Complex multicellularity
requires elaborate developmental mechanisms, often based on the
versatility of heterodimeric transcription factor (TF) interactions.
Homeobox TFs in the TALE superclass are deeply embedded in the gene
regulatory networks that orchestrate embryogenesis. Knotted-like
homeobox (KNOX) TFs, homologous to animal MEIS, have been found to drive
the haploid-to-diploid transition in both unicellular green algae and
land plants via heterodimerization with other TALE superclass TFs,
demonstrating remarkable functional conservation of a developmental TF
across lineages that diverged one billion years ago. Here, we sought to
delineate whether TALE-TALE heterodimerization is ancestral to
eukaryotes.
Results
We analyzed TALE endowment in
the algal radiations of Archaeplastida, ancestral to land plants.
Homeodomain phylogeny and bioinformatics analysis partitioned TALEs into
two broad groups, KNOX and non-KNOX. Each group shares previously
defined heterodimerization domains, plant KNOX-homology in the KNOX
group and animal PBC-homology in the non-KNOX group, indicating their
deep ancestry. Protein-protein interaction experiments showed that the
TALEs in the two groups all participated in heterodimerization.
Conclusions
Our study indicates that the
TF dyads consisting of KNOX/MEIS and PBC-containing TALEs must have
evolved early in eukaryotic evolution. Based on our results, we
hypothesize that in early eukaryotes, the TALE heterodimeric
configuration provided transcription-on switches via
dimerization-dependent subcellular localization, ensuring execution of
the haploid-to-diploid transition only when the gamete fusion is
correctly executed between appropriate partner gametes. The TALE switch
then diversified in the several lineages that engage in a complex
multicellular organization.
The
homeobox transcription factors (TFs) are ubiquitous in eukaryotes,
carrying a DNA-binding homeodomain typically 60 amino acids that folds
into three α-helices [1].
The atypical or TALE (three-amino acid length extension) superclass of
homeobox TFs shares a three-amino-acid insertion between helix 1 and 2
and plays essential roles during embryonic development by participating
in interactive TF networks. In animals, MEIS- and PBC-class TALE
proteins, such as Meis/Hth and Pbx/Exd, form heterodimers that in turn
form ternary complexes with HOX-class homeobox TFs, determining cellular
fates along the anterior-posterior axis of the developing embryo [2, 3].
In plants, the interacting KNOX- and BELL-class TFs in the TALE group
play critical roles during organ formation and the
vegetative-to-reproductive transition in the undifferentiated cell mass
known as the shoot apical meristem [4, 5].
The
heterodimerization of TALE proteins serves as a trigger for precise
execution of developmental programs. Prior to heterodimerization, animal
PBX proteins are localized in the cytosol, and upon binding to MEIS,
they translocate to the nucleus [6, 7]. Similar heterodimerization-dependent translocation is also observed for KNOX-BELL pairs in the plant Arabidopsis, implying that this mechanism is a conserved regulatory feature of TALE proteins [8]. In addition, TALE proteins differ in their DNA-binding specificity [9, 10], which is primarily determined by the homeodomain residues at positions 47, 50, and 54 [11], and heterodimerization increases target affinity by bringing two such DNA-binding domains together.
TALE-heterodimerization
is mediated by class-specific homology domains located on the
N-terminal side adjacent to the homeodomain [12, 13].
Animal MEIS and plant KNOX class proteins share readily identifiable
homology in their heterodimerization domain, leading to the proposal of
an ancestral TALE class named MEINOX [12].
In contrast, their partner classes—PBC and BELL—exhibit no apparent
sequence similarity in their heterodimerization domains. Short shared
sequence motifs and common secondary structures have been found within
the heterodimerization domains between MEINOX and PBC or BELL [14, 15], but their extent of the homology requires adequate taxon sampling to recover ancestral relationships.
An ancestral function of TALE-TALE heterodimerization was revealed in studies of the unicellular green alga Chlamydomonas reinhardtii:
the KNOX ortholog GSM1 and a second TALE protein GSP1 form heterodimers
immediately after the fusion of sexual gametes, and these drive the
haploid-to-diploid transition by activating > 200 diploid-specific
genes and inactivating > 100 haploid-specific genes [10, 16, 17].
In subsequent studies, plant-type TALE-TALE heterodimers between KNOX
and BELL were shown to be required for the haploid-to-diploid transition
of the moss Physcomitrella patens [18, 19].
Given the conserved role of TALE heterodimerization as a developmental
switch in the sexual life cycle of the plant lineage, understanding its
origins and diversification promises to shed light on the evolution of
developmental mechanisms during eukaryotic radiation and the emergence
of land plants.
The Archaeplastida consists of three monophyletic phyla [20, 21] (Fig. 1). (1) Viridiplantae include two divisions, Chlorophyta—chlorophytes and prasinophytes (a paraphyletic group of seven lineages [22])—and Streptophyta—charophyte algae and land plants [23]. (2) Rhodophyta (red algae) include diverse unicellular and multicellular organisms that diverge into four major lineages [24] (Additional file 1:
Table S1). (3) Glaucophyta members include only four cultured genera
and possess plastids that carry ancestral features of the cyanobacterial
symbiont that gave rise to photosynthetic organelles in eukaryotes [25].
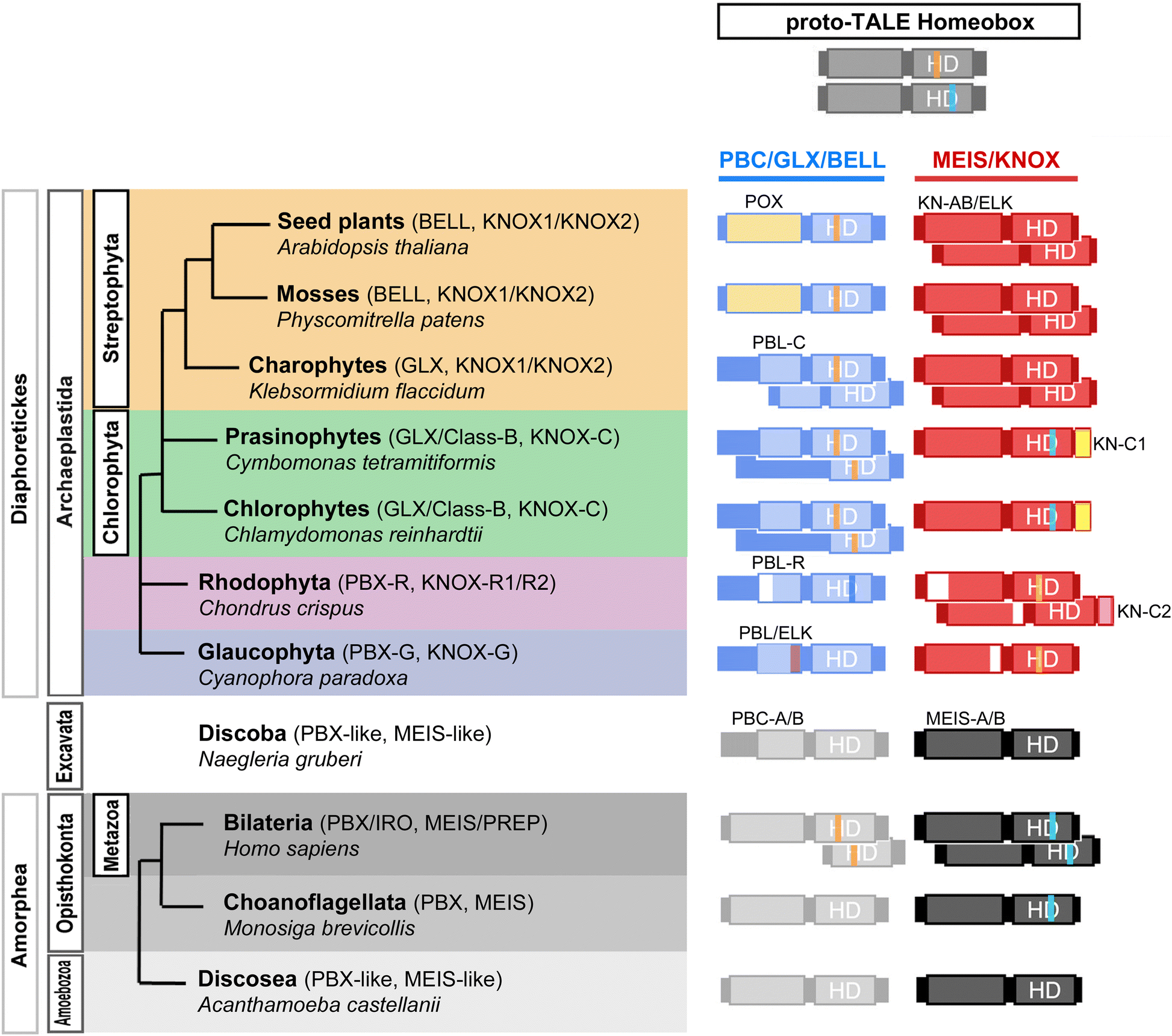
Fig. 1
Common origin of
heterodimerizing TALE homeobox TFs. Hypothesized homodimerizing
proto-TALE protein (top) duplicated before the eukaryotic radiations
into animals/fungi/amoebae vs. algae/plants. Lineage-specific
diversification soon followed, generating heterodimeric configurations
distinct at the phylum-level. (Left) Each lineage possesses one or two
classes of potential heterodimeric TALEs, which are summarized onto the
eukaryotic phylogeny. A representative species name is given for each
analyzed lineage. (Right) Summary of TALE configurations, coupling
members of the PBC/PBX/GLX group that shares PBC-homology domains and of
the MEIS/KNOX group that shows homology in the KN-A/B domains
N-terminal to the homeodomain. Lightly shaded boxes depict homology
domains, whose names are provided above. Open areas in the domain boxes
indicate the absence of MEINOX-motif for PBX-Red, KN-A for KNOX-Red1 and
ELK for KNOX-Red2. Colored vertical lines in the HD indicate two shared
introns at 44/45 (orange over “H” in HD) and 48(2/3) (blue over “D” in
HD), whose alternating existence between the two groups suggests
independent diversification of TALE heterodimerization. HD: Homeodomain;
PBL-C: PBL-Chloro; PBL-R: PBL-Red
To
delineate the ancestry of plant-type TALE heterodimerization, we
performed a phylogenetic and bioinformatics analysis of TALE TFs in the
three algal radiations of the Archaeplastida supergroup, the descendants
of a single endosymbiosis event greater than one billion years ago [26, 27].
Our analysis showed that the TALEs were already diversified into two
groups at the origin of Archaeplastida, one sharing KNOX-homology and
the other sharing PBC-homology. Together with our protein-protein
interaction data, we propose that all TALE classes participate in
heterodimerization networks via the KNOX- and PBC-homology domains
between the two ancestral groups.
TALEs in Archaeplastida are divided into two groups, KNOX and non-KNOX
To
collect all the available homeobox protein sequences, we performed
BLAST and Pfam-motif searches against non-plant genomes and
transcriptome assemblies throughout the Archaeplastida, identifying 338
proteins from 56 species as the Archaeplastida homeobox collection (30
genomes and 18 transcriptomes; Additional file 1:
Table S1). Of these, 104 possessed the defining feature of TALE
proteins, a three-amino-acid insertion between aa positions 23–24 in the
homeodomain [28].
At least two TALE genes were detected in most genomes except five
genomes in the Trebouxiophyceae class of the Chlorophyta
(Additional file 1: Table S1; see Additional file 2: Note S1 for further discussion of the absence of TALEs in Trebouxiophyceae).
The
collected TALE sequences were then classified by their homeodomain
features using a phylogenetic approach, with TALEs from animals, plants,
and early-diverging eukaryotes (Amoebozoa and Excavata) as outgroups
(Additional file 3: Figure S1). The resultant TALE homeodomain phylogeny distinguished two groups in all three phyla of Archaeplastida (Fig. 2).
(1) The KNOX-group as a well-supported clade displayed a
phylum-specific cladogram: two Glaucophyta sequences at the base (as
KNOX-Glauco) were separate from the next clade, which combines
Rhodophyta sequences (as KNOX-Red1) and a Viridiplantae-specific clade
with strong support (92/90/1.00). (2) The non-KNOX group, including the
BELL and GSP1 homologs, contained clades of mixed taxonomic
affiliations. These analyses showed that the TALE proteins had already
diverged into two groups before the evolution of the Archaeplastida and
that the KNOX-group is highly conserved throughout Archaeplastida.
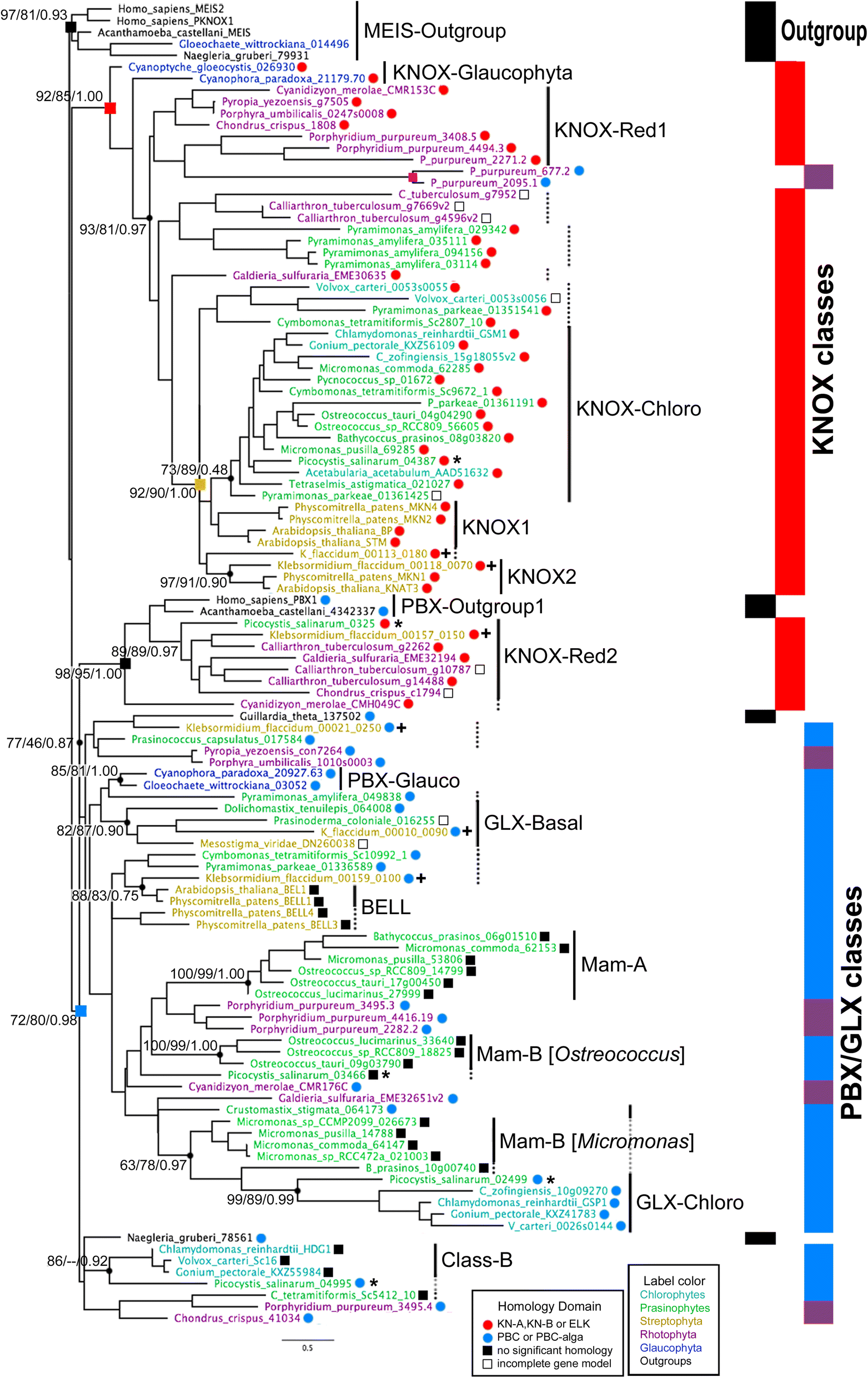
Fig. 2
Maximum
likelihood (ML) phylogeny of the TALE superclass homeodomain in
Archaeplastida supports ancient division between KNOX- and non-KNOX TALE
groups. The ML trees were generated from the homeodomain alignment with
70 amino acid positions. The consensus tree out of 1000 bootstrap trees
is shown. The three numbers at critical nodes show %bootstrap, %SH, and
Bayesian posterior probability in support of clades. The tree contains
two outgroup clades marked by black squares at nodes, and two
Archaeplastida clades, one combining most KNOX sequences marked by the
red square and the other combining all non-KNOX sequences marked by the
blue square. Vertical bars on the right depict the distribution of
outgroup in black, KNOX in red, and non-KNOX sequences in blue. Red dots
by the sequence names indicate the presence of KN-A or KN-B domains,
and blue dots indicate the presence of a PBC-homology domain. Truncated
sequences not available for homology domain analysis are marked with
open black boxes. Filled black boxes indicate the absence of a KN-A/B or
PBC-homology domain. Proposed classification is indicated by the
vertical lines. Dotted vertical lines indicate suggested class members
placed outside the main clade for the class in the phylogeny. PBX-Red
sequences are found in four separate clades, marked by purple shades on
the blue section of the vertical bars. Two PBX-Red sequences marked by
the purple square are exceptionally found in the KNOX-Red1 clade, having
divergent amino acids at highly conserved positions at Trp19, His23,
and Lys31 in their homeodomain, suggesting their false association with
the KNOX-Red1. Colors of the sequence names indicate their phylogenetic
group: Blue for Glaucophyta, purple for Rhodophyta, green for
prasinophytes, light blue for the chlorophytes, orange for Streptophyta,
and black for outgroups. The ruler shows genetic distance. Details of
the sequences analyzed by this phylogeny are provided in
Additional file 1:
Table S2. *Gloeochaete_wittrockiana_014496 is considered as a sequence
from a bannelid-type amoeba that contaminated the original culture
(SAG46.84) for the MMETSP1089 transcriptome. **Association of KNOX-Red2
class sequences to Amorphea PBC sequences is attributed to a shared WFGN
motif determining DNA-binding specificity of the homeodomain via
convergent evolution
KNOX group sequences share the same heterodimerization domains throughout Archaeplastida
The
next question was whether the plant KNOX class originated prior to the
Viridiplantae phylum. The plant KNOX proteins and the Chlorophyta GSM1
possess the KNOX-homology, consisting of KN-A, KN-B, and ELK domains,
required for their heterodimerization with other TALE proteins [10];
therefore, the presence of the KNOX homology would suggest the
potential for heterodimerization to the KNOX group. To collect homology
domains without prior information, we performed ad-hoc homology domain
searches among the KNOX group sequences. Using the identified homology
domains as anchors, we carefully curated an alignment of the KNOX-group
sequences combined with any other TALE sequences with a KNOX-homology,
(Additional file 3:
Figure S2). From this KNOX alignment, we found all KNOX group sequences
(excluding partial sequences) showing amino acid similarity scores
> 50% for at least two of the three domains comprising the
KNOX-homology region (Additional file 1:
Table S3 for calculated domain homology). To test whether the observed
similarity is specific to the TALE sequences, we generated HMM motifs
for KN-A and KN-B domains from the KNOX alignment, searched them in the
target genomes, and confirmed that KN-A and KN-B domains are found only
in the TALE sequences (Additional file 4: Data S1 and S2). We thereby defined KNOX-homologs as the TALE sequences possessing searchable KNOX homology (Fig 2,
marked by red dots following their IDs), suggesting that the
KNOX-homolog already existed before the evolution of eukaryotic
photosynthesis as represented by the Archaeplastida.
In
addition to the KNOX-homology, the same search also revealed two novel
domains at the C-terminus of the homeodomain (Additional file 3:
Figure S2): the first (KN-C1) was shared among the Chlorophyta
sequences, and the second (KN-C2) was shared among a group of KNOX
homologs in a clade outside the KNOX-group (KNOX-Red2).
KNOX classes diverged independently among the algal phyla
In
Viridiplantae, we found a single KNOX homolog in most Chlorophyta
species, whereas KNOX1 and KNOX2 divergence was evident in the
Streptophyta division, including the charophyte Klebsormidium flaccidum and land plants (Fig. 2). The newly discovered KN-C1 domain was specific to the Chlorophyta KNOX sequences and found in all but one species (Pyramimonas amylifera).
The absence of similarity between KN-C1 and the C-terminal extensions
of KNOX1/KNOX2 sequences suggests independent, lineage-specific KNOX
evolution in the Chlorophyta and Streptophyta (Additional file 3:
Figure S2). We, therefore, refer to the Chlorophyta KNOX classes as
KNOX-Chloro in contrast to the KNOX1 and KNOX2 classes in the
Streptophyta.
The
KNOX homologs in the Rhodophyta were divided into two classes: a
paraphyletic group close to the KNOX-Chloro clade, named KNOX-Red1, and a
second group near the PBX-Outgroup, named KNOX-Red2. KNOX-Red1 lacked a
KN-A, whereas KNOX-Red2 lacked an ELK and shared a KN-C2 domain
(Additional file 3:
Figure S2). We consider KNOX-Red1 as the ancestral type, since the
KNOX-Red1 sequences were found in all examined Rhodophyta taxa, whereas
the KNOX-Red2 sequences were restricted to two taxonomic classes
(Cyanidiophyceae and Florideophyceae). Interestingly, the KNOX-Red2
clade included two green algal sequences, with strong statistical
support (89/89/0.97; Fig. 2); these possessed a KN-C2 domain, suggesting their ancestry within the KNOX-Red2 class (Additional file 3: Figure S2; see Additional file 2: Note S2 for further discussion about their possible origin via horizontal gene transfer).
Available
TALE sequences were limited for the Glaucophyta. We found a single KNOX
homolog in two species, which possessed KN-A and KN-B domains but
lacked an ELK domain. We termed these KNOX-Glauco.
Non-KNOX group TALEs possess animal type PBC-homology domain, suggesting a shared ancestry between Archaeplastida and Metazoa
Following
the identification of KNOX homologs, the remaining TALE sequences were
combined as the non-KNOX group that lacks KN-A and KN-B domains in
Archaeplastida. Further classification of the non-KNOX group was
challenging due to its highly divergent homeodomain sequences. However,
we noticed that the number of non-KNOX genes per species was largely
invariable: one in most Rhodophyta and Glaucophyta genomes and two in
the majority of Chlorophyta genomes, suggesting their conservation
within each radiation.
Our
ad-hoc homology search provided critical information for non-KNOX
classification, identifying a homology domain shared among all
Glaucophyta and Rhodophyta non-KNOX sequences (Fig. 3a, b).
Since this domain showed a similarity to the second half of the animal
PBC-B domain (Pfam ID: PF03792) known as heterodimerization domain [12],
we named this domain PBL (PBC-B Like). Accordingly, we classified all
the non-KNOX TALEs in Glaucophyta and Rhodophyta as a single PBC-related
homeobox class, PBX-Glauco or PBX-Red. PBX-Glauco sequences also
possessed the MEINOX motif, conserved in the animal PBC-B domain,
indicating common ancestry of PBC-B and PBL domains (Fig. 3a).
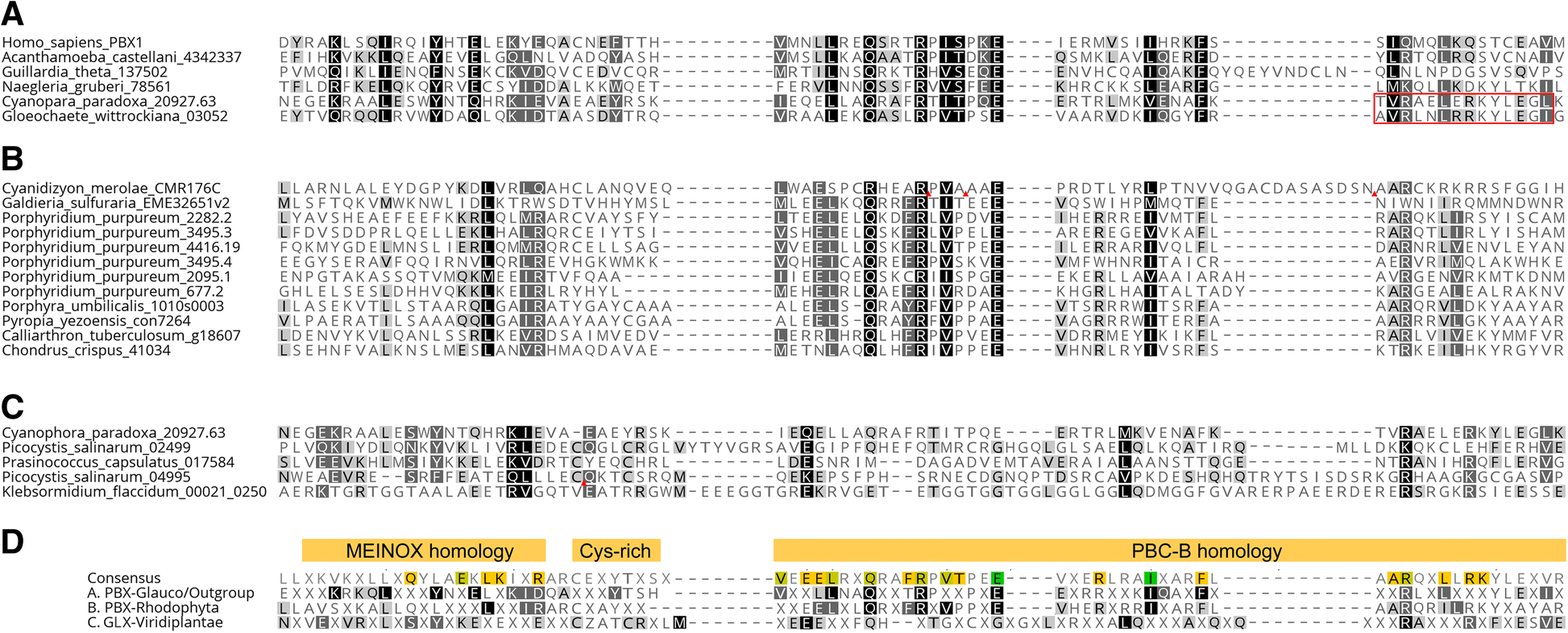
Fig. 3
Archaeplastida
non-KNOX group TALEs possess a PBC-like domain (PBL) consisting of
N-terminal MEINOX homology and C-terminal PBC-B homology. Amino acid
letters in black with gray shades, in white with light shades, and in
white with black shades show more than 60%, 80%, or 100% similarity in
each column. Inverse red triangles indicate the discarded sequences in
un-aligned insertions. a
PBL-Glauco domain alignment, including two Glaucophyta sequences sharing
homology in both MEINOX homology and C-terminal half of the PBC-B
domain with non-Archaeplastida TALE sequences. Red box indicates the ELK
domain. b PBL-Red domain alignment. All Rhodophyta non-KNOX sequences possess a PBL domain with poor MEINOX homology. c
PBL-Chloro domain alignment. Cyanophora_paradox_20927.63 is included
for comparison. Picocystis_salinarum_02499 is a founding member of GLX
class with a PBL-Chloro domain. d Comparison among PBL domains. The top row shows the consensus made from the alignment of (a), (b), and (c) combined and the lower consensus sequences are collected from the individual alignments presented in (a), (b), and (c)
GSP1 shares distant PBC-homology together with other non-KNOX group sequences in Viridiplantae
A
remaining question was the evolution of the Chlorophyta non-KNOX
sequences that apparently lacked a PBC-homology. To uncover even a
distant homology, we compared the newly defined PBL domains with the
Chlorophyta sequences by BLAST (cut-off E-value
of 1E-1) and multiple sequence alignments. This query collected three
prasinophyte and one charophyte TALE sequences that possessed a MEINOX
motif and a putative PBL-domain; however, they showed very low sequence
identity among themselves (Fig. 3c).
Further query utilizing these four sequences identified 11 additional
non-KNOX sequences. Nine of these were made into two alignments, one
including GSP1 homologs and the other combining most prasinophyte
sequences (Additional file 3:
Figure S3). The two remaining sequences (Picocystis_salinarum_04995 and
Klebsormidium_flaccidum_00021_0250) showed a homology to a PBX-Red
sequence of Chondrus cruentum
(ID:41034) in a ~ 200 aa-long extension beyond the PBL domain,
suggesting their PBX-Red ancestry (another potential case of horizontal
transfers; Additional file 3:
Figure S4). All the Chlorophyta non-KNOX sequences that carry the
PBL-homology domains were classified as GLX (GSP1-like homeobox) in
recognition of the GSP1 protein of Chlamydomonas as the first characterized member of this class [29].
Is the plant BELL class homologous to the Chlorophyta GLX class?
The BELL class is the only non-KNOX class in land plants, sharing a POX (Pre-homeobox) domain (PF07526) [13] and lacking an identifiable PBL domain. The K. flaccidum
genome, one of the two genomes available in the charophyte from which
land plant emerged, contained three non-KNOX sequences, all possessing a
PBL domain (Fig. 3, Additional file 3: Figures S3, S4). The second charophyte genome of Chara braunii
contained one putative BELL homolog that appears to be truncated for
the N-terminal sequences outside its C-terminal homeodomain possibly due
to the incomplete gene model. Therefore, the lack of PBL-homology in
the plant BELL class appears to be due to divergence or domain loss from
an old charophyte class that had PBL-homology. We found an intron at
the 24(2/3) homeodomain position of a K. flaccidum GLX homolog, which was previously identified as being specific to the plant BELL class (Additional file 3: Figure S5) [12],
suggesting that the plant BELL class evolved from an ancestral GLX
gene. More taxon sampling in charophytes is needed to confirm this
inference.
Two non-KNOX paralogs of Chlorophyta heterodimerize with the KNOX homologs
Even
with our sensitive iterative homology search, we could not identify a
PBC/PBL-homology in about half of the Chlorophyta non-KNOX sequences.
Since most Chlorophyta genomes possess one GLX homolog and one non-KNOX
sequence without the PBL-homology domain, we refer the latter
collectively to Class-B (Additional file 3:
Figure S6). Exceptions were found in one prasinophyte clade (class
Mamiellophyceae), whose six high-quality genomes all contain two
non-KNOX sequences lacking the PBL-homology. Nonetheless, these non-KNOX
sequences formed two groups, one more conserved and the other less
conserved and polyphyletic, referred to the Mam-A and Mam-B classes,
respectively (Additional file 3: Figures S7, S8). Considering the reductive genome evolution of the Mamiellophyceae [30], the conserved Mam-A class may be derived from an ancestral GLX class.
Two
divergent non-KNOX classes in Chlorophyta led to a critical question
about their dyadic networks. Previously studies had shown that TALE
heterodimers required interaction between MEIS and PBC domains in
animals and between KNOX and PBL domains in Chlamydomonas [6, 10].
It was, therefore, predicted that all Glaucophyta and Rhodophyta TALEs
form heterodimers via their KNOX- and PBL-homology domains. On the other
hand, it remained to be tested whether the Chlorophyta TALEs lacking a
PBL-domain can form heterodimers with other TALEs.
To
characterize the interaction network of TALE class proteins in
Chlorophyta, we selected three prasinophyte species for protein-protein
interaction assays: two species containing Mam-A and Mam-B genes (Micromonas commoda and Ostreococcus tauri), and another species (Picocystis salinarum),
whose transcriptome contained one GLX and one Class-B sequence. In all
three species, we found that KNOX homologs interacted with all examined
non-KNOX proteins in Mam-A, Mam-B, Class-B, and GLX class (Fig. 4a–c). No interaction was observed between the two non-KNOX proteins in any of the three species (Fig. 4a–c).
Similar to the GLX-KNOX heterodimerization, Mam-A and Mam-B also
required additional domains outside the homeodomain for their
heterodimerization with the KNOX homologs (Additional file 3:
Figure S9). These results showed that the all divergent non-KNOX TALEs
maintained their original activity to form heterodimers with the KNOX
homologs. Observed interacting network among the TALE sequences is
summarized in Additional file 3: Figure S10.
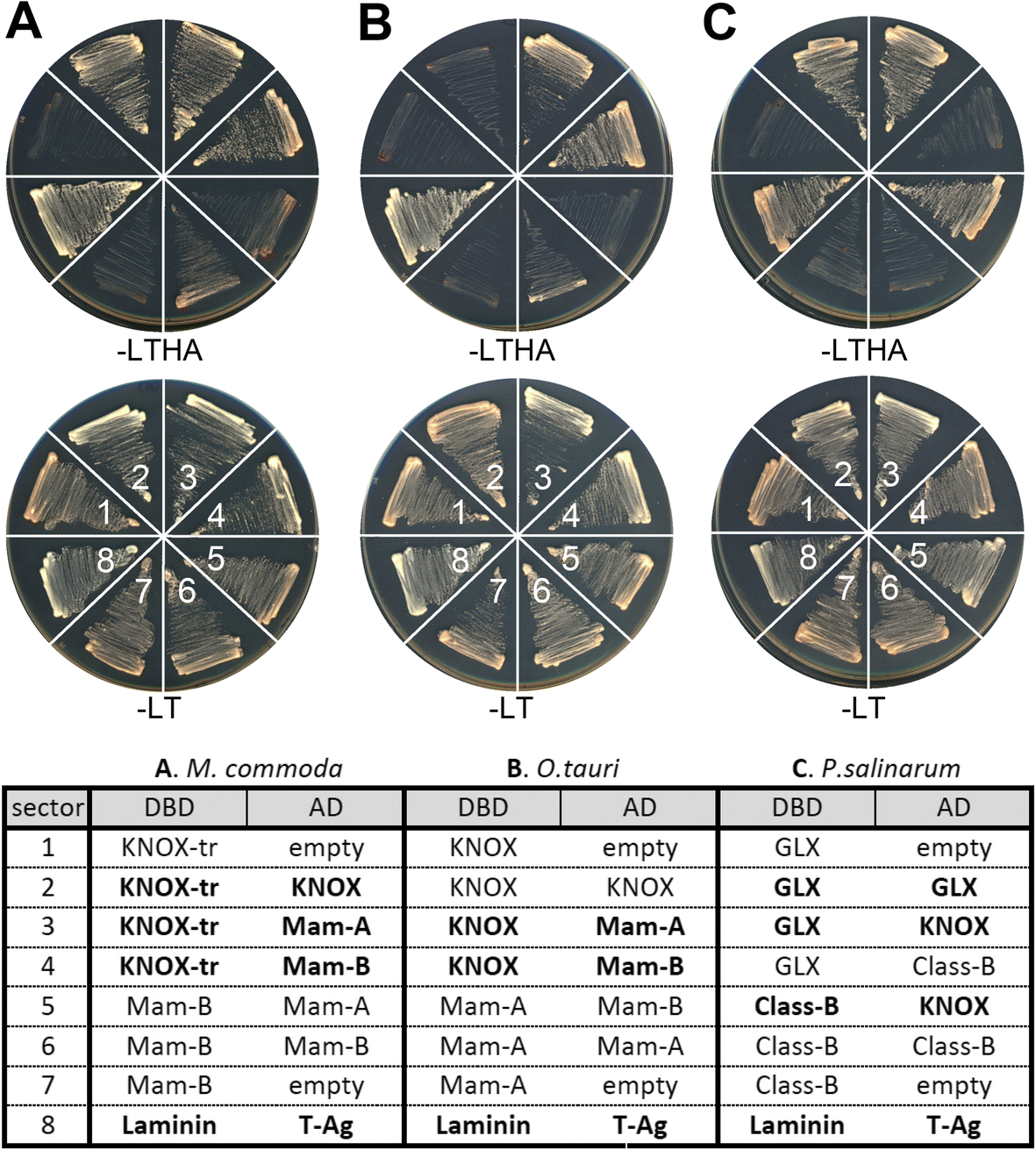
Fig. 4
TALE TFs engage
in heterodimerization networks between KNOX and non-KNOX groups. The
bait constructs conjugated to the GAL4 DNA-binding domain (DBD) and the
prey constructs conjugated to the GAL4 transcriptional activation domain
(AD) are listed in the table. Construct combinations, numbered 1–8, are
arranged in wedges clock-wise, starting at 9 o’clock as labeled in the
-LT panels. Confirmed interacting pairs are shown in bold faces in the
table. The laminin and T-Antigen (T-Ag) pair, known to be interacting
partners, was plated in the 8th sector as a positive control. a Assays using M. commoda TALEs. b Assays using O. tauri TALEs. c Assays using P. salinarum TALEs. KNOX-tr refers to the N-terminal truncated KNOX construct for preventing self-activation. d Detailed construct information is provided in Additional file 1: Table S5
TALE heterodimerization evolved early in eukaryotic history
Our
discovery of the PBC-homology in Archaeplastida suggests common
ancestry of the heterodimerizing TALES between Metazoa and
Archaeplastida. It also predicted that other eukaryotic lineages might
possess TALEs with the PBC-homology. Outside animals, the Pfam database
contains only two PBC-B domain-harboring sequences, one from a
Cryptophyta species (Guillardia theta, ID:137502) and the other from an Amoebozoa species (Acanthamoeba castillian, ID:XP_004342337) [31]. We further examined the Excavata group, near to the posited root of eukaryotic phylogeny [20]. A search of two genomes (Naegleria gruberi and Bodo saltans) collected 12 TALE homeobox sequences in N.gruberi, and none in B.saltans, of which we found one with a PBC-homology domain (ID:78561, Fig. 3a) and one with a MEIS/KNOX-homology (ID:79931, Additional file 3:
Figure S2). We searched additional genomes in the Amorphea and found
the PBC-homology and MEIS/KNOX-homology in the TALE sequences collected
from Apusozoa, Ichtyhosporea, and Choanoflagellata but not from Fungi
(Additional file 3:
Figures S11-S14). Our data suggest that the heterodimerization
domains—the PBC-homology and MEIS/KNOX-homology—originated early in
eukaryotic evolution and persisted throughout the major eukaryotic
radiations.
Intron-retention supports the parallel evolution of the heterodimeric TALE classes during eukaryotic radiations
The
ubiquitous presence of dyadic TALEs raised next question: Are all the
dyadic TALEs reported in this study the descendants of a single
ancestral dyad, or do they result from lineage-specific evolution from a
single prototypical TALE (proto-TALE) that does not engage in
heterodimerization. To probe deep ancestry, we examined
intron-retention, this being regarded as a long-preserved character and
less prone to occur by homoplasy (a character displayed by a set of
species but not present in their common ancestor) [32].
Five intron positions were shared by at least two TALE classes, of
which the 44/45 and 48(2/3) introns qualified as the most ancestral
since they were found throughout the Archaeplastida and Metazoa
(Additional file 3: Figure S5).
The
44/45 and 48(2/3) introns showed an intriguing exclusive distribution
between the two dyadic partners of each phylum: one possesses the 44/45
and the other possesses the 48(2/3) intron (Additional file 3:
Figure S5). This mutually exclusive pattern suggested that two TALE
genes with distinct intron positions existed at the onset of the
eukaryotic radiation. We consider the 44/45 intron position as the most
ancestral, given that it was conserved in most non-TALE homeobox genes [12].
In this regard, we speculate that acquisition of the 48(2/3), and loss
of the 44/45 intron, accompanied an early event wherein the proto-TALE
with the 44/45 intron was duplicated to generate a second TALE with the
48(2/3) intron. Since the 48(2/3) intron position was found within the
KNOX/MEIS group genes in Viridiplantae and Metazoa and also in the PBX
group genes in Rhodophyta and Cryptophyta, we may speculate that the
duplicated TALEs arose early and diversified to establish
lineage-specific heterodimeric configurations during eukaryotic
radiations. Alternatively, the 48(2/3) intron position in the TALE
homeodomain might have been acquired many times during eukaryotic
radiations.
Given
that the heterodimeric TALEs evolved in a lineage-specific manner, we
asked what the proto-TALE looked like at the time it underwent
duplication. The following observations suggest that the proto-TALE was a
homodimerizing protein. First, the PBC-homology domains of PBX/GLX
class proteins identified in the Archaeplastida includes the
MEINOX-motif that was originally defined for its similarity to the
MEIS/KNOX-homology domains (Fig. 3) [14]. Second, PBX-Glauco sequences possess the ELK-homology within their PBL domain (Fig. 3), which align well to the ELK domains of KNOX class sequences in Viridiplantae (Additional file 3:
Figure S15). Therefore, the MEINOX-motif and ELK-homology across the
heterodimerizing KNOX and PBX groups supported the common origin of
heterodimerizing TALE groups from a single TALE by duplication followed
by subfunctionalization.
Our
study shows that all three Archaeplastida phyla possess TALEs, diverged
into two groups with distinct heterodimerization domains, the KNOX
group with KN-A/KN-B domains and the PBX (or GLX) group with PBL
domains. The similarity between the KNOX/PBX and the animal MEIS/PBC
dyads led us to identify homologous heterodimerization domains in the
TALEs of other eukaryotic lineages including Excavata. Based on our
findings, we hypothesize that the TALE heterodimerization arose very
early in eukaryotic evolution.
During
> 1 BY of Archaeplastida history, TALE TF networks have undergone
three duplication events compared to the simple dyadic TALEs in
Glaucophyta. In Viridiplantae, the KNOX class persists as a single
member throughout the mostly unicellular Chlorophyta, whereas it
duplicated into KNOX1 and KNOX2 in the multicellular Streptophyta [33].
In Rhodophyta, two KNOX classes, KNOX-Red1 and KNOX-Red2 differ in KN-A
and KN-B domains, suggesting sub-functionalization. The third
duplication event occurred in the non-KNOX group of the Chlorophyta,
whose sequences then underwent rapid divergence in their homeodomain and
heterodimerization domains, rendering their classification trickier
than other classes. Despite this divergence, proteins in one of the two
radiations (Class-B and Mam-B) were found to heterodimerize with KNOX
homologs, suggesting that these non-KNOX members serve as regulators of
KNOX/GLX heterodimers. We summarize our finding in Fig. 1, Additional file 3: Figures S5, S10.
What would have been the critical drivers of TALE heterodimerization networks emerging from ancestral homodimers?
We
found two conserved intron positions and shared sequence motifs between
the KNOX- and PBX-groups, generating our hypothesis that a proto-TALE
protein initially engaged in homodimerization and then duplicated and
diversified into two heterodimerizing classes (Fig. 1, Additional file 3: Figure S5). Heterodimerization-dependent subcellular localization [10, 34],
coupled with numerous combinations of distinct DNA-binding modules that
fine-tune target specificity, then generated customized
transcription-on switches.
During
sexual development, it is critical to accurately detect the fusion of
two cells before initiating diploid development and to make sure that
the mating combines correct partner gametes. TF heterodimerization can
implement both steps if one TF partner is contributed by each gamete. In
fact, TALE heterodimerization plays a central role as a developmental
switch for the haploid-to-diploid transition in green algae and land
plants [10, 19]. A similar haploid-to-diploid transition triggered by TF heterodimerization has recently been documented in Dictyostelium [35] and is well described in Basidiomycete fungi that utilize non-TALE homeobox proteins such as bW and bE [36, 37].
Discovery
of new prokaryotic life forms, especially in the Archaea domain,
suggests that multiple symbiotic mergers of different life forms evolved
into the proto-eukaryotes, possibly first as a symbiotic community,
which then evolved into the last eukaryotic common ancestors (LECA) that
rapidly diverged into the eukaryotic supergroups [38, 39, 40].
This eukaryogenesis model predicts that the proto-eukaryotes ➔ LECA
transition required the faithful transmission of traits between
progenitor cells and their progeny to evolve as individual lineages by
Darwinian selection. Under this hypothesis, we anticipate that the
generation of the LECA may have been driven by the sexual mechanisms
that distinguish a cellular merger between the common descendants from a
merger between unrelated community members. Our proposal for the
evolution of heterodimeric TALEs from the homodimeric proto-TALE may
provide one of the necessary mechanisms for the first sexual mode of
reproduction that might have driven the generation of the LECA from its
proto-eukaryotic ancestors.
Does expansion of heterodimerizing TALE TFs relate to the emergence of multicellular complexity?
Plant
studies have shown that the duplicated KNOX classes serve distinct
functions: the plant KNOX1 class regulates the differentiation of an
undifferentiated cell mass into spores in mosses or leafy organs in
vascular plants, and the plant KNOX2 class regulates the transition from
haploid gametophytes to diploid sporophytes in mosses and controls
secondary cell wall development in vascular plants [18, 41, 42, 43].
On the other hand, we know very little about the function of the class B
TALE in Viridiplantae and how the heterodimerization network of TALE
proteins was restructured following the KNOX1/KNOX2 duplication in
Streptophyta. Based on the diversified functions of the KNOX1/KNOX2, we
propose that the duplicated TALE heterodimers in the Streptophyta
allowed independent regulation of cellular differentiation and life
cycle transitions, priming the emergence of land plants by expanding the
diploid phase of their life cycle from a dormant zygospore to a
multicellular individual bearing many meiotic spores. One of the
critical events coinciding the emergence of land plants was the
disappearance of the diverse non-KNOX classes except the BELL class,
which might have allowed the transition to the multicellular diploid
phase by averting the sporogenesis in the zygote. During land plant
evolution, the repertoire of TALE heterodimers continued to expand,
serving all the major organ differentiation programs in the diploid
phase of their life cycle.
Can
a similar expansion of TALE heterodimers be found during Metazoan
evolution? Our search for TALE TFs in unicellular relatives of the
Metazoa—Salpingoeca and Monosiga—revealed a simple configuration with
one MEIS- and one PBC-like TALE (Additional file 3: Figures S11, S12), whereas at the Metazoan base one finds at least three MEIS-related classes and two PBC-related classes [44].
These findings suggest the occurrence of a similar expansion of a
founding dyad during Metazoan evolution. Therefore, in both plants and
animals, the TALE TF network seems to be redeployed for complex
multicellularity, departing from its posited original function in sexual
development.
Our
results suggest that TALE TF networks represent early-evolving
developmental mechanisms. That said, the emergence of complex
multicellularity doubtless required more than TF networks. TF-based
developmental cues need to be propagated via chromatin-level regulatory
mechanisms that establish the cellular memory during embryo development.
The extent to which chromatin-level regulatory mechanisms are involved
in the development of unicellular organisms is a critical question in
elucidating the origins of complex multicellularity.
Our
study explored a deeper evolutionary history of heterodimerizing TALE
transcription factors and identified true homology among the protein
domains that mediate the TALE heterodimerization of the animal MEIS/PBC
dyads and the algal KNOX/GSP1 dyads. We showed that the homology extends
to Excavata lineage close to the last eukaryotic common ancestors.
Collectively, our findings place the origin of TALE-TALE
heterodimerization near the eukaryotic root. Considering profoundly
conserved sexual role of the TALE heterodimer in Viridiplantae [10, 18, 19],
we hypothesize that the TALE heterodimeric configuration evolved to
provide a means to ensure whether the cell fusion is correctly executed
between appropriate partner gametes as a prerequisite for the evolution
of eukaryotic sexuality.
Axenic Micromonas commoda (RCC299) and Ostreococcus tauri (OTH95) were maintained in Keller medium [45] in artificial seawater at room temperature. One hundred mL of a 14-day-old culture was harvested for genomic DNA extraction. Picocystis salinarum (CCMP1897) was obtained from the National Center for Marine Algae and Microbiota (NCMA), maintained in L1 medium [46] in artificial sea water, and plated on 1.5% Bactoagar-containing media for single-colony isolation. Genomic DNA of P. salinarum was then obtained from a culture derived from one colony.
Phylogenetic analysis and classification of homeobox genes
Archaeplastida algal TALE homeodomains were collected from the available genomes and transcriptomes listed in Additional file 1: Table S1. Details of how TALE sequence was collected is provided in Additional file 5:
Method S1. After excluding nearly identical sequences, a total of 96
sequences together with 18 reference TALE sequences were made into the
final homeodomain alignment with 70 unambiguously aligned positions with
eight gapped and one constant sites. Details of phylogenetic
reconstruction is provided in Additional file 5: Method S2.
Bioinformatics analysis
The
entire TALE collection was divided into multiple groups representing
major clades in the homeodomain tree. Each group was individually
analyzed by running MEME4.12 in the motif-discovery mode with default
option collecting up to 10 motifs at http://meme-suite.org/ [47].
The search provided multiple non-overlapping motifs, many of which were
combined according to previously identified domains such as bipartite
KN-A/KN-B, ELK, and HD [14] and independent domain searches against the INTERPRO database (http://www.ebi.ac.uk/interpro/) [48].
All the collected TALE-associated homology domains were aligned to
generate HMM motifs by HMMbuild (v3.1b2), which we used to test if these
homology domains are specific to the TALE sequences using HMMsearch
(v3.1b2) against the genome-wide protein collections with E-value of
0.01 as the per-domain inclusion threshold [49].
All the homology domain information was used to locate any error in
gene predictions, and gene models were updated if necessary (Details of
the gene model curation is provided in Additional file 5: Method S3).
Intron comparison
Introns
within the homeodomain were collected and labeled as site numbers of
the homeodomain (1–60 plus 'abc' for the three amino acid extension
shared by the TALE homeodomain). If an intron is between two codons it
is denoted N/N + 1, where N is the last amino acid site number of the
preceding exon; introns within a codon are denoted N(n/n + 1), where n
is one or two for the codon nucleotide position relative to the
splice-sites.
Yeast-two-hybrid analysis
M. commoda (affixed with Micco), O. tauri (affixed with Ostta), and P. salinarum (affixed with Picsa) TALE protein coding sequences were cloned by PCR using primers designed herein (Additional file 1)
from genomic DNAs prepared by the phenol/chloroform extraction and
ethanol precipitation method. Micco_62153 and Picsa_04684 contained a
single intron, whereas all the other nine genes lacked an intron in the
entire open reading frame. For cloning of Micco_62153, we synthesized
the middle fragment lacking the intron and ligated them via XhoI and ClaI sites. For cloning details, see Additional file 5: Method S4.
We thank Mary Berbee and her laboratory members for the valuable comments on the manuscript.
Funding
This work was supported by
Discovery Grant 418471-12 from the Natural Sciences and Engineering
Research Council (NSERC) (to J.-H.L.), by the Korea CCS R&D Center
(KCRC), Korean Ministry of Science, grant no. 2016M1A8A1925345 (to
J.-H.L.), NSF-IOS0843119 (to A.Z.W.), GBMF 3788 (to A.Z.W.), and NSF
CAREER-1453639 (to E.K.). Postdoctoral support of S.J. was from
NSF-IOS0843506 (awarded to Ursula W.Goodenough).
Availability of data and materials
All data generated or analyzed during this study are included in this published article and its Additional files 1-5.
Authors’ contributions
SJ,
AZW, and J-HL designed the study. SJ, MHW, GL, JL, EK, SS, and J-HL
collected and prepared biological materials. SJ, MHW, GL, JL, EK, SS,
and J-HL organized sequence data collection. SJ, MHW, and J-HL analyzed
data. SJ, MHW, and J-HL drafted the manuscript. SJ, EK, AZW, and J-HL
edited the manuscript. All authors read and approved the final
manuscript.
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open AccessThis article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/),
which permits unrestricted use, distribution, and reproduction in any
medium, provided you give appropriate credit to the original author(s)
and the source, provide a link to the Creative Commons license, and
indicate if changes were made. The Creative Commons Public Domain
Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
Billeter
M, Qian YQ, Otting G, Muller M, Gehring W, Wuthrich K. Determination of
the nuclear magnetic resonance solution structure of an Antennapedia
homeodomain-DNA complex. J Mol Biol. 1993;234:1084–93.View ArticleGoogle Scholar
Azpiazu N, Morata G. Functional and regulatory interactions between Hox and extradenticle genes. Genes Dev. 1998;12:261–73.View ArticleGoogle Scholar
Hudry
B, Thomas-Chollier M, Volovik Y, Duffraisse M, Dard A, Frank D, et al.
Molecular insights into the origin of the Hox-TALE patterning system.
elife. 2014;3:e01939.View ArticleGoogle Scholar
Hake
S, Smith HMS, Holtan H, Magnani E, Mele G, Ramirez J. The role of KNOX
genes in plant development. Annu Rev Cell Dev Biol. 2004;20:125–51.View ArticleGoogle Scholar
Hay A, Tsiantis M. KNOX genes: versatile regulators of plant development and diversity. Development. 2010;137:3153–65.View ArticleGoogle Scholar
Berthelsen
J, Kilstrup-Nielsen C, Blasi F, Mavilio F, Zappavigna V. The sub
cellular localization of PBX1 and EXD proteins depends on nuclear import
and export signals and is modulated by association with PREP1 and HTH.
Genes Dev. 1999;13:946–53.View ArticleGoogle Scholar
Stevens
KE, Mann RS. A balance between two nuclear localization sequences and a
nuclear export sequence governs extradenticle subcellular localization.
Genetics. 2007;175:1625–36.View ArticleGoogle Scholar
Bhatt
AM, Etchells JP, Canales C, Lagodienko A, Dickinson H. VAAMANA--a
BEL1-like homeodomain protein, interacts with KNOX proteins BP and STM
and regulates inflorescence stem growth in Arabidopsis. Gene.
2004;328:103–11.View ArticleGoogle Scholar
Smith
HMS, Hake S. The interaction of two homeobox genes, BREVIPEDICELLUS and
PENNYWISE, regulates internode patterning in the Arabidopsis
inflorescence. Plant Cell. 2003;15:1717–27.View ArticleGoogle Scholar
Lee
J-H, Lin H, Joo S, Goodenough U. Early sexual origins of homeoprotein
heterodimerization and evolution of the plant KNOX/BELL family. Cell.
2008;133:829–40.View ArticleGoogle Scholar
Noyes
MB, Christensen RG, Wakabayashi A, Stormo GD, Brodsky MH, Wolfe SA.
Analysis of homeodomain specificities allows the family-wide prediction
of preferred recognition sites. Cell. 2008;133:1277–89.View ArticleGoogle Scholar
Bürglin
TR. Analysis of TALE superclass homeobox genes (MEIS, PBC, KNOX,
Iroquois, TGIF) reveals a novel domain conserved between plants and
animals. Nucleic Acids Res. 1997;25:4173–80.View ArticleGoogle Scholar
Bellaoui
M, Pidkowich MS, Samach A, Kushalappa K, Kohalmi SE, Modrusan Z, et al.
The Arabidopsis BELL1 and KNOX TALE homeodomain proteins interact
through a domain conserved between plants and animals. Plant Cell.
2001;13:2455–70.View ArticleGoogle Scholar
Bürglin TR. The PBC domain contains a MEINOX domain: coevolution of Hox and TALE homeobox genes? Dev Genes Evol. 1998;208:113–6.View ArticleGoogle Scholar
Mukherjee
K, Brocchieri L, Bürglin TR. A comprehensive classification and
evolutionary analysis of plant homeobox genes. Mol Biol Evol.
2009;26:2775–94.View ArticleGoogle Scholar
Nishimura
Y, Shikanai T, Nakamura S, Kawai-Yamada M, Uchimiya H. Gsp1 triggers
the sexual developmental program including inheritance of chloroplast
DNA and mitochondrial DNA in Chlamydomonas reinhardtii. Plant Cell.
2012;24:2401–14.View ArticleGoogle Scholar
Joo
S, Nishimura Y, Cronmiller E, Hong RH, Kariyawasam T, Wang MH, et al.
Gene regulatory networks for the haploid-to-diploid transition of
Chlamydomonas reinhardtii. Plant Physiol. 2017;175:314–32.View ArticleGoogle Scholar
Sakakibara
K, Ando S, Yip HK, Tamada Y, Hiwatashi Y, Murata T, et al. KNOX2 genes
regulate the haploid-to-diploid morphological transition in land plants.
Science. 2013;339:1067–70.View ArticleGoogle Scholar
Horst
NA, Katz A, Pereman I, Decker EL, Ohad N, Reski R. A single homeobox
gene triggers phase transition, embryogenesis and asexual reproduction.
Nat Plants. 2016;2:15209.View ArticleGoogle Scholar
Adl
SM, Simpson AGB, Lane CE, Lukeš J, Bass D, Bowser SS, et al. The
revised classification of eukaryotes. J Eukaryot Microbiol.
2012;59:429–93.View ArticleGoogle Scholar
Worden
AZ, Follows MJ, Giovannoni SJ, Wilken S, Zimmerman AE, Keeling PJ.
Rethinking the marine carbon cycle: factoring in the multifarious
lifestyles of microbes. Science. 2015;347:1257594.View ArticleGoogle Scholar
Guillou
L, Eikrem W, Chrétiennot-Dinet MJ, Le Gall F, Massana R, Romari K,
Pedrós-Alió C, Vaulot D. Diversity of Picoplanktonic Prasinophytes
assessed by direct nuclear SSU rDNA sequencing of environmental samples
and novel isolates retrieved from Oceanic and coastal marine ecosystems.
Protist. 2004;155:193–214.View ArticleGoogle Scholar
Lewis LA, McCourt RM. Green algae and the origin of land plants. Am J Bot. 2004;91:1535–56.View ArticleGoogle Scholar
Yoon
HS, Muller KM, Sheath RG, Ott FD, Bhattacharya D. Defining the major
lineages of red algae (RHODOPHYTA). J Phycol. 2006;42:482–92.View ArticleGoogle Scholar
Jackson C, Clayden S, Reyes-Prieto A. The Glaucophyta: the blue-green plants in a nutshell. Acta Soc Bot Pol. 2015;84:149–65.View ArticleGoogle Scholar
Worden
AZ, Lee JH, Mock T, Rouze P, Simmons MP, Aerts AL, et al. Green
evolution and dynamic adaptations revealed by genomes of the marine
Picoeukaryotes Micromonas. Science. 2009;324:268–72.View ArticleGoogle Scholar
Archibald JM. Genomic perspectives on the birth and spread of plastids. Proc Natl Acad Sci. 2015;112:10147–53.View ArticleGoogle Scholar
Bertolino
E, Reimund B, Wildt-Perinic D, Clerc RG. A novel homeobox protein which
recognizes a TGT core and functionally interferes with a
retinoid-responsive motif. J Biol Chem. 1995;270:31178–88.View ArticleGoogle Scholar
Kurvari
V, Grishin NV, Snell WJ. A gamete-specific, sex-limited homeodomain
protein in Chlamydomonas. J Cell Biol. 1998;143:1971–80.View ArticleGoogle Scholar
Piganeau
G, Grimsley N, Moreau H. Genome diversity in the smallest marine
photosynthetic eukaryotes. Res Microbiol. 2011;162:570–7.View ArticleGoogle Scholar
Clarke
M, Lohan AJ, Liu B, Lagkouvardos I, Roy S, Zafar N, et al. Genome of
Acanthamoeba castellanii highlights extensive lateral gene transfer and
early evolution of tyrosine kinase signaling. Genome Biol. 2013;14:R11.View ArticleGoogle Scholar
Frangedakis
E, Saint-Marcoux D, Moody LA, Rabbinowitsch E, Langdale JA.
Nonreciprocal complementation of KNOX gene function in land plants. New
Phytol. 2016;341:95–604.Google Scholar
Longobardi
E, Penkov D, Mateos D, De Florian G, Torres M, Blasi F. Biochemistry of
the tale transcription factors PREP, MEIS, and PBX in vertebrates.
Wellik D, Torres M, Ros M, editors Dev Dyn 2014;243: 59–75.Google Scholar
Hedgethorne
K, Eustermann S, Yang J-C, Ogden TEH, Neuhaus D, Bloomfield G.
Homeodomain-like DNA binding proteins control the haploid-to-diploid
transition in Dictyostelium. Sci Adv. 2017;3:e1602937.Google Scholar
Kües
U, Asante-Owusu RN, Mutasa ES, Tymon AM, Pardo EH, O'Shea SF, et al.
Two classes of homeodomain proteins specify the multiple a mating types
of the mushroom Coprinus cinereus. Plant Cell. 1994;6:1467–75.View ArticleGoogle Scholar
Spit
A, Hyland RH, Mellor EJC, Casselton LA. A role for heterodimerization
in nuclear localization of a homeodomain protein. Proc Natl Acad Sci U S
A. 1998;95:6228–33.View ArticleGoogle Scholar
O’Malley MA. Endosymbiosis and its implications for evolutionary theory. Proc Natl Acad Sci U S A. 2015;112:10270–7.View ArticleGoogle Scholar
López-García P, Eme L, Moreira D. Symbiosis in eukaryotic evolution. J Theor Biol. 2017;434:20–33.View ArticleGoogle Scholar
Zaremba-Niedzwiedzka
K, Caceres EF, Saw JH, Bäckström D, Juzokaite L, Vancaester E, et al.
Asgard archaea illuminate the origin of eukaryotic cellular complexity.
Nature. 2017;541:353–8.View ArticleGoogle Scholar
Barton MK, Poethig RS. Formation of the shoot apical meristem in Arabidopsis thaliana: an analysis of development in the wild type and in the shoot meristemless mutant. Development. 1993;119:823–31.Google Scholar
Li
E, Bhargava A, Qiang W, Friedmann MC, Forneris N, Savidge RA, et al.
The class II KNOX gene KNAT7 negatively regulates secondary wall
formation in Arabidopsis and is functionally conserved in Populus. New
Phytol. 2012;194:102–15.View ArticleGoogle Scholar
Furumizu
C, Alvarez JP, Sakakibara K, Bowman JL. Antagonistic roles for KNOX1
and KNOX2 genes in patterning the land plant body plan following an
ancient gene duplication. PLoS Genet. 2015;11:e1004980.View ArticleGoogle Scholar
Mukherjee
K, Bürglin TR. Comprehensive analysis of animal TALE homeobox genes:
new conserved motifs and cases of accelerated evolution. J Mol Evol.
2007;65:137–53.View ArticleGoogle Scholar
Keller MD, Selvin RC, Claus W, Guillard RRL. Media for the culture of oceanic ultraphytoplankton1,2. J Phycol. 2007;23:633–8.View ArticleGoogle Scholar
Guillard RRL, Hargraves PE. Stichochrysis immobilis is a diatom, not a chrysophyte. Phycologia. 1993;32:234–6.View ArticleGoogle Scholar
Bailey TL, Johnson J, Grant CE, Noble WS. The MEME suite. Nucleic Acids Res. 2015;43:W39–49.View ArticleGoogle Scholar
Finn
RD, Attwood TK, Babbitt PC, Bateman A, Bork P, Bridge AJ, et al.
InterPro in 2017-beyond protein family and domain annotations. Nucleic
Acids Res. 2017;45:D190–9.View ArticleGoogle Scholar
HMMER: biosequence analysis using profile hidden Markov models. 2017. http://www.hmmer.org. Accessed 3 June 2017.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.




Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.