Over
the last 20 years, compression fossils of feathers surrounding
dinosaurs have greatly expanded our understanding of the origin and
evolution of feathers. One of the most peculiar feather morphotypes
discovered to date are rachis dominated feathers (RDFs), which have also
been referred to as proximally ribbon-like pennaceous feathers (PRPFs).
These elongate feathers are only found in the tail plumage, typically
occurring in pairs with both streamer (not proximally ribbon-like) and
racket-plume morphologies recognized. Here we describe a large sample
set of isolated and paired RDFs from Upper Cretaceous Burmese amber
(~ 99 Ma). Amber preserves the finest details of these fragile
structures in three dimensions, demonstrating that RDFs form a distinct
feather morphotype with a ventrally open rachis, and with significant
variability in pigmentation, microstructure, and symmetry.
The discovery of numerous feathered dinosaur specimens in China (e.g., Ji and Ji 1996; Ji et al. 1998; Chen et al. 1998; Xu et al. 2014) and other regions (e.g., van der Reest et al. 2016)
has greatly improved knowledge of early dinosaur feathers and their
evolution, displaying a wide range of feather types (Zhang et al. 2006).
Some appear comparable to those that exist in modern birds while others
are more unusual. One of the most curious morphotypes reported to date
is the rachis dominated feather (RDF), which has been documented in two
lineages of basal birds, the Confuciusornithiformes and Enantiornithes
(O’Connor et al. 2012),
but appears absent in living birds. Similar feathers may also be
present in the Scansoriopterygidae, judging from the traces of tail
feathers preserved in the holotype of Epidexipteryx hui (Zhang F.C. et al., 2008).
Although early interpretations of these feathers were confused by incompleteness and poor preservation (Zhang and Zhou 2000),
recent discoveries of more complete RDF compression fossils suggested
that this unique, extinct morphotype may have been derived from a
typical pennaceous feather (O’Connor et al. 2012; Wang et al. 2014).
The earliest known traces consisted of poorly preserved rachises, which
appear ‘ribbon-like’, and the first complete feathers described were
racket-plumes (pennaceous vanes distally with ‘ribbon-like’ base). The
term PRPF (proximally ribbon-like pennaceous feather) led to at least
one distinct morphology being lumped together with RDFs due to similarly
‘ribbon-like’ proximal portions (Xu et al. 2010).
However, recent discoveries of fully pennaceous RDFs suggest that this
morphotype may be a specialized form of normal pennaceous feather. This
hypothesis is further supported by the discovery of a medial stripe in
the unmodified remiges and rectrices of ornithothoracines (Wang et al. 2014, 2015)—a
feature that was previously thought to be restricted to elongate RDFs
in tail plumage. Furthermore, morphological differences in
enantiornithines and confuciusornithiforms suggest that a pair of
elongate racket-plume tail RDFs evolved more than once (Wang et al. 2014).
Understanding
of RDF morphology has been strongly hindered by their preservation as
compressed carbon films in sedimentary rocks: limited morphological
details can be observed in this setting. A lack of topography has led to
difficulties in establishing three-dimensional structure and areas of
overlap, and the finest morphological features are not consistently
preserved. The medial stripe is an excellent example of this problem.
The line that runs down the middle of the rachis in RDFs has been viewed
as a groove (Wang et al. 2014),
supported by the discovery of slightly three-dimensional impressions of
RDFs alongside a juvenile enantiornithine from Brazil (Carvalho et al. 2015), like the depression seen on the ventral surface of modern flight feathers (Lucas and Stettenheim 1972).
The medial stripe has also been interpreted as a thickened ridge of
keratin within a hollow, tubular rachis (e.g., Zheng et al. 2009), or a complete rachis equivalent (e.g., Zhang Z. et al., 2008; Zhang F.C. et al., 2008). Isolated Cretaceous feathers in amber from Canada, France, Japan, Lebanon, Myanmar, Spain and the USA (Schlee and Glöckner 1978; Grimaldi and Case 1995; Grimaldi et al. 2002; Delclòs et al. 2007; Perrichot et al. 2008; McKellar et al. 2011; Peñalver et al. 2017), and skeletally associated feathers in amber (Xing et al. 2016a, b, 2017)
have recently become a valuable supplement to the compression fossil
record of feathers. Amber preserves three-dimensional specimens with
microstructure and pigmentation (Thomas et al. 2014)
and has the potential to clarify problematic structures and reveal fine
morphological details unlikely to preserve in compression fossils.
Here
we describe 31 Burmese amber samples that contain feathers, which
elucidate the RDF morphology and its range of variability. Twenty-five
specimens contain diagnostic RDFs (including ten pairs of feathers), and
eight fragmentary feathers are less conclusive. The feathers reveal an
open rachis with no pith cavity like that uniformly present in
previously described feathers entombed in amber, differing significantly
from previous hypotheses regarding RDF structure based on compression
fossils.
The
material originates from the Cretaceous amber deposit of Noije Bum, in
the Kachin state of Myanmar, which is the main source of amber in the
country. Radiometric dating performed with zircons in the matrix
associated with amber has provided an estimated age of the deposit at
98.8 (±0.62) Ma (Shi et al. 2012).
This amber is frequently referred to as “Burmese amber” and has been
mined for centuries. It is the most productive source of Cretaceous
amber inclusions and is considered as one of the most important deposits
for the study of Cretaceous terrestrial life (Grimaldi et al. 2002; Ross et al. 2010).
More recently, its exceptional preservation potential for vertebrate
inclusions was highlighted by the discovery of remarkable inclusions,
such as primitive birds (Xing et al. 2016a, 2017) and a coelurosaur tail (Xing et al. 2016b).
The chemical composition and botanical inclusions found in this amber
suggest that the botanical source of the resin is among the
Araucariaceae or the Dipterocarpaceae (Ross et al. 2010).
Observations
were performed using a stereomicroscope (Leica MZ12.5) and a compound
microscope (Olympus CH30). Macrophotographs were prepared at the Royal
Saskatchewan Museum (Regina, Saskatchewan, Canada) using a Visionary
Digital photography station consisting of a Canon EOS 5D DSLR camera
equipped with Canon MP-E 65 mm or EF-50 mm lenses. The pictures obtained
at various focal lengths were subsequently combined using Helicon Focus
5.3.14 software, to achieve an increased depth of field.
Microphotographs were taken with a Sony NEX-5 camera attached to the
Olympus CH30 compound microscope, as well as with a Canon EOS Rebel T6i
attached to the Leica MZ12.5. Because of the amber thickness, some of
the samples were placed in a glycerin bath to improve their light
transmission and remove optical distortions. Flow lines within the amber
were observed in greater detail using UV light (395 nm).
SEM
observations were made at the University of Alberta, Department of
Earth and Atmospheric Science (Edmonton, Alberta, Canada), using a Zeiss
Sigma 300 VP-FESEM operated in variable pressure mode with uncoated
samples. These observations were supplemented with backscatter and
energy dispersive X-ray spectroscopy to examine the composition and
preservation of the feathers.
The
institutional abbreviation “DIP” stands for: Dexu Institute of
Palaeontology, Chaozhou, China; “MCAC” stands for Mingchuan Amber
Center, Beijing, China; “RSKM” stands for Royal Saskatchewan Museum,
Regina, Canada. Feather terminology follows that of Lucas and
Stettenheim (1972), and Prum (1999)
(specifically the anatomical orientations for the feather and follicle:
the posterior margin of the follicle corresponds to the ventral surface
of the feather in the samples analyzed); while pigmentation and feather
microstructure largely follow the terminology of Dove (2000).
It
is possible to make some generalizations about the observed structures
and their variability within the amber specimen set. These general
observations are followed by a more detailed description of each sample
(in Section 3.2).
3.1.1 Rachis morphology
Compression
fossils of RDFs have led to the interpretation that these feathers
represent modified pennaceous feathers comparable to modern bird
feathers but with an enlarged rachis (thus rachis-dominated) (O’Connor
et al. 2012),
implying a broad, hollow rachis with a crescentic cross-section.
However, preservation of a wide range of RDF morphologies in amber
(Fig. 1a–c)
reveals that unlike modern feathers or previous hypotheses based on
fossils, RDFs have an exposed rachidial ridge surrounded by two laminae
of fused and unfused barb ridges, forming a C-shaped section opening
ventrally (Figs. 1 and 2).
Put in simpler terms, the rachis is open on the underside of the
feather, and it lacks a pith cavity filled with spongy tissue.
These
general features are consistent among all amber samples analyzed, but
variability exists in the finer details of the rachises. The paper-thin
fused barbs, together with the rachial ridge, form the medially striped
rachis preserved in compression fossils (Wang et al. 2014; Carvalho et al. 2015).
In some specimens, the rachidial ridge is present as a faint or
sporadic line, while others have a pronounced, blade-like ridge running
the entire length of the rachis (e.g., Fig. 1e–g).
Despite this variability, all RDFs can be clearly distinguished from
normal flight feathers with a complete, pith-filled rachis, because the
latter samples have milky amber or mineral infills within their rachises
(e.g., Xing et al. 2016a: Fig. 2a–d), that are apparent in both cross-sectional and external views.
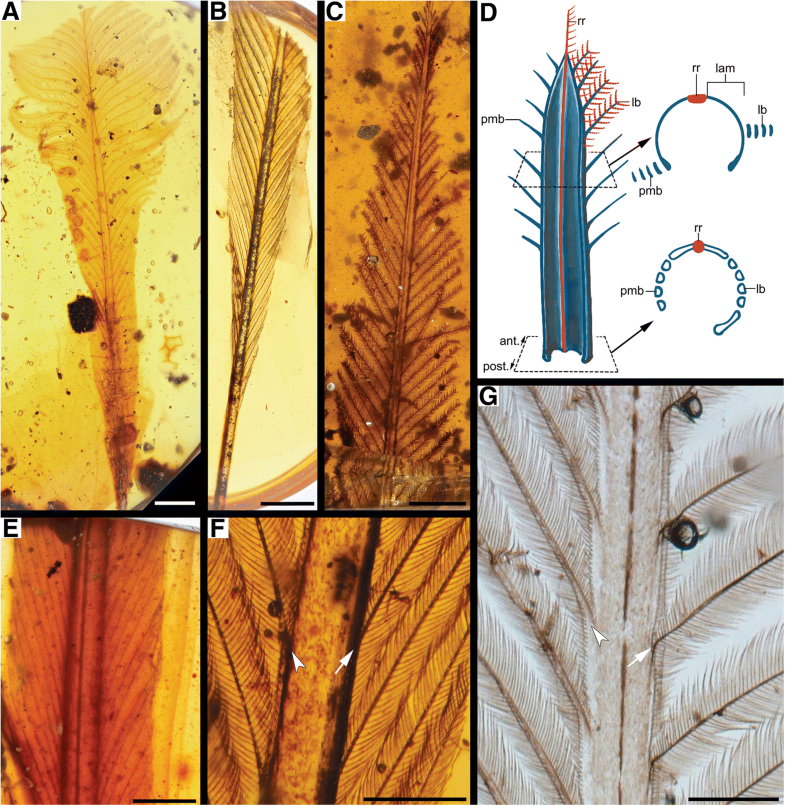
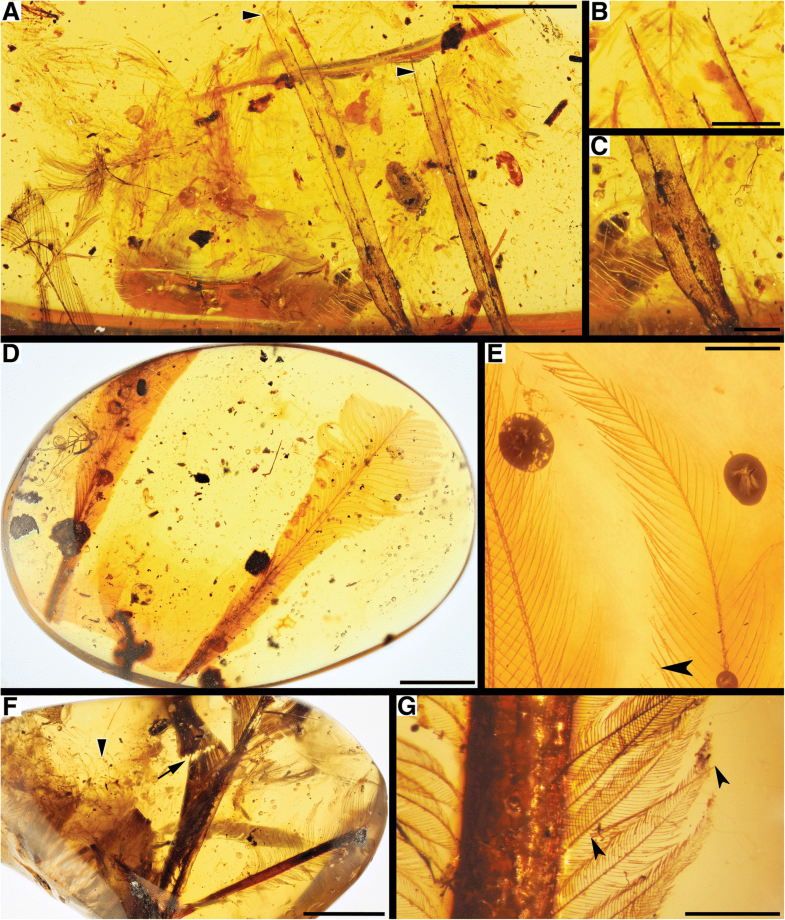
Fig. 1
Morphological diversity and rachis structure. RDFs exhibit at least three general outlines: a with barbs that appear to extend all the way down rachis, one of the two feathers in DIP-V-16186; b with naked rachis basally, DIP-V-16208 overview; c
with more pointed apices and narrower vanes, one of the two feathers in
DIP-V-16186. The general structure of all RDFs shares some common
features: d diagram of RDF
indicating rachidial ridge (dark red), and fused and unfused barb ridges
making up rachis (blue), with barbule distribution indicated only on
right side of feather apex (pale red), and with cross-sections showing
branching pattern of barbs within vaned section of RDF (upper section),
and tissue generation at follicle (lower level) required to produce
barbs at posterior margin, or along lateral margin of rachis (right side
vs. left side of each section, respectively). Rachidial ridge variants
include: e prominent ridge throughout length, DIP-V-17138; f faint, DIP-SY-06231; g or even sporadic, MCAC-0322. Arrowheads in f and g
indicate barbs attached to posterior margin of rachis, while arrows
indicate those attached to lateral surface. Abbreviations: ant –
anterior, lam – lamina, lb. – lateral barb, pmb – posterior margin barb,
post – posterior, rr – rachidial ridge. Scale bars = 2 mm in (a, b, c, e and f); 0.5 mm in (g)
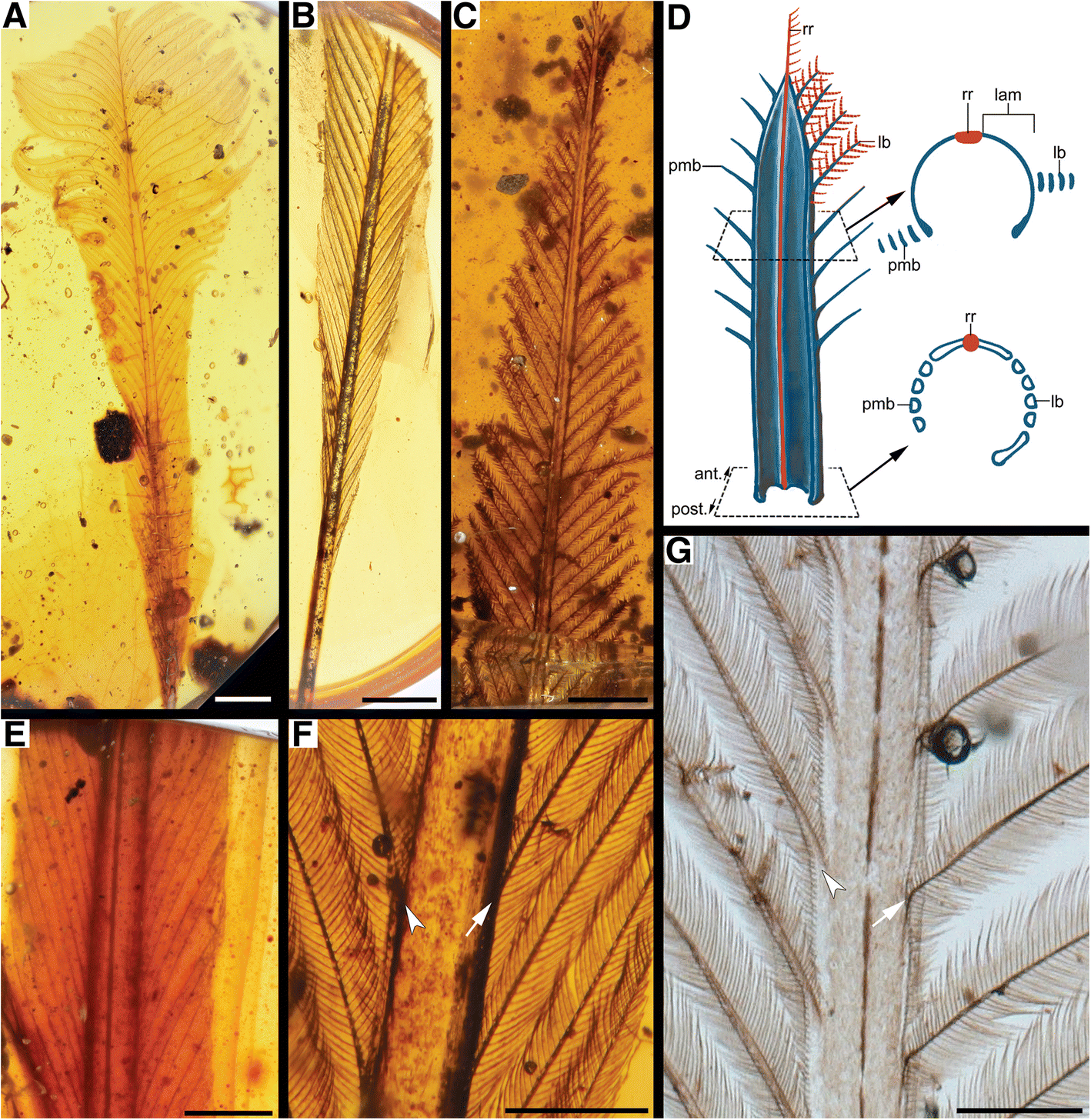
Fig. 2
RDF variations in rachis apices and cross-sectional profiles. a close-up of one of the two feather apices in DIP-V-16186, with tapered rachis leading to just rachidial ridge (arrowhead); b apex of DIP-V-17109 with broad rachis extending to the end of the vanes (arrowhead); c close-up of the tip of DIP-V-15125, with twisted rachis (arrowhead); d
oblique dorsal cross-sectional view of rachis in DIP-V-16202 exhibiting
an expansion on the edges (left and right arrowheads) and thin
rachidial ridge (center arrowhead), with barbs attaching to rachis
laterally (arrows); e cross-section of the rachis of DIP-V-16208; f SEM image of DIP-V-16207, corresponding to e; g DIP-V-17109 oblique ventral view of rachis cross-section in one of the two feathers; h
DIP-V-17127 rachis cross-section in oblique lateral view, with rami
originating slightly below (left) and exactly on the edge (right), plus
thin rachidial ridge. Arrows/arrowheads denote same structures in (d–h). Scale bars = 2 mm in (a); 0.25 mm in (b, d, g and h); 0.05 mm in (c); 0.1 mm in (e and f)
Apical
morphology of the rachis varies widely within the specimen set, but in
most RDFs, the distal ends of the laminae taper progressively. Some of
the feathers taper to just the rachidial ridge apically—in these samples
the rachis is similar in width to the adjacent barb rami along its
distal length (Fig. 2a). In contrast, specimens such as DIP-V-17109 exhibit a broad, spatulate rachis tip extending beyond the vanes (Fig. 2b), as observed in compression fossils of confuciusornithiforms, and an enantiornithine (Wang et al. 2014).
This exposed rachis tip may be a product of feather wear, or it may be a
developmental product. The removal of barbs through wear produces a
naked rachis base in the racket plumes of some modern birds (Bleiweiss 1987).
However, the barbs that lead up to the exposed rachis tip in the RDFs
with this feature show a progressive reduction in length (Fig. 5a)
that is more suggestive of a developmental source. One sample displays
an unusual rachis that tapers slightly and twists apically, bearing
three or four apical barbs (Fig. 2c). These barbs gradually diminish in length and show no sign of truncation.
The
rachis structures observed in amber imply a wider range of flexibility
than would be expected based on the straight RDFs known from compression
fossils (e.g., O’Connor et al. 2012; Wang et al. 2014; Hu et al. 2015). At the high end of the flexibility spectrum are specimens like DIP-V-17153 and DIP-V-16223 (Fig. 4d,
h, i). DIP-V-17153 contains a slender feather with barbs concentrated
toward the apex; the rachis is widely opened, appearing thin and
extremely flexible throughout its length, but gaining rigidity from its
broad, C-shaped profile; the edges of its laminae have rolled inward
near the base of the feather (Fig. 4h,
i), and are nearly flat apically.
At the low end of the flexibility
spectrum, some RDFs in amber exhibit a straight and relatively rigid
rachis, with a deep, C-shaped cross-section and thickened posterior
margins along a lamina that is otherwise less than 50 μm thick (e.g.,
Fig. 2e).
It is unclear if these marginal reinforcements are achieved through
thickening of the lamina surface, or by the posterior margin of the
lamina rolling back on itself: when combined with the rachidial ridge,
this creates a series of three somewhat tubular structures that
reinforced the C-shaped section of many RDFs (Fig. 6b).
Regardless of the depth and curvature of the rachis cross-sectional
shape, all samples observed had laminae extending from the side of the
rachidial ridge that were extremely thin compared to the surroundings of
the pith cavity in modern bird feathers (e.g., McKittrick et al. 2012:
fig. 14): excluding the rachidial ridge and any thickening near the
posterior margin of the laminae, rachis thicknesses in RDFs are
generally comparable to those of the adjacent barb rami.
3.1.2 Barb attachment
Significant
variation also exists in attachments between the lateral laminae of the
rachis and the unfused barbs that make up the vanes on each side of the
rachis. Some specimens have rami originating from the posterior margins
of the laminae (e.g., Fig. 6g), while others have rami situated laterally, removed from the posterior opening in the feather (e.g., Fig. 2d).
In the latter specimens, the attachment of barbs high on the side of
the rachis leaves a section of lamina that extends below the surface of
the vanes. This attachment position varies between specimens, and it can
also vary on either side of the same rachis. Barbs in DIP-V-17127 and
MCAC-0322 display a relative asymmetry between the two vanes (Fig. 1f,
g). On one side, rami originate slightly above the laminar margin,
while on the other side, rami are located exactly on the posterior edge,
sometimes producing a serrated rachis margin. Combined with the shape
of the rachidial ridge, and depth of the cross-sectional profile,
variability in branching from the laminae likely influences mechanical
properties of the feather, particularly its rigidity.
3.1.3 Barb branching and barbules
The dorsal and ventral surface of each RDF can be inferred from the attachment pattern of the barbules to the barbs (Figs. 2d and 4b).
In the amber specimens the barb rami extend as blade-shaped structures
ventral to the barbules, and the rachis opens ventrally to the barbs
(Fig. 2d–h).
None of the RDFs express strong asymmetry like that observed in modern
rectrices. However, some show asymmetric features related to the vanes
and the barbules. Several feathers have a short portion of the basal
shaft-bearing barbs only on one side further adding to the morphological
diversity observed in these tail ornaments (Figs. 3a, 4g, h and 7e, f). DIP-V-17153 (Fig. 4h)
bears unpaired barbs along two-thirds of the length preserved below the
main vaned feather section, with a notable dissimilarity between barbs
and barbules of the two vanes.
The left vane is characterized by
blade-shaped rami bearing thin, straight barbules, while the right vane
has similar proximal barbules, that are thick, straight and rigid
(almost acicular); but the distal barbules are thinner, curved, and much
longer than the proximal barbules (Fig. 4j).
Similarly, some RDFs exhibit a difference between proximal and distal
barbules. In these feathers, barbs show a progressive decrease in the
relative length of proximal barbules, bearing only distal barbules at
the apex (Fig. 6e).
In addition to DIP-V-17127 having barbs that attach at different
heights on each side of the rachis, there is also a clear asymmetry
between barb lengths in each vane (Figs. 2h and 8g).
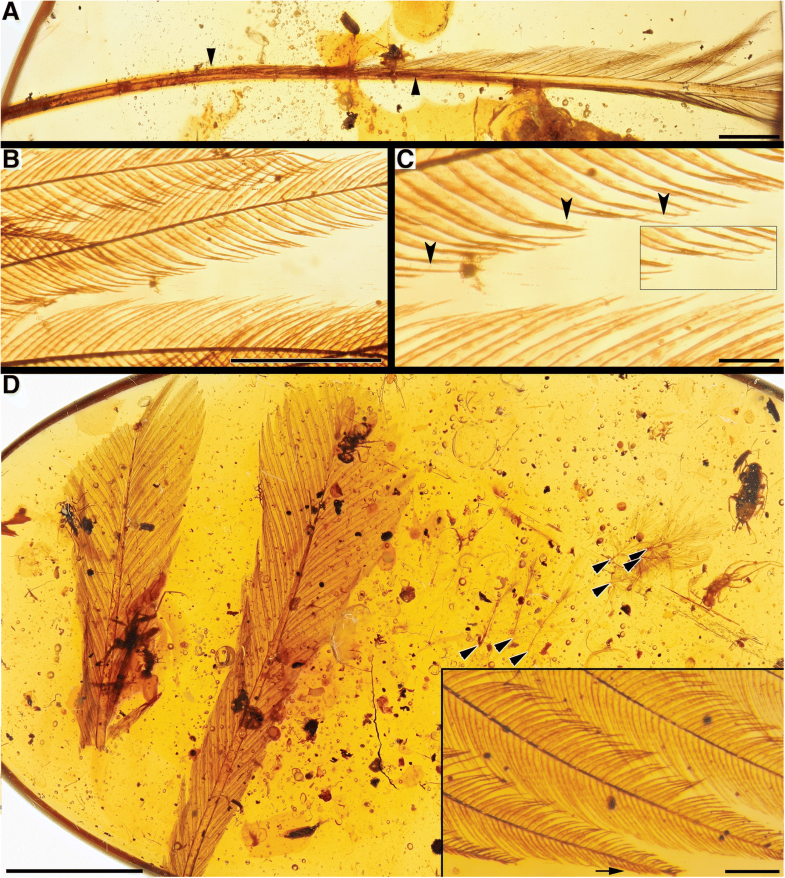
Fig. 3
RDFs with interlocking barbs. a DIP-V-16111 overview with asymmetrical vanes (arrowheads at base of each vane); b DIP-V-16111 barb structure; c DIP-V-16111 barbules with hooklets (inset and arrowheads); d
DIP-SY-06231 overview with plumulaceous feathers trapped on drying line
(arrowheads), and more detailed view of barb and barbule structure
(inset). Scale bars = 2 mm in (a); 0.5 mm in (b); 0.1 mm in (c); 5 mm in (d); 0.2 mm in (d) inset
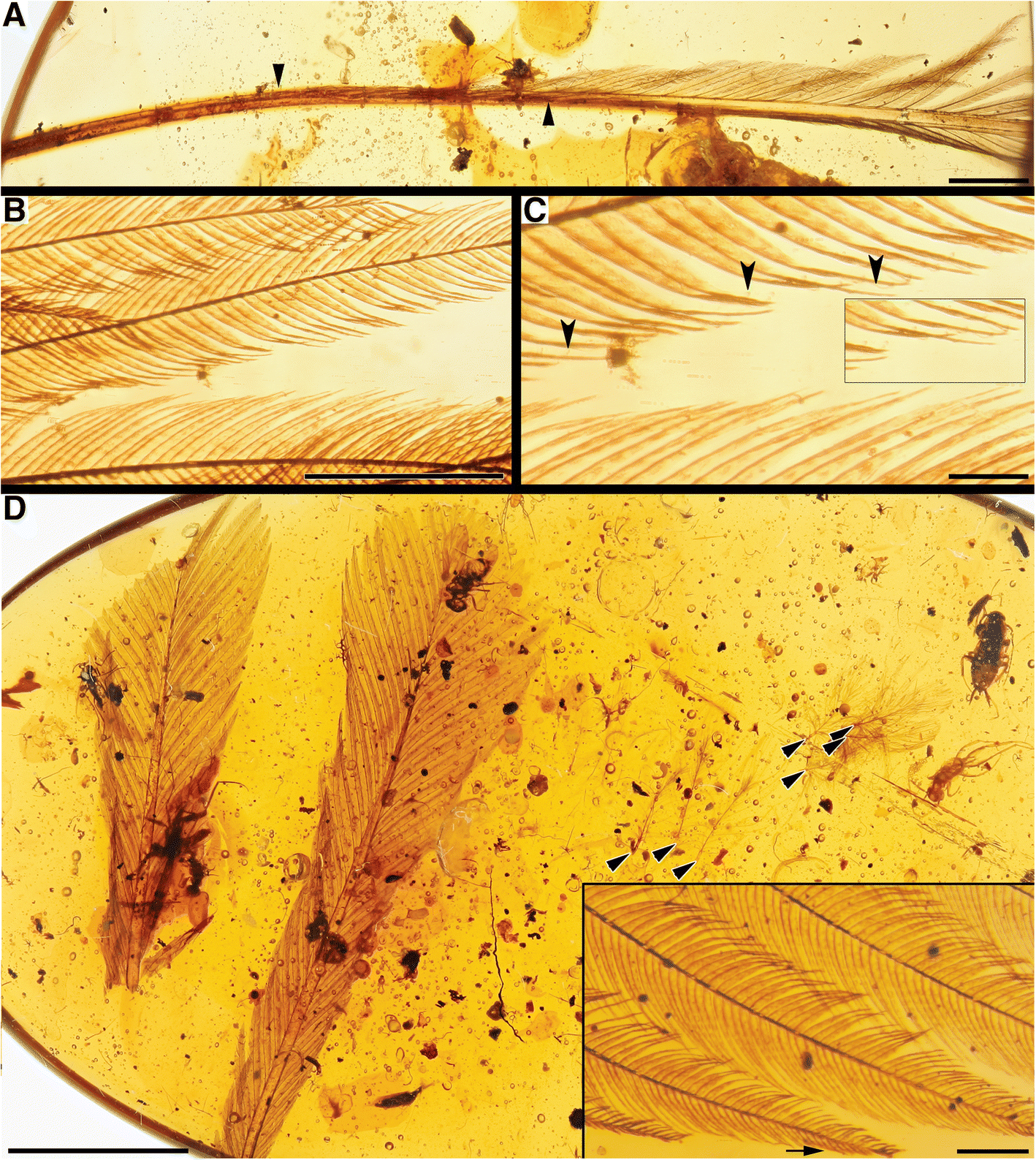
Fig. 4
RDFs with weakly interlocking barbs. a DIP-V-16202 overview; b
cross-section of the barbs of DIP-V-16202, arrowheads indicate barbule
origination points, arrow indicates barb origination on posterior margin
of rachis; c DIP-V-16202 barb, with pigment banding in barbules; d DIP-V-16223 overview, arrow indicates bending point in rachis; e detail of rachis and rachidial ridge in DIP-V-16223; f base of rachis for DIP-V-17180 (overview in g), arrow indicates breach of rachis base on polished surface of amber piece; g DIP-V-17180 overview, arrow indicates basal constriction detailed in F, arrowheads mark asymmetrical bases of feather vanes; h DIP-V-17153 overview, arrowheads mark asymmetrical bases of feather vanes, arrow indicates exposed barbs featured in j; i detail of inwardly rolled rachis base on DIP-V-17153; j cross-section of barbs of DIP-V-17153 showing elongate distal barbules. Scale bars = 2 mm in (a, d, f and i); 0.5 mm in (b); 0.2 mm in (c and j); 1 mm in (f); 5 mm in (g and h)
3.1.4 Pigmentation
The observable pigmentation in most amber specimens generally produces pale brown overall colors (e.g., Fig. 1a–c).
There is some small-scale variability in this coloration, with loosely
confined pigmentation spots corresponding to the basal cells in some
barbules (Figs. 3d and 8e). Some specimens also appear translucent and would have likely been pale or white in life (Fig. 7c, e). One remarkable specimen (Fig. 4g),
has transverse pigmentation banding on the rachis and barbs, producing
alternating dark brown and colorless bands. In general, the muted colors
preserved in amber RDFs seem to accurately represent the distribution
and intensity of pigments from melanosomes, but it is still unclear
whether these feathers would have possessed structural colors or
additional pigments such as carotenoids (Thomas et al. 2014) that would have created a greater range of colors.
3.1.5 Occurrence in pairs
In nine of the specimens studied, a pair of RDFs is preserved adjacent and subparallel to one another (Figs. 5, 6 and 7), and some have been deflected by the same resin flows (e.g., Figs. 5c and 7b).
Because of their similar taphonomy, morphology, and microstructure,
these feather pairs are each almost certainly from the same individual,
most likely representing tail streamers of basal birds. Based on
taphonomic features (detailed in Section 3.1.6),
many of these pairs appear to have been plucked on impact with resin,
as opposed to belonging to corpses. Flow lines indicate that the
streamers were dragged through the resin mass, or they are accompanied
by numerous non-RDF plumulaceous feathers stuck in a single resin drying
line.
Fig. 5
Paired RDFs. a DIP-V-17109 overview; b barbs of DIP-V-17109 with clumped barbules and blunt barb apices; c DIP-V-15153 overview, with feather apices deflected dorsally (to left) by resin flows, but rachis unaffected; d
RSKM_P3306.58 overview, horizontal arrowheads mark rachis lateral
margins, vertical arrowheads mark rachidial ridges, arrow denotes area
for detailed images in e and f; e detail of barb bases and lateral attachment to the rachis in RSKM_P3306.58; f detail of barbules in RSKM_P3306.58, arrow highlights hooklets on distal barbules. Scale bars = 5 mm in (a); 0.2 mm in (b); 2 mm in (c and d); 0.5 mm in (e and f)
Fig. 6
Paired RDFs. a DIP-V-16164 partial overview, with RDF rachises indicated by arrowheads; b oblique section through one rachis in DIP-V-16164; c distorted rachis near base of DIP-V-16164; d DIP-V-16186 overview; e barbs of DIP-V-16186, arrowhead indicating hooklets; f
DIP-V-17137 overview, arrow indicating taphonomic tear in vane,
arrowhead indicating region with numerous plumulaceous feathers; g details of rachis, barbs, and barbules in DIP-V-17137 RDF (on right side of f), with bristle-like feather floating in amber between arrowheads. Scale bars = 5 mm in (a, d and f); 1 mm in (b and c); 0.2 mm in (e); 0.5 mm in (g)
Fig. 7
Paired RDFs. a DIP-V-16105 overview; b DIP-V-17177 overview of paired, bent feathers, with arrows marking ventral deflection point; c DIP-V-17194 overview; d detail of DIP-V-17194 barbs; e MCAC-0322 overview; f detail of vane bases in MCAC-0322 (lower feather in e). Scale bars = 5 mm in (a–c and e); 2 mm in (d and f)
3.1.6 Interactions with resin flows
Sample
observations under ultraviolet light provided additional information
about the interaction between the feathers and resin flows. In some
samples the flow lines are even visible to the naked eye. In
DIP-V-16111, the resin flow lines are clearly observable and suggest
that the feather was drawn through a resin flow. Approximately at the
mid-length of the feather, the resin flow lines are distorted to form a
funnel around the rachis and the barbs (Fig. 3a).
In DIP-V-15153, the two feathers are bent apically. The flow lines
parallel the specimens, but become perpendicular apically, strongly
deflecting the barbs of the feathers (Fig. 5c).
DIP-V-17138 also exhibits a series of flow lines paralleling the
feather, which could explain the inclination of the vanes (Fig. 8f).
These observations suggest that the feathers were highly flexible, and
that some feathers were plucked when the animal hit the resin flow
rather than being molted feathers or those sourced from corpses.
Subparallel RDF alignments (Figs. 5a, d, c, 6d and 7a,
c, e) and shared deflections—combined with the presence of only RDFs,
or RDFs and a scattering of plumulaceous feathers in pieces of amber
without any additional vertebrate inclusions (Figs. 3d and 6a)—point toward impact and adhesion on drying lines in the sample set.
Fig. 8
Diagnostic RDF fragments. a DIP-V-15125 overview, arrowhead marks twist in rachis; b details of barbs and barbules in DIP-V-15125; c DIP-V-16115 overview; d DIP-V-17121 overview; e detail of DIP-V-17121 rachis and barbs, where they are cross-cut by polished surface of amber (left margin of d), arrow indicates rachidial ridge; f DIP-V-17138 overview; g DIP-V-17127 feather section; h DIP-V-16207 overview. Scale bars = 1 mm in (a); 0.5 mm in (b); 5 mm in (c, d and g); 0.2 mm in (e); 2 mm in (f and h)
3.2 Details of individual RDF specimens
From
a functional standpoint, the sample set can be generally grouped into
RDFs that exhibit evidence for interlocking barbs (forming more of an
aerodynamic surface), and those that show weakly interlocking barbs.
Some specimens are observed as pairs, while fragmentary remains can be
identified as either diagnostic or putative RDFs based on their
observable characteristics.
3.2.1 RDFs with interlocking barbs
The feather contained in DIP-V-16111 (Fig. 3a–c),
appears slender and slightly curved when observed with the naked eye.
The basal-most quarter of the preserved rachis does not bear any barbs
along its length, while the second quarter exhibits barbs only on one
side. These barbs are sparse, short and adpressed. The rest of the
rachis bears pennaceous barbs with blade-shaped rami attached to the
posterior edges of the laminae with a divergence angle of ~ 20 degrees.
Barbules stem from barbs, also from the laminae in all regions of the
feather that bear barbs. These barbules appear analogous to the
rachidial barbules that occur in contour feathers of modern birds (Lucas
and Stettenheim 1972).
In DIP-V-16111 the barbules are blade-shaped, with proximal barbules
tapering to a narrow apex, whereas distal barbules exhibit a thick
pennulum bearing hooklets (Fig. 3c).
In distal regions of the barbs, proximal barbule lengths decrease, and
they disappear at the extremity, leaving barbules on only the distal
side of the ramus. Pigmentation within the barbules is diffuse and
interrupted by barbule subdivisions (Fig. 3c).
Overall, the apparent preserved color of the feather is pale brown,
with darker rami and rachidial ridge, and pale brown laminae.
DIP-SY-06231 (Figs. 1f and 3d)
contains two types of feathers probably belonging to the same animal –
two ornamental feathers are preserved alongside numerous plumulaceous
feathers (Fig. 3d).
The RDFs differ in general morphology from DIP-V-16111 (described
above) but share some key features. Similarities include: barb rami that
are blade-shaped and branching from the posterior edges of the laminae
(average divergence angle of 23 degrees); exhibiting rachidial barbules
and asymmetry between proximal and distal barbules that is reduced
apically (Fig. 3d);
and having barbules that are similar in morphology and bear hooklets.
However, in DIP-SY-06231, the rachis is progressively tapering, with the
distal sections of the exposed rachidial ridge very similar in
morphology and size to the surrounding barb rami.
The same apical taper
is observed in DIP-V-15153 and DIP-V-16186. In DIP-SY-06231, the
rachidial ridge is barely visible as a simple brown line. Pigmented
areas within the RDFs in this sample give the specimens an overall
medium brown appearance. At a finer scale, diffuse pigmentation within
the barbules is interrupted by circular spots that appear to correspond
to the center of each barbule internode (Fig. 3d).
Pigmentation becomes more concentrated toward the apex of each barbule
and is significantly darker within the barb rami. The plumulaceous
feathers found as syninclusions in DIP-SY-06231 are preserved all the
way down to the calamus, preserving a hollow, pith-filled rachis. The
bases of the feathers are plumulaceous, while the tips of some feathers
are pennaceous. These feathers appear to lack pigmentation as preserved,
suggesting a pale or white appearance in life.
3.2.2 RDFs with weak barb interlocking
In DIP-V-16202 (Figs. 2d and 4a–c) the pronounced rachidial ridge is slightly displaced from the rachis midline (Fig. 4a).
The angle between the rami and the laminae is approximately 30 degrees,
and barbules are present between rami. Barb rami are blade-shaped,
originating laterally from the side of the laminae. Both proximal and
distal barbules are straight, narrow, and blade-like, with no visible
differentiation into a pennulum bearing hooklets in the distal barbules.
Diffuse pigmentation is preserved in the base of the barbules, and its
minor mottling hints at barbule subdivisions.
DIP-V-16208 (Figs. 1b and 2e)
shares many structural features with DIP-V-16202. However, in the
former specimen, the rachidial ridge is less pronounced, and appears to
be absent under transmitted light (Fig. 1b)
due to a thin veneer of pyrite along the inner surface of the rachis. A
rachis cross-section shows a thin rachidial ridge, as well as a tubular
thickening at the posterior margins of the laminae, slightly ventral to
the barb origination points (Fig. 2e).
Barb and barbule structure in this specimen are reduced and poorly
interlocking, but the high density of blade-like barbules combine to
produce relatively closed and complete vanes. Visible pigmentation gives
an overall medium brown coloration that is overprinted distally and in
the rachis by opaque deposits of pyrite.
The feather in DIP-V-16223 (Fig. 4d, e) has a proportionally wide (1.14 mm basally) and deep rachis (Fig. 4e).
The rachidial ridge is unusual, with a prominent blade-shape. The rami
branching points are located laterally on the laminae, and barb and
barbule morphology are similar to those found in the other weakly
interlocking RDFs. Traces of pigmentation give a pale brown apparent
coloration to the feather, overprinted by deposits of pyrite on the
rachis and distal half of the barbs. The rachis is folded ventrally 90
degrees at the base, where it has been redirected by resin flows.
Between this deflection and the base of the feather’s vanes, the lateral
margin of the rachis is torn longitudinally and curled adapically,
revealing that the lamina is thinner than a barb ramus.
DIP-V-17180 (Fig. 4f,
g) is elongate, with narrow and asymmetrical vanes apically, and an
extensive rachis without barbs basally. The feather apex is largely
preserved, but both the feather apex and base breach the amber surface:
it is unclear how much of its length is missing. Four broad color bands
extend across the feather’s length, with the bands toward the apex of
the feather forming a subtle chevron pattern (Fig. 4g).
The pigments that produce the apparent medium brown color bands are
diffuse within the rachis and barbs, and slightly concentrated within
the rachidial ridge. Barbs appear to originate from the ventral margin
of the rachis on both sides, and end in acute apices. Barbs are widely
separated, with limited potential for overlap or interlocking to form a
closed vane. The barbules are blade-shaped. The basal-most pigmented
area of the rachis has drawn inward, forming a complete ring (Fig. 4f).
There is an air bubble trapped within this region of the rachis, so it
is unclear if the constriction is related to high flexibility, partial
decay, or if the constricted region may represent the follicle insertion
point for the feather. Unfortunately, the feather is cross-cut by the
polished surface of the amber basally, making it impossible to determine
if the absolute base is preserved.
DIP-V-17153 (Fig. 4h–j) contains a slender feather with few barbs inserted on the posterior edges of the laminae (Fig. 4h).
As in DIP-V-16233, the rachis in DIP-V-17153 is widely open, but in the
latter specimen it seems to be very thin and extremely flexible—near
the base of the feather, on both sides of the rachidial ridge, the edges
of the laminae are rolled inward, toward the rachidial ridge (Fig. 4i).
The apex of the rachis is almost flat. Along the laminae, barbs are
borne only on one side across the middle one-third of the feather’s
preserved length, and the barbs and barbules exhibit a different
morphology depending on the vane observed. The left vane (in ventral
view) is formed by blade-shaped rami bearing thin, relatively straight
barbules, while the right vane has rami that are also blade-shaped but
appear much thicker. The proximal barbules are especially thick,
straight and rigid, appearing acicular, while distal barbules are thin,
curved, and several times longer than the proximal ones (Fig. 4j).
There is no evidence of the presence of hooklets on any of the visible
barbules. It is unclear if the inwardly rolled base of DIP-V-17153 has
been torn free from the remainder of the rachis, or if it might
represent the insertion point into the follicle (Fig. 4i).
The rachidial ridge becomes less deep, and the laminae appear to taper
inward near the base of the rachis, but they also present a somewhat
jagged edge basally. If this is the true base of the feather, it is not
surrounded by a calamus despite being fully encapsulated in resin. This
is unlike the bases of plumulaceous feathers that are found alongside
RDFs in specimens such as DIP-SY-06231 (Fig. 3d).
3.2.3 Paired RDFs
In DIP-V-17109 (Figs. 1c, 2b, g and 5a,
b), the rachis is partially obscured by pyrite deposits, but it is very
broad, and ends in a blunt tip that extends to the edge of the feather
vanes (Figs. 2b and 5a). This character has been reported in confuciusornithiforms as well as in an enantiornithine (O’Connor et al. 2012) compression fossils. Given their morphology, and the other findings from Burmese amber (Xing et al. 2016a, b, 2017),
these feathers probably originated from the tail of an enantiornithine.
The rachidial ridge is twice as thick as the rami within the feather
(Fig. 2g).
The rami are inserted relatively high on the sides of the laminae, far
removed from the posterior margin, and diverging with an angle of about
30 degrees. Barbules possess a large base and are locally joined to form
clusters of 3–5 barbules (Fig. 5b).
It is not entirely clear whether this clumping is part of the original
structure of the feather, or taphonomic in origin. Nonetheless, the same
clumping feature is observed in DIP-V-16113, along with similar blunt
barb apices. This points toward a structural as opposed to taphonomic
explanation. Regions of the feather in DIP-V-17109 that are not obscured
by pyrite deposition appear to lack pigmentation, suggesting a pale or
white feather in life.
DIP-V-15153 (Fig. 5c)
contains two feathers that are truncated basally. The amber surface
provides a lateral view of the feathers side by side and with the tips
deflected dorsally (towards the right in Fig. 5c)
by interactions with a resin flow. The rachis tapers distally to just
the rachidial ridge, and rami origination points are located on the very
posterior edges of the laminae. Long barbs with blunt apices contain
reduced, slightly curved barbules that continue as rachidial barbules.
Apparent pale brown coloration is generated through diffuse pigmentation
in the barbules, barb rami, and rachis.
RSKM_P3306.58 (Fig. 5d–f)
contains two closely adpressed RDFs with different rachis morphologies.
It is unclear whether these specimens represent two distinct
morphotypes, or if they are a matching pair that have simply been
sectioned at different points along their lengths. The RDF with an
exceptionally broad rachis has a barbless base that exceeds 5 mm in
width (lower specimen in Fig. 5d),
but it tapers apically to less than 1 mm in width. It also bears barbs
apically that are similar to those found on the RDF with a narrower
rachis (upper specimen in Fig. 5d).
The narrower specimen is vaned throughout its preserved length. Both
RDFs have a prominent rachidial ridge that appears slightly concave on
the dorsal surface of the rachis, and both RDFs have barbs that attach
to the posterior margin of the rachis on one side (Fig. 5e),
but the lateral surface on the other side. Barb rami are deeply
blade-shaped. Proximal barbules are blade shaped with a weak apical
taper and curvature and a divergence angle of approximately 38°, while
distal barbules have broad, blade-shaped bases that diverge at
approximately 50°, followed by a distinct pennulum that curves apically
and bears hooklets (Fig. 5f).
Preserved coloration appears to indicate a dark brown rachis and rami,
with medium-brown pigmentation in the barbules that is diffuse but
provides weak indications for seven nodes within the basal plate of each
barbule.
DIP-V-16164 (Fig. 6a–c)
preserves two adjacent and subparallel rachis bases without any barbs.
One of them exhibits a distortion basally: the laminae deflect to create
regions of the rachis that are broad and narrow, showing some
flexibility (Fig. 6c).
The amber also contains a contour feather and more than 20 plumulaceous
feathers, many of which have a similar orientation to one another but
do not attach to a sheet of skin (right side of Fig. 6a).
These feathers appear to be trapped along a different plane, but under
UV light, all feather inclusions are clearly part of the same resin
flow. This association seems to support a scenario in which an animal
struck the resin flow and left behind a mixture of predominantly down
feathers and a pair of RDFs consistent with tail feathers.
DIP-V-16186 (Figs. 1a and 6d, e) contains two feathers, one of which preserves the rachis tapering suddenly to just the rachidial ridge (Fig. 1a).
Thick rami insert slightly above the posterior edges of the laminae.
The angle between the rami and the rachis is about 20 degrees. The
barbules bear hooklets and a diffuse, weak pigmentation, suggesting a
pale original feather color, and weak attachment between the barbs (Fig.
6e).
DIP-V-17137 contains two types of feathers (Fig. 6f, g): two overlapping RDF fragments, and numerous plumulaceous feathers preserved down to the calami (Fig. 6f).
Some plumulaceous feathers exhibit a pennaceous tip (suggesting contour
feathers), but detailed observation is complicated by abundant bubbles
and milky amber in this region. The largest piece of RDF exhibits strong
taphonomic distortion: the vanes, which are composed of barbs that are
straight and interlocking, are folded and torn. The rami have a
divergence angle of approximately 15 degrees and are inserted relatively
low on the sides of the laminae. The barbules have a short and weakly
developed pennulum-bearing hooklets, and a base with a faint
pigmentation concentrated near the apex of the basal cell (Fig. 6g).
The second fragment of RDF exhibits rami that are less thick. The barbs
are folded and more open in their arrangement, with some barbs bound by
spider silk. Although this amber piece contains a pair of RDFs, and may
also be related to an impact event, the feathers are not as uniform in
morphology and subparallel in orientation as the other pairs. These
feathers may have entered the resin due to an impact, but they cannot be
as confidently interpreted as paired tail feathers. Next to the right
vane (highlighted in Fig. 6g),
is a structure that looks like a hollow spike. The morphology and the
size are compatible with the interpretation of a filament feather, such
as those observed in other enantiornithines found in Burmese amber (Xing
et al. 2017).
Some of the plumulaceous feathers found as syninclusions within this
amber piece have faint pigmentation, with nodes and internodes barely
visible.
DIP-V-16105 (Fig. 7a)
contains two adjacent feathers that underwent a strong distortion: the
rachises and the rami are taphonomically flattened. Due to this
flattening, details of the rachis are unclear, and the rachidial ridge
appears to be absent. The rami have thick bases that insert deeply and
broadly into the margins of the laminae. The barbs are highly
asymmetrical (with one vane more than four times as wide as the other).
Proximal barbules are blade-like and gently curved, while distal
barbules have a blade-like base followed by a weakly developed pennulum
that angles adapically from the base. Given the combined differences
between DIP-V-16105 and the rest of the sample set, these feathers are
less confidently placed in the RDF category.
Paired RDFs within DIP-V-17177 (Fig. 7b)
are obscured by numerous fractures within the surrounding amber, but
many of their details remain clear. The two feathers are subparallel to
one another, including a deflection of roughly 135 degrees that occurs
within the basal part of their rachises. Rachis details basal to this
deflection point are less clear, but they appear to have flared
laterally, likely because their C-shaped cross-sections were spread open
by resin flows. Within the vaned portion of each feather, the rachises
are preserved with greater translucency, while basal details are masked
by milky amber. Rachidial ridges are prominent, and barbs branch from
the lateral surfaces of each lamina. Barbs within one vane (uppermost in
Fig. 7b)
appear to branch from the rachis at a lower angle than in the other
vanes, but this may be influenced by resin flows. The apex of each vane
is not preserved. The loosely arranged barbs have blade-shaped rami and
taper gradually along their lengths. Barbules are reduced and
blade-shaped with no visible traces of hooklets or segmentation.
Pigmentation appears weak and diffuse within the barbules, and slightly
darker within the barb rami and rachis, but the thick and fractured
surrounding amber precludes study under high magnifications.
DIP-V-17194 (Fig. 7c,
d) contains the vaned apices of two subparallel RDFs partially veiled
by pyrite deposits. The basal barbs within each vane are curled
ventrally, likely from being drawn through the resin or having resin
flow around the feathers (a couple of broken barb sections between the
two feathers are also displaced ventrally). Pigmentation within the
barbs and rachis is faint or absent, likely indicating a pale or white
feather in life. The rachidial ridge is tubular and approximately 40 μm
in diameter, with a mottled appearance (Fig. 7d)—it
is not entirely clear whether this mottling is the result of
pigmentation or of spongy tissue within the rachidial ridge. The
C-shaped cross-section of the rachis is broader than it is deep, and
barbs attach to its posterior margins on both sides to form loose vanes.
The rachis does not taper strongly, extending slightly past the apical
barbs as a spatulate tip, and the rachidial ridge becomes faint
apically. The barbs have blunt tips (barbules remain long near the apex,
extending well beyond the tip of the barb ramus). Barbules have a
reduced, blade-shaped morphology, and there is little differentiation
between proximal and distal barbules. Proximal barbules diverge at a
slightly lower angle than distal barbules, but these angles are
inconsistent because all barbules are clumped into groups of two or
three, albeit less dramatically clumped than in DIP-V-17109 (Fig. 5b).
MCAC-0322 (Figs. 1g and 4e,
f) contains paired RDFs surrounded by bubbles that have developed
expansion cracks, but the surrounding amber is exceptionally clear.
Reduced pigmentation in the feathers permits detailed study of their
rachis structure (Fig. 1g).
Each feather has most of its vaned section preserved, along with part
of the barbless rachis. In general, the outer margin of each feather has
shorter barbs with more robust rami and less divergence than the inner
margin, and an asymmetric absence of barbs exists within the basal
one-third of the outer vane (Fig. 7f).
Many of the barbs within these feathers are widely spaced and cannot
connect. The rachis appears to remain broad all the way to the apex of
each feather, although the apices are not fully preserved. The rachidial
ridge is faint and sporadic. Barbs attach laterally along the inner
edge of each feather and attach to the posterior margin of the rachis
along the outer edge of each feather. This variation in branching can be
seen in multiple specimens, but is most clearly observed in the thin
strip of lamina present ventral to the barb bases, as well as the
continuation of rachidial barbules along the lamina’s lateral surface
(i.e., ‘right’ rachis margin in Fig. 1g)—this
branching pattern contrasts with the posterior margin originations that
leave a notched appearance and barbules along the posterior margin
(i.e., ‘left’ rachis margin in Fig. 1g).
MCAC-0322 displays blunt barb tips, because barbule length does not
diminish rapidly toward the apex of each barb. The barbules are
blade-shaped and simple, lacking hooklets. Weak, diffuse pigmentation is
present within the barbules, and slightly darker pigmentation is
present within the barb rami and rachidial ridge. These feathers would
likely have been pale in life, and their structure and asymmetry
strongly suggest that they are tail paired streamers, likely from an
enantiornithine.
One additional pair of RDFs was encountered: DIP-SY-06231. This specimen is described in Section 3.2.1 because of its interlocking barbs.
3.2.4 Diagnostic RDF fragments
Sample DIP-V-15125 (Figs. 2c and 8a,
b) yielded a small fragment (length: 4.1 mm) of a feather that is
partially veiled by a film of pyrite on parts of the rachis and barbs
(Fig. 8a). The rachis tapers strongly and presents an unusual twisted shape with 3 or 4 barbs attached at the extremity apically (Fig. 2c).
No rachidial ridge is visible basal to this twisted apex. The barbs
share the same blunt ends observed in sample DIP-V-17109, but the barb
terminations in DIP-V-15125 are less abrupt. The barbules display a
preserved pigmentation with spots of reduced pigment density outlining
at least five basal cells, and hooklets are present (Fig. 8b).
DIP-V-16115 (Fig. 8c)
is very rich in particulate inclusions, trichomes, organic material and
insects, suggesting an origin on the forest floor (Perrichot 2004).
One portion of a feather (lacking the tip and basal parts) was found in
this sample, and it has a tapering rachis. The barbs are very long and
branch out from the posterior edges of the laminae. The rami are
slightly inclined toward the rachidial ridge and bear relatively
straight proximal barbules whereas the distal barbules are more curved.
The sample is too thick to fully observe pigment distribution, but the
apparent color for the entire feather is pale brown.
DIP-V-17121 (Fig. 8d,
e) contains a large botanical inclusion that obscures much of a feather
with rachis width decreasing gradually toward the apex. The angle
between the laminae and the rami is about 35 degrees. The left vane (in
dorsal view) is formed by very long barbs, which are much more elongate
than those forming the right vane, even after accounting for truncation
at the edge of the amber piece (Fig. 8d).
There is a high density of barbules on all feather sections, and the
terminal section of each barb bears proximal barbules decreasing in
length until they disappear just at the tip. The pennulum that is
present in the distal barbules is about one-third or less of the
barbule’s total length. Well-visible pigmentation produces a dark brown
apparent coloration to the feather and, at a finer scale, pigmentation
is concentrated in discreet areas within the barbules (Fig. 8e).
The fragment of oxidized feather in DIP-V-17138 (Figs. 1e and 8f)
preserves part of the mid-length of a RDF with a particularly deep
rachis and clearly visible rachidial ridge. The vanes are oriented
toward the midline of the feather and the rami origination points are
located on the posterior edges of the laminae. Thick barbs are present,
and these contain simple, blade-shaped (reduced) barbules. Oxidation of
the amber imparts a reddish coloration, but aside from this taphonomic
feature, the feather appears to preserve a pale brown overall color.
DIP-V-17127 (Figs. 2h and 8g)
contains a fragment of feather that underwent distortion. The thin
rachidial ridge is barely preserved apically and appears to have been
offset to the right (in ventral view) compared to the basal portion.
Barbs branch from different locations on either side of the main shaft:
slightly below the posterior edge of the lamina and exactly on the edge
(Fig. 2h).
Rami are blade-shaped; and the apical portion of the barbs possesses
proximal barbules decreasing in length to become shorter than distal
ones, progressively disappearing at the tip. Barbules are present on the
laminae as well as the barbs. Overall, the specimen is preserved with
an apparent brown pigmentation.
The feather in DIP-V-16207 (Figs. 2f and 8h)
is poorly preserved, partially fragmented, degraded, and hidden by
milky amber, making observations difficult. However, the characteristic
deep C-shaped rachis with rachidial ridge is observable. Dark brown
pigmentation is present throughout the barbs and rachis of this sample. A
cross-section of the rachis provided material for SEM imaging (Fig. 2f).
DIP-V-16113 (Fig. 9a)
contains the tip of a feather. The study of the specimen is strongly
hindered by numerous syninclusions, bubbles, and resin flows. The rachis
is strongly curved apically and bears flexible barbs with blunt apices
similar to DIP-V-17109. Like DIP-V-17109, the barbules are also locally
aggregated to form clumps.
Fig. 9
Diagnostic and putative RDF fragments. a DIP-V-16113 overview and close-up of the tip of the feather (inset); b DIP-V-17232 overview with prominent rachidial ridge and deep C-shaped rachis; c DIP-V-15130 overview; d DIP-V-15137 overview; e DIP-V-15141 overview; f DIP-V-15142 overview; g DIP-V-15159 overview; h DIP-V-16178 overview, arrowheads mark lateral margins of rachis at both ends. Scale bars = 2 mm in (a and h); 1 mm in (b); 5 mm in (c–g)
The feather fragment in DIP-V-17232 (Fig. 9b)
encompasses part of the vaned section of a RDF. Its broad rachis is
deeply concave in cross-section, and it bears a prominent rachidial
ridge. The dark, mottled appearance of the rachis, and the darker
rachidial ridge appear to reflect original pigment distribution, as
opposed to being taphonomic features. Barbs within DIP-V-17232 have
blade-shaped rami that attach to the lateral surface of the rachis to
form laterally directed vanes. Barbules have a pale brown apparent color
because of diffuse pigmentation, with faint areas of reduced
pigmentation along the proximal margin of each internode highlighting
segmentation within the barbules. Both proximal and distal barbules have
a reduced, blade-shaped morphology, and faint traces of hooklets are
visible on the distal barbules. Proximal barbules do not continue to the
apex of each barb, leaving an acute tip with a short section bearing
only distal barbules.
3.2.5 Putative RDF fragments
Additional
samples have been identified as potential RDFs, but limited
preservation does not allow confident identifications. DIP-V-15130 (Fig.
9c)
contains an oxidized fragment of feather with numerous transverse
cracks and abundant pyrite. Apparent brown reddish coloration in this
sample is due to the taphonomic oxidation. A tapering, deep, C-shaped
rachis with a rachidial ridge allows placement among the RDFs with some
confidence. Short barbs within this feather appear to branch from
slightly below the posterior margins of the laminae. DIP-V-15137 (Fig. 9d)
contains a fragment of feather trapped within a piece of amber with
abundant syninclusions and organic matter, as well as clumps of
trichomes from plants. The poorly preserved feather in DIP-V-15141 (Fig.
9e)
is partially fragmented, with a strong distortion. Rami (particularly
in one vane) are strongly coiled as are some barbules at the tip of the
feather. Basally, at the edge of the piece of amber, a portion of a
C-shaped rachis with a faint ridge is observable, suggesting an RDF
identification. DIP-V-15142 (Fig. 9f)
contains a fragment of feather with a rachis that is slightly tapering,
and a blade-like rachidial ridge. Barbs branch laterally, low on the
sides of the rachis. Deposits of pyrite are concentrated on the rami and
the rachis. DIP-V-15159 (Fig. 9g)
contains a feather that is sectioned basally and apically at the
surface of the amber piece. Abundant deposits of pyrite complicate the
study of the specimen. The rachis tapers apically, and the vanes are
folded toward the anterior mid-line. DIP-V-16178 (Fig. 9h)
contains a fragment of feather at the periphery: the rachis was
partially polished away. A few long barbs are curved and oriented toward
the inner part of the sample. In the apical portion of the barbs, the
proximal barbules decrease in length and disappear at the tip. A loosely
confined brown pigmentation is preserved, with pale spots outlining the
basal cells of the barbules, and hooklets are present.
The
differences observed in RDF cross-sectional morphology would have
produced a range of flexural stiffness without differences in the
material properties of keratin (Bonser and Purslow 1995). Some of the specimens have bent or folded because of interactions with resin flows (Figs. 4d, 5c and 7b), indicating that the open rachis may have bent more easily than the closed rachis found in modern racket plumes (Bleiweiss 1987) or normal flight feathers (Lucas and Stettenheim 1972).
Normal pennaceous flight feathers from enantiornithines do not exhibit
the same degree of flexion in amber, except for at their very tips (Xing
et al. 2016a, 2017).
Although RDFs display asymmetry, their microstructure indicates an
ornamental role. At most, the barbules in the RDFs have reduced hooklets
that would not form strong connections between barbs and produce rigid
vanes (Figs. 3c and 5f).
Coupled with their C-shaped or flattened rachis profiles, poor closure
of the vanes would have yielded feathers with limited aerodynamic
properties but perhaps effective as lightweight tail streamers. The lack
of pith, and a rachis that only forms three-quarters of a cylinder in
cross-section may have been bolstered by thickened sections of rachis
near the rachidial ridge and posterior margins of the laminae, but fully
testing the functional constraints on these feathers is beyond the
scope of the present study.
In addition to the evidence from compression fossils (e.g., O’Connor et al. 2012; Wang et al. 2014),
structural evidence from amber supports RDFs as a means of visual
display. In modern birds, ornamental feathers are colorful and play an
important role in behaviors such as courtship (Hill 2006).
The high proportion of specimens in amber that are recovered as pairs
with no sign of an adjacent corpse, or with surrounding feather patterns
that seem to indicate a striking contact with the resin surface, would
suggest that these feathers are easily removed. The apparent ease of
removal and muted colors observed in amber RDFs may indicate a
sacrificial role in defense, as well as usefulness in visual signaling
(Peters and Peters 2009).
The reduced amount of material involved in building an elongate RDF
with an open and thin-walled rachis may have helped to reduce the
energetic costs of producing feathers that were in many cases as long as
the total body length of their bearers (Carvalho et al. 2015).
One caveat associated with easy removal is that it may limit the
usefulness of RDFs in recognizing the sexes of compression fossils,
assuming that RDFs were a dimorphic feature within any of their bearers
(Chiappe et al. 2008; Peters and Peters 2009; O’Connor et al. 2012).
4.2 Evolutionary development
The unusual morphology of RDFs can be accommodated within accepted follicle collar development patterns (Prum 1999),
but requires greater variation in the production of new tissue than in
modern birds. Production must halt along the posterior margin of the
follicle in RDFs to create feathers that have an open rachis on their
ventral surface. In the amber sample set that is currently available,
the laminae appear to remain open all the way to the follicle (e.g.,
Fig. 4f,
i), but in situ feathers are required to observe if the follicle collar
is closed at the skin surface. Based on comparison between
well-preserved RDFs and normal flight feathers in Early Cretaceous
compression fossils, RDFs were inferred to represent secondarily
modified normal pennaceous flight feathers (O’Connor et al. 2012; Wang et al. 2014).
While the fact they are pennaceous feathers is not debatable at this
point, the detailed morphology gleaned from amber specimens reveals
striking differences that reopen the question of whether this morphotype
evolved from a normal pennaceous feather, in which the rachis is hollow
and pith filled, or whether it has an independent evolutionary pathway.
Based on the normal rachis morphology of the remiges in Burmese amber
enantiornithines, it is likely that the medial stripe observed in RDFs
and that in unmodified remiges and rectrices in compression fossils is
not homologous. If homologous, it may suggest that the superficially
‘modern’ remiges and rectrices in Jehol ornithothoracines in fact
represent a primitive stage in feather evolution. Ultimately, this
question will require additional specimens with exceptional preservation
to address.
4.3 Habitat evidence
Many
of the amber specimens in this study have been polished into thin
pieces that are yellow and very clear. However, some samples are very
rich in syninclusions, such as particulates, trichomes, insects and
botanical inclusions. DIP-V-17109, for example, is very rich in insect
and particulate inclusions (Fig. 5a).
DIP-V-16186 and DIP-V-17153 contain more diagnostic plant remains—fern
sporangia with the “catapult” structure characteristic of the polypod
ferns. These syninclusions provide some associations between the RDF
feather inclusions and plant groups beyond the trees within
Araucariaceae or Dipterocarpaceae that are thought to have produced the
resin (Ross et al. 2010).
However, there are no consistent indicators for a particular habitat or
height within the forest canopy (e.g., ‘litter amber’; Perrichot 2004) found among the RDF sample set.
Cretaceous
birds retained numerous primitive feather morphotypes, most of which
are better understood through preservation in amber. The new RDFs
preserved in three dimensions within amber clearly indicate that this
feather type had an open rachis much different from the rachises found
in modern bird flight feathers. An open rachis may have been a
cost-saving measure in the production of feathers that could grow as
long as their bearers. The RDF morphotype may also have evolved
specifically to create streamers that were elongate but lightweight. It
is possible that these feathers represent a primitive holdover (prior to
the development of a full rachis), or they may represent a secondary
loss of the tubular rachis. Notably, flexion was not previously inferred
from compression fossils found in rocks of Early Cretaceous age (based
on the straight and unbent preservation of the feathers in all known
specimens in the Jehol Biota). This may suggest that the flexibility is a
derived feature, or that the feathers are only able to flex ventrally
under significant force. The relative abundance of paired RDFs in amber
suggests that many of the samples in Burmese amber were originally
paired tail feathers, and that these were plucked without the entire
bird becoming ensnared in resin. If the feathers were easy to remove,
their role may have gone beyond visual signalling, also serving as a
defensive decoy, and their presence or absence in compression fossils
should be used as a sexual indicator with caution.
Christopher
Somers (University of Regina), Ray Poulin and Danae Frier (RSM) for
discussions and research support; and Nathan Gerein (University of
Alberta) for SEM assistance.
Funding
We thank National Natural
Science Foundation of China (41772008), Fundamental Research Funds for
the Central Universities (2652017215), National Geographic Society,
U.S.A. (EC0768-15), Natural Sciences and Engineering Research Council,
Canada (2015-00681), RSM and Paul Riegert scholarships (to P.C.) for
support.
Availability of data and materials
All data generated or analyzed during this study are included in this published article.
Authors’ contributions
LX,
PC, JO, and RCM designed the project, performed the research, and wrote
the manuscript. All the authors contributed to reviewing the
manuscript.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open AccessThis article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/),
which permits unrestricted use, distribution, and reproduction in any
medium, provided you give appropriate credit to the original author(s)
and the source, provide a link to the Creative Commons license, and
indicate if changes were made.
Bleiweiss, R. 1987. Development and evolution of avian racket plumes: fine structure and serial homology of the wire. Journal of Morphology 194: 23–39.View ArticleGoogle Scholar
Bonser, R.H.C., and P.P. Purslow. 1995. The Young’s modulus of feather keratin. Journal of Experimental Biology 198: 1029–1033.Google Scholar
Carvalho,
I.S., F.E. Novas, F.L. Agnolín, M.P. Isasi, F.I. Freitas, and J.A.
Andrade. 2015. A Mesozoic bird from Gondwana preserving feathers. Nature Communications 6: 7141.View ArticleGoogle Scholar
Chen, P., Z. Dong, and S. Zhen. 1998. An exceptionally well-preserved theropod dinosaur from the Yixian formation of China. Nature 391: 147–152.View ArticleGoogle Scholar
Chiappe, L.M., J. Marugán-Lobón, S. Ji, and Z. Zhou. 2008. Life history of a basal bird: Morphometrics of the early cretaceous Confuciusornis. Biology Letters 4: 719–723.View ArticleGoogle Scholar
Delclòs,
X., A. Arillo, E. Peñalver, E. Barrón, C. Soriano, R. López Del Valle,
E. Bernárdez, C. Corral, and V.M. Ortuño. 2007. Fossiliferous amber
deposits from the cretaceous (Albian) of Spain. Comptes Rendus Palevol 6: 135–149.View ArticleGoogle Scholar
Dove, C.J. 2000. A descriptive and phylogenetic analysis of plumulaceous feather characters in Charadriiformes. Ornithology Monographs 51: 1–63.Google Scholar
Grimaldi,
D.A., and G.R. Case. 1995. A feather in amber from the Upper Cretaceous
of New Jersey. American Museum Novitates 3126: 1–6.Google Scholar
Grimaldi,
D.A., M.S. Engel, and P.C. Nascimbene. 2002. Fossiliferous Cretaceous
amber from Myanmar (Burma): its rediscovery, biotic diversity, and
paleontological significance. American Museum Novitates 3361: 1–71.Google Scholar
Hill, G.E. 2006. Female mate choice for ornamental coloration in birds. In Bird coloration: function and evolution, ed. G.E. Hill and K.E. McGraw, 137–200. Cambridge: Harvard University Press.Google Scholar
Hu,
H., J.K. O’Connor, and Z. Zhou. 2015. A new species of Pengornithidae
(Aves: Enantiornithes) from the Lower Cretaceous of China suggests a
specialized scansorial habitat previously unknown in early birds. PLoS
One 10: e0126791.Google Scholar
Ji, Q., P.J. Currie, M.A. Norell, and J. Shu-An. 1998. Two feathered dinosaurs from northeastern China. Nature 393: 753–761.View ArticleGoogle Scholar
Ji, Q., and S.-A. Ji. 1996. On the discovery of the earliest fossil bird in China (Sinosauropteryx gen. nov.) and the origin of birds. Chinese Geology 233: 30–33.Google Scholar
Lucas, A.M., and P.R. Stettenheim. 1972. Avian anatomy integument. Washington, D.C.: United States Department of Agriculture.Google Scholar
McKellar,
R.C., B.D.E. Chatterton, A.P. Wolfe, and P.J. Currie. 2011. A diverse
assemblage of Late Cretaceous dinosaur and bird feathers from Canadian
amber. Science 333: 1619–1622.Google Scholar
McKittrick,
J., P. Chen, S. Bodde, W. Yang, E. Novitskaya, and M.A. Meyers. 2012.
The structure, functions, and mechanical properties of keratin. The Journal of the Minerals, Metals & Materials Society 64: 449–468.View ArticleGoogle Scholar
O’Connor,
J.K., L.M. Chiappe, C.M. Chuong, D.J. Bottjer, and H. You. 2012.
Homology and potential cellular and molecular mechanisms for the
development of unique feather morphologies in early birds. Geosciences 2: 157–177.View ArticleGoogle Scholar
Peñalver,
E., A. Arillo, X. Delclòs, D. Peris, D.A. Grimaldi, S.R. Anderson, P.C.
Nascimbene, and R. Pérez-de la Fuente. 2017. Ticks parasitised
feathered dinosaurs as revealed by Cretaceous amber assemblages. Nature
Communications 8: 1924.Google Scholar
Perrichot,
V. 2004. Early Cretaceous amber from South-Western France: insight into
the Mesozoic litter fauna. Geologica Acta 2: 9–22.Google Scholar
Perrichot,
V., L. Marion, D. Néraudeau, R. Vullo, and P. Tafforeau. 2008. The
early evolution of feathers: Fossil evidence from Cretaceous amber of
France. Proceedings of the Royal Society of London B, Biological
Sciences 275: 1917–1202.Google Scholar
Peters, W.S., and D.S. Peters. 2009. Life history, sexual dimorphism and ‘ornamental’ feathers in the Mesozoic bird Confuciusornis sanctus. Biology Letters 5: 817–820.View ArticleGoogle Scholar
Prum, R.O. 1999. Development and evolutionary origin of feathers. Journal of Experimental Zoology 285: 291–306.View ArticleGoogle Scholar
Ross,
A., C. Mellish, P. York, and B. Crighton. 2010. Burmese amber. In
Biodiversity of fossils in amber from the major world deposits, ed. D.
Penney, 208–235. Manchester: Siri Scientific Press.Google Scholar
Schlee, D., and W. Glöckner. 1978. Bernstein: Bernsteine und Berstein-Fossilien. Stuttgarter Beiträge zur Naturkunde - Serie B 8: 1–72.Google Scholar
Shi,
G., D.A. Grimaldi, G.E. Harlow, J. Wang, J. Wang, M. Yang, W. Lei, Q.
Li, and X. Li. 2012. Age constraint on Burmese amber based on U–Pb
dating of zircons. Cretaceous Research 37: 155–163.View ArticleGoogle Scholar
van
der Reest, A.J., A.P. Wolfe, and P.J. Currie. 2016. A densely feathered
ornithomimid (Dinosauria: Theropoda) from the Upper Cretaceous Dinosaur
Park Formation, Alberta, Canada. Cretaceous Research 58: 108–117.Google Scholar
Wang,
M., X. Zheng, J.K. O’Connor, G.T. Lloyd, X. Wang, Y. Wang, X. Zhang,
and Z. Zhou. 2015. The oldest record of Ornithuromorpha from the Early
Cretaceous of China. Nature Communications 6: 6987.Google Scholar
Wang,
X., J.K. O’Connor, X. Zheng, M. Wang, H. Hu, and Z. Zhou. 2014.
Insights into the evolution of rachis dominated tail feathers from a new
basal enanthiornine (Aves: Ornithothoraces). Biological Journal of the Linnean Society 113: 805–819.View ArticleGoogle Scholar
Xing,
L., R.C. McKellar, M. Wang, M. Bai, J.K. O’Connor, M.J. Benton, J.
Zhang, Y. Wang, K. Tseng, M.G. Lockley, G. Li, W. Zhang, and X. Xu.
2016a. Mummified precocial bird wings in mid-Cretaceous Burmese amber.
Nature Communications 7: 12089.Google Scholar
Xing,
L., R.C. McKellar, X. Xu, G. Li, M. Bai, W.S.I.V. Persons, T.
Miyashita, M.J. Benton, J. Zhang, A.P. Wolfe, Q. Yi, K. Tseng, H. Ran,
and P.J. Currie. 2016b. A feathered dinosaur tail with primitive plumage
trapped in mid-Cretaceous amber. Current Biology 26: 3352–3360.Google Scholar
Xing,
L., J.K. O’Connor, R.C. McKellar, L.M. Chiappe, K. Tseng, G. Li, and M.
Bai. 2017. A mid-Cretaceous enantiornithine (Aves) hatchling preserved
in Burmese amber with unusual plumage. Gondwana Research 49: 264–277.Google Scholar
Xu, X., X. Zheng, and H. You. 2010. Exceptional dinosaur fossils show ontogenetic development of early feathers. Nature 464: 1338–1341.View ArticleGoogle Scholar
Xu,
X., Z. Zhou, R. Dudley, S. Mackem, C.-M. Chuong, G.M. Erickson, and
D.J. Varrichio. 2014. An integrative approach to understanding bird
origins. Science 346: 1253293.View ArticleGoogle Scholar
Zhang, F., and Z. Zhou. 2000. A primitive enantiornithine bird and the origin of feathers. Science 290: 1955–1959.View ArticleGoogle Scholar
Zhang, F., Z. Zhou, and G. Dyke. 2006. Feathers and ‘feather-like’ integumentary structures in Liaoning birds and dinosaurs. Geological Journal 41: 395–404.View ArticleGoogle Scholar
Zhang,
F.C., Z.H. Zhou, X. Xu, X.L. Wang, and C. Sullivan. 2008. A bizarre
Jurassic maniraptoran from China with elongate ribbon–like feathers. Nature 455: 1105–1108.View ArticleGoogle Scholar
Zhang,
Z., Z. Zhou, and M.J. Benton. 2008. A primitive confuciusornithid bird
from China and its implications for early avian flight. Science in China Series D: Earth Sciences 51: 625–639.View ArticleGoogle Scholar
Zheng,
X.T., H.L. You, X. Xu, and Z.M. Dong. 2009. An Early Cretaceous
heterodontosaurid dinosaur with filamentous integumentary structures.
Nature 458: 333–336.Google Scholar













Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.