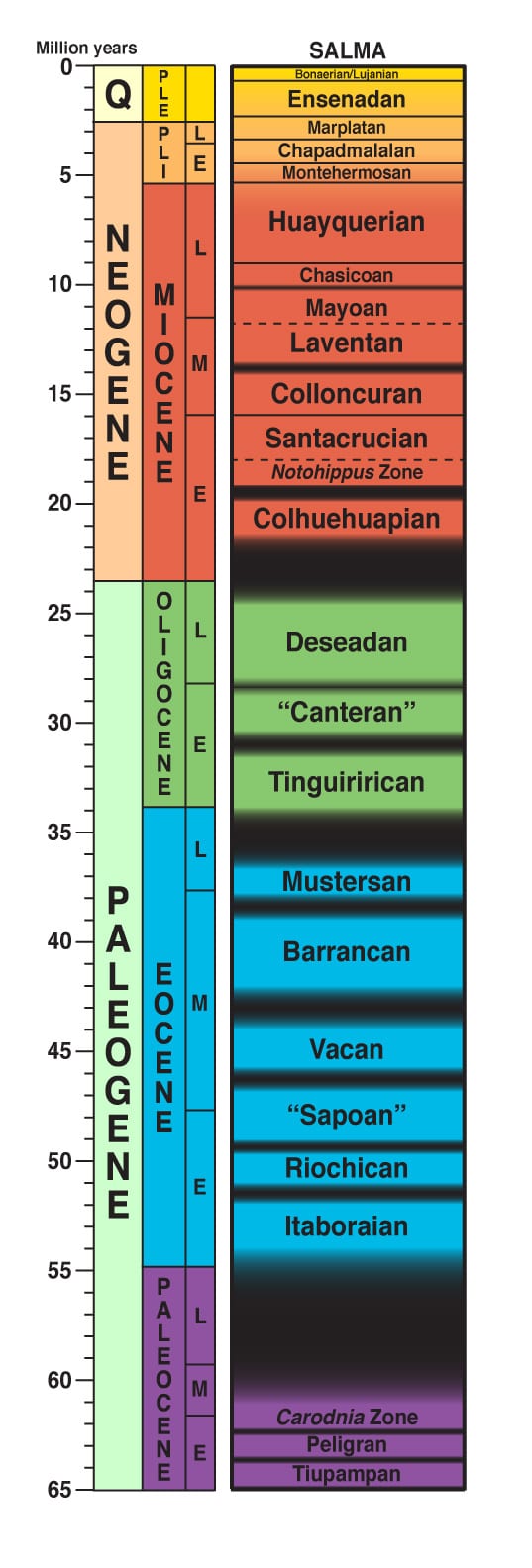
Idades dos mamíferos terrestres da América do Sul (SALMAs) de Croft (2016). Os quatro SALMAs mais recentes são condensados em um único intervalo apenas para fins ilustrativos.
Você não pode falar sobre fósseis sem falar sobre o tempo, e os paleontólogos de mamíferos (também conhecidos como paleomamamologistas) geralmente têm duas maneiras diferentes de falar sobre há quanto tempo um animal viveu.
A primeira maneira é discutir a idade em termos de tempo absoluto: dizer que um fóssil tem 25 milhões de anos, por exemplo. A única maneira de determinar a idade de um fóssil em termos de milhões de anos é datar diretamente as rochas nas quais o espécime foi encontrado usando técnicas de datação radiométrica. Uma idade um pouco menos precisa pode ser determinada datando as rochas acima e abaixo de um fóssil, dando uma gama de idades possíveis: mais de 25 milhões de anos, mas menos de 30 milhões de anos, por exemplo. O problema com a datação direta de rochas é que o método só funciona para certos tipos de rochas, e os fósseis raramente são encontrados nesses tipos de rochas. Mais frequentemente, uma camada de rocha acima e abaixo de um fóssil pode ser datada, fornecendo uma faixa etária. Falar sobre faixas etárias pode ser um pouco complicado, no entanto, os paleontólogos costumam dizer que o fóssil pertence a uma parte específica da tempo geológica: o Período Jurássico Superior ou o início da Época Oligoceno , por exemplo. A maioria dos paleontólogos tem uma ideia de como esses períodos de tempo são organizados e suas idades aproximadas, por isso é uma maneira conveniente de falar sobre o tempo.
Além disso, como todos os paleontólogos (não apenas paleomamologistas) usam a mesma escala de tempo, é fácil discutir a idade dos eventos com cientistas de outras partes do mundo ou com aqueles que estudam diferentes tipos de organismos. Em alguns casos, a faixa etária resultante é tão grande que é essencialmente sem sentido. (Uma faixa de 5 a 25 milhões de anos não é muito útil para um paleomamologista.) Outras vezes, as rochas acima e abaixo de um fóssil não podem ser datadas, ou rochas datáveis estão presentes, mas os pesquisadores não tem tempo, dinheiro e/ou experiência para fazer a datação radiométrica. Então o que você faz?
O que os paleomamamologistas costumam fazer é referir o fóssil (ou a fauna) ao que é conhecido como “idade” de um mamífero terrestre. As idades dos mamíferos terrestres podem ser consideradas como um sistema alternativo para descrever a idade de algo; o sistema é baseado na observação de que faunas de diferentes idades são compostas por diferentes espécies de mamíferos. Cada idade de mamífero terrestre tende a ser caracterizada por uma ou mais espécies de mamíferos (ou uma combinação de espécies de mamíferos) que são exclusivas daquele período de tempo. Examinando um grande número de faunas de diferentes idades, toda uma escala de tempo pode ser erguida com base na sucessão de espécies de mamíferos. Cada unidade na escala de tempo é conhecida como idade dos mamíferos terrestres.
Como exemplo, pense em encontrar um depósito de ossos fósseis que inclui um osso do braço de um mamute colombiano ( Mammuthus columbi ), o dente de um gato dente-de-sabre ( Smilodon fatalis ) e o crânio de um lobo gigante ( Canis dirus ). Sabemos que esses animais estão extintos agora, mas que estavam vivos há pouco tempo. Sem precisar datar esses ossos radiometricamente, podemos dizer com bastante confiança que eles pertencem à “Era” dos Mamíferos Terrestres Rancholabrean (cerca de 250.000 a 11.000 anos atrás); esse período de tempo é exemplificado pelas espécies de mamíferos que são preservadas em La Brea Tar Pits , na Califórnia. Da mesma forma, se formos para o ermo de Dakota do Sul e encontrarmos a mandíbula fossilizada de um grande titanothere ( Brontops sp. a 34 milhões de anos); essa idade é caracterizada, entre outras coisas, pela presença de algumas das maiores e mais impressionantes espécies desses titanoteres.
O único problema com as idades dos mamíferos terrestres é que elas não são aplicáveis em todos os continentes. Pense na Austrália, por exemplo. Não há um mamífero nativo da Austrália que também seja nativo da América do Norte, e tem sido assim há dezenas de milhões de anos. Assim, não há como saber se duas faunas desses continentes têm a mesma idade com base apenas nas espécies de mamíferos representadas. Devido a este problema geográfico, existe um sistema separado de idades de mamíferos terrestres para cada continente. O conjunto de idades dos mamíferos terrestres com o qual estamos mais preocupados, é claro, é a sequência da “idade” dos mamíferos terrestres da América do Sul (SALMA).
Na América do Sul, o registro fóssil cenozoico foi dividido em aproximadamente 20 dessas “idades” de mamíferos terrestres (veja a figura no canto superior esquerdo). A maioria deles foi tradicionalmente baseada em localidades fósseis em uma área do sul da América do Sul conhecida como Patagônia. A grande extensão latitudinal da América do Sul e sua grande proporção localizada nos trópicos equatoriais (cerca de 70% da área do continente) tornaram desafiadora a integração de localidades fósseis do norte da América do Sul na sequência SALMA tradicional; estimar o tempo com base na similaridade da fauna é difícil quando diferentes espécies de animais vivem nas partes norte e sul de um continente. Este problema de correlação foi agravado pela falta de datas radiométricas ou paleomagnéticas (ou seja, datas “absolutas”) para a maioria das faunas e a presença de lacunas de tempo significativas no registro fóssil. Avanços recentes nas técnicas de datação radiométrica (por exemplo, datação por fusão a laser 40 Ar/ 39 Ar) e a descoberta de novas e importantes localidades fósseis cenozoicas dentro de camadas de rochas datáveis (como o Tinguiririca Fauna do Chile) ajudaram a melhorar nossa compreensão das relações entre esses SALMAs.
Referências citadas:
- Croft, DA 2016. Tatus Horned e Macacos Rafting: Os Fascinantes Mamíferos Fósseis da América do Sul . Indiana University Press, Bloomington, Indiana, 320 pp.







:max_bytes(150000):strip_icc():format(webp)/GettyImages-1147987049-471ab72f8b684f73ae27ee1dd9763a68.jpg)