The evolution of
photosynthesis has been a major driver in eukaryotic diversification.
Eukaryotes have acquired plastids (chloroplasts) either directly via the
engulfment and integration of a photosynthetic cyanobacterium (primary
endosymbiosis) or indirectly by engulfing a photosynthetic eukaryote
(secondary or tertiary endosymbiosis). The timing and frequency of
secondary endosymbiosis during eukaryotic evolution is currently unclear
but may be resolved in part by studying cryptomonads, a group of
single-celled eukaryotes comprised of both photosynthetic and
non-photosynthetic species. While cryptomonads such as Guillardia theta
harbor a red algal-derived plastid of secondary endosymbiotic origin,
members of the sister group Goniomonadea lack plastids. Here, we present
the genome of Goniomonas avonlea—the
first for any goniomonad—to address whether Goniomonadea are
ancestrally non-photosynthetic or whether they lost a plastid
secondarily.
Results
We sequenced the nuclear and mitochondrial genomes of Goniomonas avonlea and carried out a comparative analysis of Go. avonlea, Gu. theta, and other cryptomonads. The Go. avonlea
genome assembly is ~ 92 Mbp in size, with 33,470 predicted
protein-coding genes. Interestingly, some metabolic pathways (e.g.,
fatty acid biosynthesis) predicted to occur in the plastid and
periplastidal compartment of Gu. theta appear to operate in the cytoplasm of Go. avonlea,
suggesting that metabolic redundancies were generated during the course
of secondary plastid integration. Other cytosolic pathways found in Go. avonlea are not found in Gu. theta, suggesting secondary loss in Gu. theta and other plastid-bearing cryptomonads. Phylogenetic analyses revealed no evidence for algal endosymbiont-derived genes in the Go. avonlea
genome. Phylogenomic analyses point to a specific relationship between
Cryptista (to which cryptomonads belong) and Archaeplastida.
Conclusion
We found no convincing genomic or phylogenomic evidence that Go. avonlea
evolved from a secondary red algal plastid-bearing ancestor, consistent
with goniomonads being ancestrally non-photosynthetic eukaryotes. The Go. avonlea
genome sheds light on the physiology of heterotrophic cryptomonads and
serves as an important reference point for studying the metabolic
“rewiring” that took place during secondary plastid integration in the
ancestor of modern-day Cryptophyceae.
The
acquisition of photosynthesis in eukaryotes can be traced back to a
primary endosymbiosis in which a eukaryotic host engulfed and
assimilated a photosynthetic cyanobacterium, which ultimately became the
plastid (chloroplast) [1, 2].
Canonical “primary” plastids are surrounded by two membranes and are
generally thought to have evolved on a single occasion in the common
ancestor of Archaeplastida, a tripartite eukaryotic “supergroup”
comprised of Viridiplantae (also known as Chloroplastida), Rhodophyta
(Rhodophyceae), and Glaucophyta [3, 4, 5].
Eukaryotes have also acquired photosynthesis indirectly on multiple
occasions via “secondary” (i.e., eukaryote-eukaryote) endosymbiosis.
Indeed, secondary (and in some cases tertiary) endosymbiosis is thought
to have given rise to plastids scattered amongst the stramenopiles,
alveolates, rhizarians, euglenozoans, haptophytes, and cryptomonads [6].
The latter lineage is divided into two clades, the plastid-bearing,
mostly photosynthetic Cryptophyceae and the heterotrophic Goniomonadea.
The evolutionary distinctness of these two clades makes for an
interesting case study with which to understand the transition from a
plastid-lacking eukaryote to a photosynthetic, secondary plastid-bearing
organism.
Guillardia theta and the recently described Goniomonas avonlea [7] are representatives of plastid-bearing and plastid-lacking cryptomonads [5, 8], respectively. Together with several paraphyletic plastid-lacking lineages, including katablepharids and Palpitomonas, cryptomonads constitute a clade known as Cryptista [9, 10].
The position of Cryptista on the eukaryotic tree of life is a point of
contention. Some phylogenomic studies have placed it sister to
Haptophyta (e.g., [11]),
with the Cryptista-Haptophyta clade itself branching either next to the
SAR supergroup (Stramenopiles, Alveolata, Rhizaria; e.g., [12]) or the Archaeplastida (e.g., [13]).
Other studies have suggested that Cryptista and Haptophyta are not
specifically related, with the former branching within the
Archaeplastida [14]. Our knowledge of Cryptista and their evolutionary history has suffered from a paucity of genomic data [15]. Only one species, Gu. theta,
which has been studied mainly for its plastid and nucleomorph (the
vestigial nucleus of the red alga acquired by secondary endosymbiosis) [16, 17] has had its nuclear genome sequenced [18].
The diversity of plastid-lacking species within cryptomonads and, more
broadly, Cryptista, has received relatively little attention.
The
transition from a heterotrophic, aplastidic cell to a plastid-bearing
one is associated with the acquisition of a wide range of metabolic
capabilities, such as photosynthesis and novel amino acid biosynthetic
capacities [19].
The acquisition of photosynthesis and carbon fixation by a
heterotrophic protist impacts the regulation of many of its metabolic
pathways [20].
In addition, pathways operating in different subcellular compartments
can become partially or completely redundant. This allows the organism
to tinker with the regulation of pathways that may be adapted to a
particular cellular compartment and/or set of metabolites. Endosymbiosis
can also give rise to mosaic metabolic pathways comprised of enzymes
with different evolutionary origins [18, 21].
Proteins may be derived from the host, from the endosymbiont (both
primary and secondary), or as a result of lateral gene transfer (LGT)
from different prokaryotic and eukaryotic organisms. Understanding how
cells adapt from living in a solitary state to having another organism
within it is fundamental to understanding the evolution of
plastid-bearing organisms.
We have sequenced the nuclear genome and transcriptome of the plastid-lacking goniomonad Go. avonlea CCMP3327 [7]
with the goal of shedding light on its physiology and, more generally,
the metabolic transformation that accompanied the transition from
heterotrophy to phototrophy in its plastid-bearing sister taxa. Using
comparative genomics and phylogenomics, we found little evidence for a
photosynthetic ancestry in Go. avonlea
and show that the acquisition of a plastid in an ancestor of
present-day Cryptophyceae resulted in extensive reshuffling of metabolic
pathways. Annotation of carbohydrate-active enzymes (CAZymes) [22]
including glycosyltransferases (GTs), glycoside hydrolases (GHs),
polysaccharide lyases (PLs), and carbohydrate esterases (CEs) allows us
to make several predictions about the lifestyle of Go. avonlea and other goniomonads, including the possibility that they feed on multiple organisms, including eukaryotic algae.
Cell culture, nucleic acid preparation, and genome sequencing
Goniomonas avonlea CCMP3327 was grown in ESM medium [23]
supplemented with ATCC’s 1525 Seawater 802 medium. One day prior to
harvesting, a dose of Penicillin-Streptomycin-Neomycin antibiotic
mixture (Thermo Fisher cat #15640055) was administered in order to
reduce the number of co-cultured bacteria. Cells were harvested in two
steps. First, liquid culture was filtered through a 2-μm pore-sized
polycarbonate membrane disc in order to deplete bacterial cells; the
remaining protist cells were re-suspended in artificial seawater and
transferred to a falcon tube. Cells were pelleted by centrifugation at
3000 RCF for 8 min. DNA was extracted using a PureLink® Genomic DNA Kit
(Thermo Fisher Scientific, cat# K182001). For RNA preparation, cells
were lysed and phase-separated using TRIzol™ reagent (Thermo Fisher
Scientific, cat #15596018), followed by the use of the RNeasy Mini Kit
(QIAGEN, cat #74104) for precipitation, washes, and elution.
DNA
and RNA samples were sent to Génome Québec and the Beijing Genomics
Institute (BGI) for library preparation and sequencing on the Illumina
HiSeq2000 platform. At Génome Québec, genomic data were generated from a
short-insert library, while 2 kb and 6 kb mate pair libraries were
sequenced at the BGI. A total of 209,988,904, 12,244,898, and 35,906,071
forward and reverse reads, up to 100 bp in length, were generated for
the short-insert, 2 kb, and 6 kb mate pair libraries, respectively. For
the transcriptome, 62,428,409 forward and reverse reads were sequenced,
up to 100 bp in length, from a library prepared with the TruSeq protocol
at Génome Québec.
Genome assembly, gene prediction, and quality control
Transcriptome reads were quality trimmed using Trimmomatic [24] and assembled de novo with Trinity [25]. The genome was assembled with ALLPATH-LG [26], Abyss [27], Minimus2 [28] and Ray [29]. We considered the N50 values of the two “best” assemblies (Abyss and ALLPATHS-LG) and used BOWTIE2 [30] coupled with ALE [31] and CGAL [32]
to evaluate which assembly was optimal for our purposes. The
ALLPATHS-LG assembly was selected and subjected to a blastn analysis [33]; contigs with bacterial hits with E-values lower than 1e− 50 were considered bacterial and removed. We then predicted protein-coding genes using both Augustus [34, 35] and PASA [36],
which allowed correction of gene models using transcriptome data. To
further reduce the chance of bacterial contamination, we carried out
blastp searches of our predicted proteins against NCBI nr (ftp://ftp.ncbi.nlm.nih.gov/blast/db/) and the Marine Microbial Eukaryote Transcriptome Sequencing Project database (ftp://ftp.imicrobe.us/camera/) [37].
For each protein sequence, we took the first 10 hits and considered the
protein to be from Eukaryota if > 60% of the hits were eukaryotic,
or bacterial or archaeal if > 60% of the hits were to Bacteria or
Archaea. Sequences that did not pass either threshold were assessed
manually. In such cases, genes were mapped to their contigs and if
> 60% of the gene models on the contig were eukaryotic, we considered
the contig to be derived from the Goniomonas avonlea nuclear genome.
To identify as many protein-coding genes as possible and to ensure their full length, we predicted all ORFs from the Go. avonlea
transcriptome, generating six frame translations for each transcript.
From the pool of possible ORFs, we took the four longest translations
and blasted them against the nr and MMETSP databases. All translated
transcripts with a hit below 1e− 05 were kept. To these
transcriptome-derived sequences, we added protein sequences predicted
from the genomic data using Augustus and PASA. The added sequences were
those that had a hit < 1e− 05 against the nr or MMETSP
databases and that did not already match proteins from the transcriptome
dataset with sequence identity > 90%. The resulting set of 18,429
protein coding sequences, used for all subsequent analyses, represents
the union of predicted gene models and predicted ORFs and represents a
refined set of protein sequences demonstrably from Goniomonas and likely to have homologs in other organisms.
We assessed genome “completeness” using BUSCO (v1; [38]), which is based on a set of 429 protein-coding genes purported to be universally present in eukaryotes as single copies [37, 38]. The 18,429 Go. avonlea proteins were analyzed; the BUSCO results were compared to those obtained for Gu. theta and the amoebozoan Dictyostelium discoideum.
Orthologous protein annotation and KOG classification
For Go. avonlea, Gu. theta, Bigelowiella natans, Emiliania huxleyi, Adineta vaga, and Arabidopsis thaliana, we clustered orthologous sequences using OrthoVenn (http://www.bioinfogenome.net/OrthoVenn/) [39], with E-value and inflation value settings at 1e− 5
and 1.5, respectively. In addition, we compared the size and diversity
of KOG functional categories (EuKaryotic Orthologous Groups) inferred
from both the Go. avonlea and Gu. theta genomes using the WebMGA server (http://weizhong-lab.ucsd.edu/webMGA/server/kog/) with an E-value cut-off of 1e− 03 [40].
Protein annotation and sub-cellular localization prediction
Protein annotation was performed using KOBAS [41]; proteins that were not annotated using this approach were analyzed using GhostKOALA [42]. Annotations were then used to generate KEGG metabolic pathway maps (http://www.genome.jp/kegg/tool/map_pathway.html) [43].
In order to predict the sub-cellular locations of Go. avonlea
proteins, we first selected 13,508 sequences inferred from our genome
assembly that (i) start with a methionine and (ii) match the amino
(N)-termini of proteins in our final set of 18,429 proteins, reasoning
that proteins derived from our genomic (and not transcriptomic) data
were more likely to possess full length N termini. Given the uncertainty
of whether or not Go. avonlea
and other goniomonads evolved from a plastid-bearing ancestor, we
carried out different predictions using a combination of PredSL [44], TargetP [45], and Predotar [46] under the “plant” and “non-plant” modes (Additional file 1). Considering the formal possibility that Go. avonlea could, like Gu. theta, have a plastid acquired by secondary endosymbiosis, we used SignalP 4.1 [47, 48] coupled with ASAFind [49] to predict periplastidial compartment (PPC) and plastid proteins. For Gu. theta, the predicted protein coding gene set from Curtis et al. [18] was used, as were the signal peptide predictions for the purposes of comparison with Go. avonlea.
Annotation of carbohydrate-active enzymes (CAZymes)
We performed a manual annotation of CAZymes [22] using a mix of BLAST [33] and HMM searches [50], similar to that done previously for Gu. theta [18].
To assess the similarity between the two species across CAZyme
families, we generated heat maps derived from an average linkage
hierarchical clustering based on Bray-Curtis dissimilarity matrix
distances and Ward’s method [51, 52, 53, 54]. The phylogenetic heat maps were generated with Rstudio software (https://www.rstudio.com/) using vegan in the R package (http://cc.oulu.fi/~jarioksa/softhelp/vegan/html/vegdist.html) [55] with vegdist and hclust commands.
Phylogenetic analysis of “algal” genes
We sought to identify putative algal-derived homologs in the Go. avonlea genome by comparing its gene/protein set to that of Gu. theta
and other algae. More specifically, we used blastp to search a custom
database of 508 proteins predicted to be the product of endosymbiotic
gene transfer (EGT) in Gu. theta [18]. For each Go. avonlea protein with a significant hit to this database (E-value < 1e− 10), we used DIAMOND with the “more sensitive” option [56] to retrieve the top 2000 homologs above an E-value cut-off of 1e− 10 from the nr and MMETSP [37] databases. Paralogs in the Go. avonlea
candidate-EGT set were then identified by pairwise comparison of
DIAMOND outputs; if two queries had > 50% overlap in hits they were
considered paralogous and merged. Candidate Go. avonlea EGTs were annotated using InterPro [57] and their subcellular localizations were predicted as above.
Single-gene/protein trees were generated from alignments initially produced using MAFFT (version 7.205 [58]). Ambiguously aligned regions were removed using BMGE (version 1.1 [59])
with the BLOSUM30 scoring matrix and a block size of 4; trimmed
alignments shorter than 50 amino acids were discarded. For the remaining
candidates, an approximately maximum likelihood phylogeny was generated
using FastTree [60]
and used in an in-house tree-trimming script to reduce taxonomic
redundancy in each dataset. Reduced datasets were then re-aligned using
MAFFT-linsi (version 7.205 [58]),
trimmed as above, and filtered to discard alignments shorter than 70
amino acids. Maximum-likelihood (ML) phylogenies were inferred for each
remaining candidate in IQ-TREE (Version 1.5.5 [61]) under the LG4X substitution model [62] with 1000 ultra-fast bootstrap approximations (UFboot) [63]. The resulting trees were manually evaluated and sorted based on the topology of the Go. avonlea and Gu. theta
proteins in relation to each other and to sequences from various
combinations of primary and secondary plastid-bearing photosynthetic
lineages.
Additional
genes/proteins of particular interest (for example, the CAZymes
glycosyltransferase 28 and glucan water dikinase) were investigated on a
case-by-case basis using a similar approach as above and as described
in several other studies [64, 65]. In these cases, homologs to predicted Go. avonlea
genes/proteins were identified in various additional
genomic/transcriptomic datasets, sequence redundancy was reduced using a
combination of manual inspection and an automated treetrimmer analysis [66] and “final” alignments were produced with MUSCLE [67].
Phylogenomics
To
investigate the phylogenetic position of Cryptista on the eukaryotic
tree of life, a 250-marker gene, un-aligned dataset consisting of 150
operational taxonomic units (OTUs) corresponding to Burki et al. [14] was obtained from the Dryad Digital Repository [68].
The number of OTUs was systematically reduced to decrease the
complexity of phylogenetic analyses while maintaining taxonomic
diversity and minimizing missing data. Proteins predicted from the Go. avonlea transcriptome data were added to the dataset to increase marker gene coverage for the Goniomonadea. Go. avonlea
homologs were identified using blastp with any Cryptista sequence (if
available) or the first sequence in the marker gene set as the query;
the best hit (E-value < 1e− 10) in Go. avonlea was added to the dataset. Each marker gene/protein was aligned using MAFFT-linsi (version 7.205; [58]), and ambiguously aligned sites were removed using BMGE (version 1.1 [59]). Single gene trees were inferred using ML methods in IQTREE (Version 1.4.3 [61]) under the substitution model LG4X [62] with 1000 UFboot [63]
and manually inspected to identify any obvious potential artifacts
(e.g., long branch attractions). Marker genes/proteins were then
realigned and subject to block removal prior to concatenation. The
resulting supermatrix was used to infer a ML phylogeny in IQTREE
(Version 1.4.3 [61]) using the model LG + C60 + F + PMSF [69] (selected according to the Bayesian Information Criterion (BIC) based on the outcome of a model test implemented in IQTREE [70]) with 100 standard bootstrap iterations.
In
order to further investigate the phylogenetic position of Cryptista,
phylogenetic trees were inferred (as above) based on modified versions
of the supermatrix in which (i) sequences from plastid-bearing
cryptistan taxa (i.e., Cryptophyceae) were removed from the dataset and
(ii) PhyloMCOA [71]
was used to identify and remove discordant genes in each OTU based on
phylogenetic positioning across single-gene trees (using nodal
distances) and multiple co-inertia analysis (MCOA). To evaluate the
significance of differences in branch support, the standard error of
bootstrap values was used with a 95% confidence interval. To explore
alternative signals emerging from Cryptista in the marker gene dataset,
183 of the 250 genes (those that contained a homolog in Go. avonlea
and at least one other cryptomonad) were randomly partitioned into four
equally sized bins. Each bin of marker genes was concatenated and used
to infer a phylogeny based on ML methods in IQTREE (Version 1.4.3; [61])
under the model LG + C20 + F. This process was repeated 25 times,
resulting in 100 randomly generated marker gene subset trees; for each
tree, the phylogenetic position of Cryptista was manually evaluated.
The Goniomonas avonlea nuclear and mitochondrial genomes
We sequenced the Go. avonlea
nuclear genome to a depth of ~ 24× coverage. In part due to the
presence of repetitive sequences, the assembly is highly fragmented. For
the final assembly, we retained contigs at least 500 bps in length
resulting in 31,852 contigs (N50 = 3831) totalling 91.5 Mb and a GC
content of 55.2% (Table 1).
From our initial assembly, 33,470 genes were predicted; further
investigation revealed that this number was artificially inflated due to
assembly fragmentation. We thus merged protein-coding genes predicted
from the genome with those inferred from transcriptome data (see “Methods”). This resulted in a set of 18,429 non-redundant protein-coding genes. When analyzed with BUSCO [38],
our protein sequence data set was predicted to be 69% “complete,” 20%
“fragmented,” and 9.7% “missing”; this is comparable to the previously
sequenced genome of Gu. theta, which was inferred to be 78% “complete,” 12% “fragmented,” and 8.8% “missing” (Additional file 2: Table S1). This suggests that despite the level of assembly fragmentation, the Go. avonlea protein coding gene set is similar in terms of completeness to that of Gu. theta;
conclusions about the presence/absence of metabolic pathways in the two
genomes (see below) are probably not adversely affected by missing
data. It should also be noted that in general such analyses are limited
by a distinct lack of knowledge of cryptomonads and their large
phylogenetic distance from the organisms used to create the BUSCO
reference dataset. For reference, the well-annotated genome of the
amoebozoan protist D. discoideum is inferred to be 5.1% “missing” using BUSCO (Additional file 2: Table S1).
Table 1
General genome features for Guillardia theta and Goniomonas avonlea
Guillardia theta
Goniomonas avonlea
Assembly size
87.1 Mb
91.5 Mb
# scaffolds
669
31,852
# contigs
5126
31,852
N50 scaffolds
40,445 bp
3831 bp
GC%
52.9
55.2
# of protein coding genes
24,822
33,470
# of introns
132,885
112,740
Percentage of genes with introns
79%
84%
Number of forward genes
12,482
16,836
Number of reverse genes
12,441
16,638
Average size of gene (nt)
1863
1626
Intron size average (nt)
106
171
Intron size mode (percent of total)
47 (5.1%)
46 (4.7%)
Average # of introns per gene
5.3
3.5
Analysis of orthologous groups of proteins shared between the cryptomonads Go. avonlea and Gu. theta, as well as the rhizarian Bigelowiella natans, the haptophyte Emiliania huxleyi, the rotifer Adineta vaga, and the model land plant Arabidopsis thaliana, shows that, as expected, Go. avonlea and Gu. theta share more orthologous protein families with each other (4321 in total) than they do with other organisms (Go. avonlea shares 3647, 3441, 3173, and 2955 protein families with B. natans, E. huxleyi, A. vaga, and A. thaliana, respectively; Fig. 1a). Nevertheless, comparison of KOGs present in both Go. avonlea and Gu. theta reveals differences in the size and complexity of certain KOG functional categories (Fig. 1b).
For example, KOG categories corresponding to cytoskeleton and
intracellular trafficking, secretion, and vesicular transport are more
abundant in Go. avonlea. Such differences may in part be due to the obligate phagotrophic lifestyle of Go. avonlea (see below). In contrast, Gu. theta appears somewhat enriched (relative to Go. avonlea)
in functions associated with translation, ribosomal structure and
biogenesis, as well as cell cycle control/division and chromosome
partitioning (Fig. 1b).
Fig. 1
Comparative genomics of Goniomonadea, Cryptophyceae, and other eukaryotes. a Venn diagram showing orthologous clusters shared between the goniomonad Goniomonas avonlea (red), the cryptophyte Guillardia theta (green), the rhizarian Bigelowiella natans (blue), the haptophyte Emiliania huxleyi (yellow), the opisthokont Adineta vaga (orange), and the land plant Arabidopsis thaliana (brown). Go. avonlea shares 4321 families with Gu. theta, higher than is shared with other eukaryotes (B. natans (3647), E. huxleyi (3441), A. vaga (3173), A. thaliana (2955)). b KOG classification of proteins in Go. avonlea (brown) and Gu. theta (red). Within most functional categories, the number of proteins in the two organisms is similar. However, Go. avonlea
possesses more proteins in some categories, in particular the
cytoskeleton and the intracellular trafficking, secretion, and vesicular
transport families
We also sequenced and assembled the mitochondrial genome of Go. avonlea,
which at 41.2 kb in size is similar to the circular mapping genomes of
cryptophytes (when repeated sequences are excluded; e.g., [72, 73]) and the linear mtDNA of Palpitomonas bilix [74]. There are also interesting similarities amongst these mtDNAs in terms of gene content (Additional file 2:
Figure S1). When compared to the mitochondrial genomes of other
eukaryotic lineages, all members of Cryptista considered here share very
similar gene repertoires, especially for complexes I-V of the electron
transport chain (Additional file 2: Figure S1): they all have nad1–4, 4L, 5–11, sdh3, 4, cob, cox1–3, atp1, 3, 4, 6, 8, and 9. That said, Go. avonlea and P. bilix share an rpl2 gene that is not present in cryptophyte mtDNAs, and all sequenced cryptophyte mtDNAs possess an rps2 gene [73] that is not present in Go. avonlea or P. bilix. Go. avonlea mtDNA also lacks the tatA and tatC
genes found in other Cryptista mtDNAs, which encode components of the
twin arginine translocator. Of particular note, like cryptophytes, Go. avonlea lacks the mitochondrial ccmA, B, C, and F genes recently found in the mtDNA of P. bilix [74]. These genes encode a bacterial-type cytochrome c
maturation system (“system I”); our data support the hypothesis that
goniomonads use a nucleus-encoded holocytochrome c synthase (HCCS)
system (i.e., “system III”). We searched for, and found, the HCCS gene
in the nuclear genome of Go. avonlea
(comp53045_c0_seq2_6_ORF10_179 and comp39203_c0_seq2_6_ORF3_158). This
confirms the authenticity of such genes found in transcriptome data from
Go. pacifica and the katablepharid Roombia sp. NY0200 [74]. With its mitochondrial-encoded “system I” cytochrome c maturation system, P. bilix
is thus an outlier amongst cryptistan protists, which raises
interesting questions about how the type I and III systems evolved in
these and other organisms (see Nishimura et al. [74]
for discussion). All things considered, our mitochondrial genome
analyses are consistent with phylogenomic data suggesting that although
the organisms that comprise Cryptista are not closely related, they
represent a monophyletic assemblage on the eukaryotic tree of life [9, 10, 14].
Goniomonas avonlea does not have a plastid
On the basis of electron microscopy, Go. avonlea cells do not have any obvious plastid-like internal structures [7].
Nevertheless, with complete genome and transcriptome sequences in hand,
we explored its predicted metabolic pathways as well as putative
TOC-TIC proteins in an effort to detect any hint of evidence for a
cryptic plastid—none was found (Additional file 2: Figure S2). Moreover, we predicted the sub-cellular locations of all of the Go. avonlea
proteins under the following hypothetical scenarios: (i) the organism
does not have a plastid, (ii) it has a cryptic plastid derived from
primary endosymbiosis, or (iii) it has a cryptic plastid of secondary
endosymbiotic origin. In short, we found no evidence supporting the
presence of a plastid of primary or secondary endosymbiotic ancestry;
hundreds of candidate proteins were identified using various search
procedures (e.g., presence of bipartite N-terminal targeting sequences) but closer investigation revealed these to be false positives (Additional file 1 and Additional file 2: Figure S3). This is consistent with previous analyses performed on transcriptome data from Goniomonas pacifica [10] as well as microscopic observations of several Goniomonas strains, including Go. avonlea [7, 75, 76].
Absence of endosymbiotically derived algal genes in Goniomonas avonlea
Curtis et al. [18] identified 508 genes of probable endosymbiont (i.e., algal) ancestry in the Gu. theta
nuclear genome. Many of these endosymbiotic gene transfers (EGTs)
encode proteins that are predicted to have been repurposed and to
function in the host cytosol of Gu. theta or other host-associated compartments; if Go. avonlea
lost a red-algal-derived plastid secondarily, one might thus predict
that at least some of these algal genes would still be present in its
genome [18, 77]. Using sequence homology searches, we found that Go. avonlea has one or more homologs to 212 of the 508 Gu. theta EGT genes (285 Go. avonlea proteins in total). Manual investigation of the phylogenies of each of these 285 proteins (Fig. 2) revealed that only six show any obvious red algal signal in both Cryptophyceae (including Gu. theta) and Go. avonlea,
none of which were predicted to be targeted to a plastid or function in
plastid metabolism. In contrast, the Cryptophyceae showed a significant
red-algal signal to the exclusion of Go. avonlea in 75 of these 285 phylogenies (e.g., tryptophanyl-tRNA synthetase; Fig. 3). Similar to the results of Curtis et al. [18],
a large proportion of these trees were found to be ambiguous with
respect to the nature of their algal signal. In some cases, the
cryptophyte homologs branch closest to green or glaucophyte algae
(31/285 and 97/285 trees where a Go. avonlea homolog branches with or without the predicted Gu. theta EGT in the phylogeny, respectively), while in others the primary algal lineage is entirely unclear (12/285 trees without a Go. avonlea homolog branching with the predicted cryptophyte EGT, 13/285 where a Go. avonlea
homolog branches with the predicted cryptophyte EGT). However, given
that the phylogenetic position of Cryptista relative to Archaeplastida
and other eukaryotic supergroups is unclear [14],
extreme caution is needed when considering these “green,” “glaucophyte”
or ambiguous algal genes as bona fide EGTs, particularly in cases where
obvious plastid targeting signals and/or plastid-associated functions
are not observed [18]. Here, plastid-targeting signals and/or plastid-associated functions were not observed for any Go. avonlea homolog that branched with a Gu. theta predicted EGT showing a common “green,” “glaucophyte,” or ambiguous algal phylogenetic signal.
Fig. 2
Algal genes in Guillardia theta and Goniomonas avonlea. The diagram shows the distribution of topologies observed in Go. avonlea homologs to 508 “algal” endosymbiotic gene transfers predicted by Curtis et al. [18] in Gu. theta. Phylogenies were evaluated and sorted based on the relative positioning of Go. avonlea and Gu. theta
(and other Cryptophyceae) and their relationship to Archaeplastida
lineages and secondarily photosynthetic taxa. An exclusive relationship
indicates a direct relationship with an Archaeplastida lineage while an
inclusive one indicates there are intervening secondarily photosynthetic
taxa. A given topological pattern was only assigned if the
corresponding UFboot support was greater than 80%. Of the 285 homologs
identified in Go. avonlea, only six show an affinity to Cryptophyceae and red algae
Fig. 3
Maximum
likelihood (ML) phylogeny of tryptophanyl-tRNA synthetase in diverse
eukaryotes and prokaryotes. The tree was inferred under the model LG4X
(with 100 standard bootstrap replicates) and shows an apparent red algal
ancestry for homologs in Cryptophyceae but not in Go. avonlea. Eukaryotic OTUs are colored according to their known or predicted “supergroup” affinities with sequences from Go. avonlea and predicted Gu. theta EGTs [17]
highlighted in bright red (Viridiplantae are in green, Glaucophyta are
in turquoise, Rhodophyta are in dark red, Cyanobacteria are orange and
other Bacteria are in gold, Cryptophyta are in pink, Haptophyta are in
purple, Stramenopiles are in dark blue, Alveolata are in blue, Rhizaria
are in light blue). The tree shown is midpoint rooted. Black dots
indicate maximal support for particular nodes. When not maximal, only
bootstrap support values > 70% are shown. The scale bar shows the
inferred number of amino acid substitutions per site
Is this small “red algal” footprint in the Go. avonlea genome (6/508 predicted algal EGTs in Gu. theta)
meaningful? Comparing this signal to that observed against a control
taxon (i.e., an unrelated amoebozoan with an unambiguous
non-photosynthetic ancestry) allowed us to assess the expected signal
due to background phylogenetic noise [77]. We found that Go. avonlea
appeared sister to Amoebozoa in 24/285 single-gene trees, substantially
higher than the red algal fraction. These analyses strongly suggest
that the common red-algal footprint in Go. avonlea and Gu. theta
is not significant, consistent with the lack of evidence for the
existence of a cryptic plastid from microscopy and protein subcellular
localization predictions. It is unclear to what extent our phylogenomic
results are biased by taxonomic sampling; genome sequence data are
presently stacked in favor of green algae and land plants over red algae
[15].
It will thus be interesting to see whether the number of “red-algal”
genes in Cryptophyceae (as well as other complex red algae-derived
plastid bearing taxa) and plastid-lacking lineages such as Go. avonlea will go up or down as databases become more inclusive.
Considering the Go. avonlea
predicted proteome as a whole, a top blast hit analysis revealed an
expected affinity to other Cryptista ~ 33% of the time, with the next
most common top hits being to Alveolata (~ 13%), Viridiplantae (~ 12%),
and Stramenopiles (~ 11%) (Fig. 4).
Notably, the number of instances in which an amoebozoan protein was the
most similar sequence (1263 proteins, 8%) was considerably greater than
those where a red algal homolog was most similar (128, 0.7%),
suggesting again that the red algal signal in the Go. avonlea genome is minimal and not the result of EGT. In the case of the phototroph Gu. theta, a ~ 4.4 times enrichment in red algal signal relative to Go. avonlea
was seen in a top blast hit analysis and a ~ 4.5 times greater
enrichment in terms of archaeplastidal signal (compared to an amoebozoan
control and adjusted for relative database representation to minimize
database composition bias [77]). There is thus no indication of a red algal signal above background noise in the Go. avonlea genome on the basis of top blast hits.
Fig. 4
Taxonomic distribution of top blast hits for Goniomonas avonlea proteins. The top blast hit was defined as the most significant homolog to Go. avonlea (i.e., lowest E-value with a cutoff of 1e−10) excluding any other Goniomonas sequence
The position of Cryptista on the eukaryotic tree of life
The
Cryptista comprises a diverse collection of heterotrophic and
photosynthetic lineages, one that has only recently been recognized as a
monophyletic entity [10, 78].
Not surprisingly, Cryptista has been difficult to place in the
eukaryotic tree of life; some or all of its members have been shown to
branch sister to Haptophyta (e.g., [8]) or, alternatively, sister to Archaeplastida (e.g., [13]). Phylogenies inferred here based on a modified Burki et al. [68] dataset (98 OTUs, 250 marker genes) recovered identical relationships to those inferred by Burki et al. [14];
however, we were able to evaluate branch support using standard
bootstrapping under the complex model LG + C60 + F + PMSF (Fig. 5) [69]. In Burki et al. [14],
Bayesian analyses on the original dataset did not result in global
convergence, an issue that is common when analyzing such large
phylogenomic datasets (e.g., [13]).
Nevertheless, these authors considered the tree topology resulting from
the non-converged Bayesian analysis and found only minor differences
with regard to the position of the Cryptista to Archaeplastida. Due to
the large size of our dataset, and in light of the observations of Burki
et al. [14],
we did not attempt Bayesian analyses; we instead focused on the ML
analysis whose tree topology could be statistically evaluated using
standard (i.e., nonparametric) bootstrapping. With the exception of
Archaeplastida, and the Excavata (whose monophyly is still debated;
e.g., see [79]),
the monophyly of eukaryotic supergroups (including SAR) was recovered
with maximum support, and Haptista (i.e., haptophytes + centrohelids)
branched with nearly maximum support as sister to SAR (99% standard
bootstrap support). The monophyly of Archaeplastida was disrupted by the
positioning of Cryptista, which was found to branch with Archaeplastida
with maximum support; more specifically, Cryptista branched with a
standard bootstrap value of 82% as sister to Viridiplantae and
Glaucophyta (99% standard bootstrap support) to the exclusion of
Rhodophyta.
Fig. 5
Phylogenomic
analysis of the eukaryotic tree of life. Tree shown is a maximum
likelihood (ML) phylogeny of a 250 marker gene/protein dataset as in
Burki et al. [14] that includes new transcriptome data from Go. avonlea.
The phylogeny is based on a concatenated marker gene alignment of
71,151 unambiguously aligned sites across 98 OTUs inferred under the
model LG + C60 + F + PMSF with 100 standard bootstrap replicates. The
tree shown is midpoint rooted. Black dots indicate maximal support for a
particular node. When not maximal, only bootstrap support values
> 70% are shown. The scale bar shows an inferred 0.2 substitutions
per site
Removing single genes in specific OTUs determined to be discordant via PhyloMCOA [71]
did not change the tree topology, but rather significantly increased
the support of Cryptista branching internal to Archaeplastida (90%
standard bootstrap support), suggesting that the observed relationship
is not caused by a few genes in Archaeplastida and Cryptista that
overwhelm the dataset with non-phylogenetic signal. The removal of
Cryptophyceae from the dataset also resulted in no change in tree
topology, recovering non-photosynthetic Cryptista as sister to
Viridiplantae and Glaucophyta to the exclusion of Rhodophyta with 75%
standard bootstrap support (not substantially different from Fig. 5),
suggesting that this association is not entirely due to the presence of
plastid-bearing lineages. It remains possible, however, that instances
of EGT have gone undetected within Cryptista due to the close
evolutionary relationship of their nuclear genes (either Viridiplantae
and Glaucophyta specifically or Archaeplastida as a whole) and the
source of their plastid (Rhodophyta), making it extremely difficult to
disentangle the source of genes in the nucleus and resolve the exact
position of the phylum Cryptista within eukaryotes [80].
Further
investigation into the position of Cryptista on the eukaryotic tree of
life using random subsets of marker genes resulted in Cryptista
branching consistently with some combination of one or more
Archaeplastida sub-groups (93/100 iterations) (Fig. 6).
While Cryptista was most frequently observed as sister to the clade
comprising Viridiplantae and Glaucophyta (30%), it was also often
recovered as sister to Glaucophyta exclusively (24%), Rhodophyta
exclusively (13%), and to a monophyletic Archaeplastida (20%).
Interestingly, in stark contrast to the 24% of iterations that resulted
in a Cryptista-Glaucophyta-specific relationship, only 3% showed
Cryptista branching with Viridiplantae exclusively. This may suggest
that Cryptista shares a closer ancestry with Glaucophyta, but it could
also simply be the result of similar compositional biases or slow
evolutionary rates causing “short branch exclusion” [81].
Notably, a sister relationship between Cryptista and
Haptophyta/Haptista was never observed. While the exact position of
Cryptista relative to Archaeplastida is uncertain, its association with
Archaeplastida appears stable. As discussed above, this relationship
makes it difficult to assign “algal genes” as EGTs in Cryptista, and it
remains to be determined if they are of endosymbiotic origin or vertical
ancestry.
Fig. 6
Impact of gene
sampling on the phylogenetic position of Cryptista on the tree of
eukaryotes. The diagram shows the phylogenetic position of Cryptista
within each ML tree inferred using randomly generated subsets of 250
marker genes from the Burki et al. [14]
dataset (four gene bins were used; for each iteration, three bins had
46 genes and one bin had 47 genes). Only marker genes for which a
homolog was present in Goniomonas avonlea
and at least one additional Cryptista were included. The distribution
shown is based on a total of 100 randomly generated marker gene subset
trees
Metabolic “rewiring” in Guillardia theta linked to plastid acquisition
Given that there is no plastid in Go. avonlea, we compared the predicted metabolic capacities of Gu. theta and Go. avonlea
with the goal of deducing the metabolic and enzymatic functions gained
with the acquisition of a red algal-derived secondary plastid. In our
study, four main biochemical pathways/processes are predicted to be
plastid-localized in Gu. theta
and thus obviously related to plastid acquisition: photosynthesis,
isoprenoid biosynthesis via the non-mevalonate (MEP/DOXP) pathway,
carotenoid biosynthesis, and porphyrin and chlorophyll metabolism
(Additional file 2:
Figure S4–S8). Several other pathways that may also have been acquired
by secondary endosymbiosis but are not obviously plastid-localized in Gu. theta are ubiquinone and terpenoid-quinone biosynthesis, as well as thiamine biosynthesis (Additional file 2: Figure S9 & S10). As expected, Gu. theta
pathways clearly localized to the plastid include those associated with
pigment biosynthesis and photosynthesis (carotenoid biosynthesis,
chlorophyll and porphyrin biosynthesis (Additional file 2: Figure S7 & S8)). The presence of a thiamine (vitamin B1) pathway (Additional file 2:
Figure S10), which does not appear to be plastid-localized, as well as
ubiquinone and menaquinone/phylloquinone biosynthesis, which are
involved in electron transport, also seems to correlate with secondary
plastid acquisition in Cryptophyceae. It should be noted that while
menaquinone biosynthesis should take place in the plastid, signal
peptides have not been detected on the requisite proteins in Gu. theta [18]. We also observed that while a peroxisome-localized primary bile biosynthesis pathway is present in Go. avonlea, it is apparently absent in Gu. theta (Additional file 2: Figure S11). This suggests either loss of this pathway in Gu. theta or later acquisition in Go. avonlea.
In Go. avonlea,
fatty acid biosynthesis is predicted to occur partly in the
mitochondrion (FabF and FabB) and partly in the cytosol (FAS1)
(Additional file 2: Figure S4, S12, S13); while in Gu. theta, it is predicted to be plastid-localized. Interestingly, while the mevalonate pathway in Go. avonlea is found in the cytosol (Additional file 2: Figure S4, S6), Gu. theta possesses both the mevalonate and MEP/DOXP pathways, which use acetyl-CoA and GA3P (d-glyceraldehyde-3-phosphate) with pyruvate, respectively, to synthesize isoprenoid precursor (Additional file 2: Figure S2 and S6). Gu. theta
(and perhaps other Cryptophyceae) thus appear to have redundant
metabolic capacities with which to synthesize isopentenyl diphosphate
(i.e., either the mevalonate or the MEP/DOXP pathway) which may
represent the ancestral eukaryotic metabolism or the endosymbiotically
derived one, respectively.
Storage polysaccharides in Goniomonas avonlea
Alpha
glucans are the most common storage polysaccharides and can be found in
different forms (e.g., glycogen and starch). Production of alpha
glucans can be assessed by the presence of certain CAZyme families:
glycoside hydrolase (GH)13, glycosyltransferase (GT)35 and GT5 for all
organisms, and GT3, GH133, and GT8 for eukaryotes in particular [82]. These enzymes are all found encoded in the Go. avonlea genome and are very similar to those involved in classical eukaryotic glycogen metabolism (Table 2).
Several proteins with GT8 domains can putatively be assigned as
glycogenins since their best blast hits are to bona fide glycogenins in
other organisms such as Saccharomyces cerevisiae,
albeit with poor E-values (data not shown). Additionally, a complete
metabolic pathway for the production of an alpha glucan storage
polysaccharide seems to be present in Go. avonlea,
as supported by the presence of catabolic enzymes such as GH13 and
GH14. Because of the presence of the carbohydrate-binding module 45
(CBM45) coupled to a pfam01326 domain (corresponding to a glucan water
dikinase (GWD); see below) (Table 2), Go. avonlea could be a starch accumulating organism, even though electronic microscopy has not revealed the presence of starch granules [7].
Table 2
CAZy family enzymes in Goniomonas avonlea and Guillardia theta (those predicted to be secreted are in parentheses)
In addition to genes associated with alpha glucan metabolism, the Go. avonlea genome encodes putative beta glucan-specific proteins, i.e., enzymes falling in the GT2 and GT48 families (Table 2).
These enzymes are implicated in either the production of cellulose in
the cell wall or the synthesis of beta storage polysaccharides [83]. No genes for GH9 enzymes were found in the Go. avonlea genome, consistent with the fact that cellulose has not been observed in goniomonads [76].
Even if we cannot exclude the possibility of the presence of glucan in
the periplast component of cryptomonads, we suggest that the presence of
GT2 and GT48 family enzymes could be related to the synthesis of beta
storage polysaccharides. The catabolism of beta polysaccharides in Go. avonlea
could be performed by GH16 family enzymes, laminarinase in particular.
However, the laminarinase-like enzymes appear to be secreted, suggesting
they are involved in the degradation of exogenous rather than
endogenous polysaccharides (Table 2).
Goniomonas avonlea appears capable of digesting both bacteria and eukaryotes
Many heterotrophic eukaryotes ingest bacteria by phagocytosis and Go. avonlea
is no exception. The CAZy database includes glycoside hydrolases (GHs)
clustered into 136 families, and our analysis of carbohydrate-active
enzymes (CAZymes) in Go. avonlea provides insight into what its prey might be. The Go. avonlea genome contains genes for three families of signal peptide-containing lysozymes (GH22, GH24, and GH25) (Table 2) that are likely associated with bacterial phagocytosis. The GH2 family in Go. avonlea also includes several enzymes with secretion signals (Table 2). Interestingly, the presence of several GH enzymes suggests that phagocytosis in Go. avonlea
may also involve eukaryotic prey, specifically algae: these are
proteins belonging to the GH45, GH5, and GH3 families, which are
putative cellulases, agarases (GH50), and putative hemicellulases (GH43
and GH54; Table 2).
Although the cellulases have signal peptides, suggesting that they are
involved in the degradation of algal cellulose, it should be noted that
cellulose is also found in some bacteria. More intriguing is the
identification of genes for signal peptide-containing agarases (GH50) in
the Go. avonlea nuclear genome, since agar is found only in red algae [84] (Table 2). This suggests that Go. avonlea
could feed on red algae by phagocytosis, although agarase is also known
to degrade alginate, which is found in some bacterial biofilms. The
presence of putative secreted hemicellulases in Go. avonlea is also consistent with the hypothesis that Go. avonlea
preys on algae. Several amylases (GH13) and two beta-amylases (GH14)
were found to have signal peptides, and may therefore be involved in the
degradation of storage polysaccharides from organisms taken up by
phagocytosis (Table 2).
While
plastid-bearing photosynthetic organisms fix carbon through the Calvin
cycle and transform it into sugars for various purposes (most notably,
energy), heterotrophic organisms need to acquire sugar from their
environment. Thus, photosynthetic organisms typically possess fewer GHs
than heterotrophic organisms. This general pattern holds when the
heterotroph Go. avonlea is compared to the phototroph Gu. theta. Go. avonlea possesses 183 GHs (111 of which are predicted to be secreted), compared to only 57 in Gu. theta (Table 2). Gu. theta also appears to lack certain GH families that are typically absent in autotrophs. Nevertheless, 18 of the 57 GHs in Gu. theta are predicted to be secreted, consistent with the gene-based model that predicts Gu. theta to be mixotrophic [85], as has been suggested for several other Cryptophyceae [86, 87, 88]. Another interesting observation is the co-occurrence of certain GH families in Gu. theta and Go. avonlea, most notably GH116. However, whereas Go. avonlea is predicted to secrete these enzymes in order to obtain exogenous polysaccharides, Gu. theta presumably uses them to digest its own endogenous polysaccharides (Table 2). Moreover, while Gu. theta does not have more GTs than Go. avonlea, some classes that are only present in Gu. theta (GT14, GT29) are involved in protein glycosylation (Table 2).
Global CAZome analysis
In order to better understand the biology of Go. avonlea relative to Gu. theta
and vice versa, we performed a global analysis of the CAZomes of both
organisms and compared them to those of other eukaryotes. On the basis
of similarity clustering (Additional file 2: Figure S14), we observed a close relationship between the GT families of Gu. theta and Go. avonlea. However, when the GH profiles are compared (Additional file 2: Figure S15), Go. avonlea is more similar to the rotifer Adineta vaga, an organism known to be able to degrade cellulose [89], and Gu. theta
is closer to metazoans. When the CAZome as a whole is analyzed, i.e.,
all of the carbohydrate-active enzymes predicted for each organism, the Go. avonlea profile is most similar to that of Gu. theta (Fig. 7)
and, together, these two cryptomonads are generally similar to other
organisms with secondarily derived plastids. To determine whether the
link between the CAZome profiles of Go. avonlea and plastid-bearing organisms is simply due to the presence of Gu. theta, we removed the latter organism and repeated the clustering analysis (Additional file 2: Figure S16). Even without Gu. theta, a specific relationship between the CAZomes of Go. avonlea and diverse algae (stramenopiles, Haptophyta, Rhizaria) is observed, suggesting that the carbohydrate-active enzyme profile of Go. avonlea is broadly similar to some algae (Additional file 2: Figure S16). In contrast, when Go. avonlea is removed from the analysis, Gu. theta was found to be closest to primary plastid-bearing algae, particularly Chlorophyta and Prasinophyta (Additional file 2:
Figure S17). At the present time, the broader significance of these
patterns is far from clear, but clearly the CAZomes—and carbohydrate
metabolisms—of Gu. theta and Go. avonlea are similar in some ways and different in others.
Fig. 7
Carbohydrate-active enzyme (CAZyme) families in the heterotroph Goniomonas avonlea, the phototroph Guillardia theta,
and other eukaryotes. Diagram shows a heatmap of CAZyme prevalence in
each taxon (abundance within a particular CAZy family divided by the
whole number of CAZy families predicted from the genome); the white to
blue color scheme indicates low to high prevalence, respectively.
Dendogram shows the relative proximity of taxa, or of the co-occurrence
of CAZyme families on the left. We observed a close relationship between
Gu. theta and Go. avonlea (salmon), and that Cryptophyta have a set of CAZyme families similar to that seen in other secondarily photosynthetic algae
From heterotroph to phototroph: the complexity of cryptomonads
Subsequent
to the evolution of primary plastids in Archaeplastida, a wide range of
eukaryotes acquired photosynthesis secondarily via the engulfment of a
red or green algal endosymbiont [2, 4].
Despite more than a decade of cell biological, biochemical,
phylogenetic, and phylogenomic investigation, it is still not clear how
many times this occurred during the evolution of eukaryotes [6, 90, 91].
On the eukaryotic tree of life, secondary plastid-bearing organisms are
scattered amongst plastid-lacking ones, and whether plastid gain or
plastid loss has been the dominant mode of organelle evolution has
proven difficult to discern.
In
the case of red algal-derived plastids, recent evidence strongly
suggests a single evolutionary origin of the plastid protein import
machinery operating in cryptophytes, haptophytes, photosynthetic
stramenopiles, and many alveolates: these organisms all use a
host-derived multi-protein complex called SELMA (symbiont-specific
endoplasmic reticulum-associated degradation-like machinery) for protein
translocation across the second outermost plastid membrane of their
four membrane-bound plastids [92, 93, 94]
(the three membrane-bound plastids of dinoflagellates are an
exception). This could mean that the plastids in each of these lineages
stem from a single, ancient secondary endosymbiosis involving a red alga
and a heterotrophic host (the so-called “chromalveolate” hypothesis [91]),
followed by extensive plastid loss in their heterotrophic relatives.
Alternatively, one or more cryptic tertiary endosymbioses could have
spread the original secondary red plastid (and the genes for SELMA
proteins) across the eukaryotic tree (see, e.g., [80, 95, 96]).
Distinguishing between these two scenarios is fraught with challenges,
not least of which is the fact that the deep structure of the eukaryotic
tree of life continues to evolve (see above; Fig. 5) and the biology of many heterotrophic protist lineages remains poorly described.
Using genomic and transcriptomic data, we have explored the metabolic capacities of Go. avonlea—the
first member of the heterotrophic Goniomonadea to have its genome
sequenced—and compared them to those of the model plastid-bearing
cryptophyte Gu. theta. We have shown that in Gu. theta,
endosymbiosis led to the gain of metabolic pathways/processes
presumably already present in the host, such as fatty acid biosynthesis,
as well as de novo acquisition of photosynthesis and the MEP/DOXP
pathway (Additional file 2:
Figure S5 and S6). Despite its apparent redundancy with the cytosolic
mevalonate (MVA) pathway, the plastid-localized MEP/DOXP pathway in Gu. theta
and other cryptophytes is presumably “useful” given that its end
product feeds into other metabolic pathways. For example, the MEP/DOXP
pathway generates phytyl-PP, which is used to produce several important
components for photosynthesis such as phylloquinone, tocopherol, and
chlorophyll [97, 98, 99].
Some organisms use the plastid MEP/DOXP pathway instead of the
cytosolic MVA pathway, e.g., chlorophyte algae and certain
dinoflagellates [100]. In Gu. theta,
the benefits of photosynthesis presumably offset the costs associated
with partial or complete metabolic redundancies. Nevertheless, Gu. theta
does appear to have lost certain metabolic capacities that were present
in its heterotrophic ancestors, such as primary bile biosynthesis
(Additional file 2: Figure S4 and S11).
Glycosyl
transferases (GTs) catalyze the transfer of sugars from donor to
acceptor molecules, thereby creating glycosidic bonds. The evolution of
photosynthesis is generally associated with the acquisition of new GT
families and this is indeed what seems to have happened in the
cryptophytes. Interestingly, however, we did not observe a significant
increase in the total number of genes for GT enzymes in Gu. theta relative to Go. avonlea, and in fact, some GTs are present in Go. avonlea but not in Gu. theta; some GTs (GT3, GT48, GT34) appear to have been lost in Gu. theta and others replaced (e.g., GT3 has probably been replaced by a GT5 in Gu. theta) [82]. In addition, the loss of GT48 seems to correlate with the loss of beta glucan synthesis, as Gu. theta appears capable of generating only alpha-glucan polysaccharides, as in the red algal progenitor of its plastid.
Most unexpected was the discovery of a sequence of the GT28 family in Go. avonlea (Table 2),
a category of enzymes known to be involved in the synthesis of cell
wall components in bacteria (MurG is a GT28 enzyme acting as a UDP-N-acetylglucosamine:lipopolysaccharide N-acetylglucosamine
transferase) and the synthesis of galactoglycerolipids in
plastid-bearing organisms (MGD is a GT28 enzyme synthesizing
1,2-Diacyl-3-beta-d-galactosyl-sn-glycerol). All GT28-containing eukaryotes in the public CAZy database have plastids (http://www.cazy.org/GT28_eukaryota.html),
and more recent investigation shows that of the more than 1000
eukaryotes that have had their CAZomes annotated, only eight have GT28
genes and lack plastids (most of these are fungi; data not shown). The
GT28 gene in Go. avonlea resides on a contig with six other genes, five of which have a top blast hit to another eukaryote (2 being Gu. theta), and the sixth is a bacterial-like gene that also has a close homolog in the Gu. theta nuclear genome (Additional file 2: Figure S18). In addition, the GT28 gene in Go. avonlea contains spliceosomal introns, confirming its provenance as a eukaryotic nuclear gene. The exact function of the Go. avonlea
GT28 enzyme is difficult to predict with confidence. Based on sequence
similarity, it could be a 1,2-diacylglycerol
3-beta-galactosyltransferase [EC 2.4.1.46] and is predicted to be
targeted to the mitochondrion by TargetP and PredSL; this is also
thought to be the case in some non-photosynthetic algae [101]. We speculate that the Go. avonlea
enzyme might be involved in the synthesis of phosphate-free
mitochondrial lipids, which could serve in a
phospholipid-to-galactolipid exchange as observed in Arabidopsis thaliana mitochondria during phosphate starvation [102]. The origin and function of the GT28 gene in Go. avonlea
thus appears distinct from the MGD genes of plastid-bearing eukaryotes.
Biochemical, physiological, and ideally functional genomic studies need
to be undergone to solve this question. Regardless of its precise
function, it is noteworthy that in phylogenetic analyses, the GT28
homologs of Go. avonlea and Go. pacifica branch robustly with cryptophytes (including Gu. theta), red algae, and haptophytes (Fig. 8).
Fig. 8
Phylogenetic
analysis of glycosyltransferase (GT) 28. The tree shown is a maximum
likelihood tree with ultrafast bootstrap values mapped onto the nodes.
The tree shown is midpoint rooted. Sequences are colored according to
their taxonomic affiliation: Viridiplantae are in green, Glaucophyta are
in turquoise, Rhodophyta are in dark red, Cyanobacteria are orange and
other Bacteria are in gold, Cryptophyta are in pink, Goniomonas avonlea
is dark red and bolded, Haptophyta are in purple, Stramenopiles are in
dark blue, Alveolata are in blue, Rhizaria are in light blue. The GT28
from Go. avonlea groups with other cryptomonads and with Rhodophyta. It is noteworthy that GT28 grouping with Go. avonlea
seem to be mitochondrial based on signal targeting prediction while
GT28 on the upper part could be targeted to the plastid, based on
targeting prediction signal. The scale bar shows the inferred number of
amino acid substitutions per site
Another unexpected carbohydrate-active enzyme in Go. avonlea is glucan water dikinase (GWD). The discovery of GWD in a goniomonad is unexpected because it has long been assumed that Gu. theta
and other cryptophytes acquired starch metabolism as a result of the
acquisition of its secondary plastid. GWD has thus far only been found
in organisms known to accumulate starch; the enzyme has been proposed to
have evolved concomitantly with the primary plastid found in
Viridiplantae, Rhodophyta, and Glaucophyta (i.e., Archaeplastida) [103].
Indeed, all eukaryotic starch accumulators either have a plastid or
have been proposed to have once had a plastid—such as ciliates [104]—at some point during their evolution [82, 103] (Fig. 9). In Go. avonlea,
the GWD gene contains introns and resides on a contig with seven other
genes, three of which are clearly of eukaryotic provenance
(Additional file 2: Figure S19). Given the absence of obvious starch granules in Go. avonlea,
it is possible that the organism only synthesizes starch at certain
stages of its life cycle, as is the case in the apicomplexan Toxoplasma gondii [105]. In phylogenetic analyses, the Go. avonlea
GWD homolog does not branch with cryptophytes, but rather is weakly
associated with a clade containing sequences from Rhodophyta,
Glaucophyta, and various algae with red algal type plastids. How Go. avonlea came to possess and retain its GWD gene is an open question.
Fig. 9
Glucan Water
Dikinase (GWD) phylogenetic tree. The tree shown is a maximum likelihood
tree with ultrafast bootstrap values mapped onto the nodes. The tree is
rooted with the Phosphoglucan dikinase (PWD) sequences. Sequences are
colored according to their taxonomic affiliation: Viridiplantae are in
green, Glaucophyta are in turquoise, Rhodophyta are in dark red,
Cyanobacteria are orange and other Bacteria are in gold, Cryptophyta are
in pink, Goniomonas avonlea is dark
red and bolded, Haptophyta are in purple, Stramenopiles are in dark
blue, Alveolata are in blue, Rhizaria are in light blue. Some bacteria
(in gold) could have obtained their GWD gene by LGT. The GWD homolog
from Go. avonlea branches close to
its counterpart in Rhodophyta and Glaucophyta; GWDs in Cryptophyta
appear more distantly related. The scale bar shows the inferred number
of amino acid substitutions per site
We have sequenced the nuclear genome of Goniomonas avonlea, the first of its kind for a plastid-lacking cryptomonad and only the second to be sequenced for all of Cryptista. The Go. avonlea
genome provides a much-needed first glimpse into the biology of a
heterotrophic protist residing on a large, poorly understood branch of
the eukaryotic tree. Amongst the 33,470 predicted protein-coding genes
in the Go. avonlea genome
are hundreds of genes for carbohydrate-active enzymes that provide
important clues as to what this phagotrophic protist eats in nature—the
organism appears capable of digesting bacteria as well as eukaryotes,
including algae. We found no convincing phylogenetic evidence to support
the notion that Go. avonlea
evolved from a secondary plastid-bearing ancestor; in terms of
abundance, the handful of “algal” genes in the genome do not rise above
background “noise.” Nevertheless, this enigmatic protist possesses genes
for enzymes such as GT28 and GWD, which are almost invariably found in
plastid-bearing organisms. This is interesting for various reasons, not
least of which is the fact that analysis of a 250-protein dataset placed
cryptophytes, goniomonads (including Go. avonlea), and other heterotrophic Cryptista within the primary plastid-bearing Archaeplastida. While Go. avonlea
serves as an important reference point for studying the metabolic
transformation that took place during secondary endosymbiosis in the
ancestor of modern-day Cryptophyceae, aspects of biochemistry and
molecular biology may be linked to its deep ties with primary
plastid-bearing organisms. More genomic data from diverse heterotrophic
members of the Cryptista will hopefully allow us to test this
hypothesis.
We thank two anonymous reviewers for helpful comments on an earlier version of this manuscript.
Funding
The Archibald Lab
acknowledges support in the form of a Discovery grant (RGPIN-2014-05871)
from the Natural Sciences and Engineering Research Council of Canada.
EK is the recipient of a Simons Foundation award (SF-382790). EM is
supported by the French National Research Agency (ANR-10-LABEX-04 GRAL
Labex, Grenoble Alliance for Integrated Structural Cell Biology;
ANR-11-BTBR-0008 Océanomics). SJS was supported by a NSERC CGS-M
graduate scholarship and a Nova Scotia Graduate Scholarship.
Availability of data and materials
The genomic and
transcriptomic datasets supporting the conclusions of this article are
available in GenBank under accession numbers QUTJ00000000 and
GGUN00000000, respectively, while the Go. avonlea mitochondrial genome is available under the following accession number: AP018919.
Authors’ contributions
EK
and JMA designed the study. UC, SJS, BAC, RK, LE, DM, BH, EM, MC, and
CD collected data and carried out analyses; all authors contributed to
data interpretation; and UC, SJS, BAC, EK, and JMA wrote and revised the
manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open AccessThis article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/),
which permits unrestricted use, distribution, and reproduction in any
medium, provided you give appropriate credit to the original author(s)
and the source, provide a link to the Creative Commons license, and
indicate if changes were made. The Creative Commons Public Domain
Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
Moreira D, Le Guyader H, Philippe H. The origin of red algae and the evolution of chloroplasts. Nature. 2000;405(6782):69–72.PubMedGoogle Scholar
Keeling PJ. The endosymbiotic origin, diversification and fate of plastids. Philos Trans R Soc B Biol Sci. 2010;365:729–48.Google Scholar
Adl
SM, Simpson AGB, Lane CE, Lukeš J, Bass D, Bowser SS, et al. The
revised classification of eukaryotes. J Eukaryot Microbiol.
2012;59:429–514.PubMedPubMed CentralGoogle Scholar
Archibald JM. The puzzle of plastid evolution. Curr Biol. 2009;19:R81–8.PubMedGoogle Scholar
Kim E, Archibald JM. Ultrastructure and molecular phylogeny of the cryptomonad Goniomonas avonlea sp. nov. Protist. 2013;164:160–82.PubMedGoogle Scholar
Okamoto N, Chantangsi C, Horák A, Leander BS, Keeling PJ. Molecular phylogeny and description of the novel katablepharid Roombia truncata gen. Et sp. nov., and establishment of the Hacrobia taxon nov. PLoS One. 2009;4:e7080.PubMedPubMed CentralGoogle Scholar
Yabuki A, Kamikawa R, Ishikawa SA, Kolisko M, Kim E, Tanabe AS, et al. Palpitomonas bilix represents a basal cryptist lineage: insight into the character evolution in Cryptista. Sci Rep. 2014;4. https://doi.org/10.1038/srep04641.
Patron
NJ, Inagaki Y, Keeling PJ. Multiple gene phylogenies support the
monophyly of cryptomonad and haptophyte host lineages. Curr Biol.
2007;17:887–91.PubMedGoogle Scholar
Burki
F, Inagaki Y, Bråte J, Archibald JM, Keeling PJ, Cavalier-Smith T, et
al. Large-scale phylogenomic analyses reveal that two enigmatic protist
lineages, telonemia and centroheliozoa, are related to photosynthetic
chromalveolates. Genome Biol Evol. 2009;1:231–8.PubMedPubMed CentralGoogle Scholar
Katz
LA, Grant JR. Taxon-rich phylogenomic analyses resolve the eukaryotic
tree of life and reveal the power of subsampling by sites. Syst Biol.
2015;64:406–15.PubMedGoogle Scholar
Burki
F, Kaplan M, Tikhonenkov DV, Zlatogursky V, Minh BQ, Radaykina LV, et
al. Untangling the early diversification of eukaryotes: a phylogenomic
study of the evolutionary origins of Centrohelida, Haptophyta and
Cryptista. Proc R Soc B Biol Sci. 2016;283:20152802.Google Scholar
Sibbald SJ, Archibald JM. More protist genomes needed. Nat Ecol Evol. 2017;1:0145.Google Scholar
Archibald JM. Nucleomorph genomes: structure, function, origin and evolution. BioEssays. 2007;29:392–402.PubMedGoogle Scholar
Moore CE, Curtis B, Mills T, Tanifuji G, Archibald JM. Nucleomorph genome sequence of the cryptophyte alga Chroomonas mesostigmatica CCMP1168 reveals lineage-specific gene loss and genome complexity. Genome Biol Evol. 2012;4:1162–75.PubMedPubMed CentralGoogle Scholar
Curtis
BA, Tanifuji G, Burki F, Gruber A, Irimia M, Maruyama S, et al. Algal
genomes reveal evolutionary mosaicism and the fate of nucleomorphs.
Nature. 2012;492:59–65.PubMedGoogle Scholar
Rolland
N, Curien G, Finazzi G, Kuntz M, Maréchal E, Matringe M, et al. The
biosynthetic capacities of the plastids and integration between
cytoplasmic and chloroplast processes. Annu Rev Genet. 2012;46:233–64.Google Scholar
Pfannschmidt
T, Schutze K, Brost M, Oelmuller R. A novel mechanism of nuclear
photosynthesis gene regulation by redox signals from the chloroplast
during photosystem stoichiometry adjustment. J Biol Chem.
2001;276:36125–30.PubMedGoogle Scholar
Maruyama
S, Suzaki T, Weber AP, Archibald JM, Nozaki H. Eukaryote-to-eukaryote
gene transfer gives rise to genome mosaicism in euglenids. BMC Evol
Biol. 2011;11:105.PubMedPubMed CentralGoogle Scholar
Lombard
V, Golaconda Ramulu H, Drula E, Coutinho PM, Henrissat B. The
carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res.
2014;42:D490–5.PubMedGoogle Scholar
Okaichi
T, Nishio S, Imatomi Y. Collection and mass culture. In: Yûdoku
Purankuton-Hassei, Sayôkikô, Dokuseibun, editors. Toxic phytoplankton -
Occurence, mode of action and toxins. Tokyo:Jpn Fish Soc. 1982;22–34.Google Scholar
Bolger AM, Lohse M, Usadel B. Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics. 2014;30:2114–20.PubMedPubMed CentralGoogle Scholar
Grabherr
MG, Haas BJ, Yassour M, Levin JZ, Thompson DA, Amit I, et al.
Full-length transcriptome assembly from RNA-Seq data without a reference
genome. Nat Biotechnol. 2011;29:644–52.PubMedPubMed CentralGoogle Scholar
Butler
J, MacCallum I, Kleber M, Shlyakhter IA, Belmonte MK, Lander ES, et al.
ALLPATHS: De novo assembly of whole-genome shotgun microreads. Genome
Res. 2008;18:810–20.PubMedPubMed CentralGoogle Scholar
Simpson
JT, Wong K, Jackman SD, Schein JE, Jones SJM, Birol I. ABySS: a
parallel assembler for short read sequence data. Genome Res.
2009;19:1117–23.PubMedPubMed CentralGoogle Scholar
Sommer DD, Delcher AL, Salzberg SL, Pop M. Minimus: a fast, lightweight genome assembler. BMC Bioinformatics. 2007;8:64.PubMedPubMed CentralGoogle Scholar
Boisvert
S, Laviolette F, Corbeil J. Ray: simultaneous assembly of reads from a
mix of high-throughput sequencing technologies. J Comput Biol.
2010;17:1519–33.PubMedPubMed CentralGoogle Scholar
Langmead B, Salzberg SL. Fast gapped-read alignment with bowtie 2. Nat Meth. 2012;9:357–9.Google Scholar
Clark
SC, Egan R, Frazier PI, Wang Z. ALE: a generic assembly likelihood
evaluation framework for assessing the accuracy of genome and metagenome
assemblies. Bioinformatics. 2013;29:435–43.PubMedGoogle Scholar
Altschul
SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, et al. Gapped
BLAST and PSI-BLAST: a new generation of protein database search
programs. Nucleic Acids Res. 1997;25:3389–402.PubMedPubMed CentralGoogle Scholar
Stanke
M, Diekhans M, Baertsch R, Haussler D. Using native and syntenically
mapped cDNA alignments to improve de novo gene finding. Bioinformatics.
2008;24:637–44.PubMedGoogle Scholar
Stanke
M, Schöffmann O, Morgenstern B, Waack S. Gene prediction in eukaryotes
with a generalized hidden Markov model that uses hints from external
sources. BMC Bioinformatics. 2006;7:62.PubMedPubMed CentralGoogle Scholar
Haas BJ. Improving the Arabidopsis genome annotation using maximal transcript alignment assemblies. Nucleic Acids Res. 2003;31:5654–66.PubMedPubMed CentralGoogle Scholar
Keeling
P, Burki F, Wilcox J, Allam B, et al. The marine microbial eukaryote
transcriptome sequencing project (MMETSP): illuminating the functional
diversity of eukaryotic life in the oceans through transcriptome
sequencing. PLoS Biol. 2014;12:e1001889.PubMedPubMed CentralGoogle Scholar
Wang
Y, Coleman-Derr D, Chen G, Gu YQ. OrthoVenn: a web server for genome
wide comparison and annotation of orthologous clusters across multiple
species. Nucleic Acids Res. 2015;43:W78–84.PubMedPubMed CentralGoogle Scholar
Wu
S, Zhu Z, Fu L, Niu B, Li W. WebMGA: a customizable web server for fast
metagenomic sequence analysis. BMC Genomics. 2011;12:444.PubMedPubMed CentralGoogle Scholar
Mao
X, Cai T, Olyarchuk JG, Wei L. Automated genome annotation and pathway
identification using the KEGG Orthology (KO) as a controlled vocabulary.
Bioinformatics. 2005;21:3787–93.PubMedGoogle Scholar
Kanehisa
M, Sato Y, Morishima K. BlastKOALA and GhostKOALA: KEGG tools for
functional characterization of genome and metagenome sequences. J Mol
Biol. 2016;428:726–31.Google Scholar
Kanehisa
M, Goto S, Sato Y, Furumichi M, Tanabe M. KEGG for integration and
interpretation of large-scale molecular data sets. Nucleic Acids Res.
2012;40:D109–14.PubMedGoogle Scholar
Petsalaki
EI, Bagos PG, Litou ZL, Hamodrakas SJ. PredSL: a tool for the
N-terminal sequence-based prediction protein subcellular localization.
Genomics Proteomics Bioinformatics. 2006;4:48–55.PubMedPubMed CentralGoogle Scholar
Emanuelsson
O, Brunak S, von Heijne G, Nielsen H. Locating proteins in the cell
using TargetP, SignalP and related tools. Nat Protoc. 2007;2:953–71.PubMedGoogle Scholar
Small
I, Peeters N, Legeai F, Lurin C. Predotar: a tool for rapidly screening
proteomes forN-terminal targeting sequences. Proteomics.
2004;4:1581–90.PubMedGoogle Scholar
Petersen
TN, Brunak S, von Heijne G, Nielsen H. SignalP 4.0: discriminating
signal peptides from transmembrane regions. Nat Methods. 2011;8:785–6.PubMedPubMed CentralGoogle Scholar
Gruber
A, Rocap G, Kroth PG, Armbrust EV, Mock T. Plastid proteome prediction
for diatoms and other algae with secondary plastids of the red lineage.
Plant J. 2015;81:519–28.PubMedPubMed CentralGoogle Scholar
Mistry
J, Finn RD, Eddy SR, Bateman A, Punta M. Challenges in homology search:
HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids
Res. 2013;41:e121.PubMedPubMed CentralGoogle Scholar
Bray JR, Curtis JT. An ordination of the upland forest communities of southern Wisconsin. Ecol Monogr. 1957;27:325–49.Google Scholar
Ward JH. Hierarchical grouping to optimize an objective function. J Am Stat Assoc. 1963;58:236.Google Scholar
Soverini
M, Turroni S, Biagi E, Quercia S, Brigidi P, Candela M, et al.
Variation of carbohydrate-active enzyme patterns in the gut microbiota
of Italian healthy subjects and type 2 diabetes patients. Front
Microbiol. 2017;8. https://doi.org/10.3389/fmicb.2017.02079.
Buchfink B, Xie C, Huson DH. Fast and sensitive protein alignment using DIAMOND. Nat Methods. 2015;12:59–60.PubMedGoogle Scholar
Finn
RD, Attwood TK, Babbitt PC, Bateman A, Bork P, Bridge AJ, et al.
InterPro in 2017—beyond protein family and domain annotations. Nucleic
Acids Res. 2017;45:D190–9.PubMedGoogle Scholar
Katoh
K, Standley DM. MAFFT multiple sequence alignment software version 7:
improvements in performance and usability. Mol Biol Evol.
2013;30:772–80.PubMedPubMed CentralGoogle Scholar
Criscuolo
A, Gribaldo S. BMGE (block mapping and gathering with entropy): a new
software for selection of phylogenetic informative regions from multiple
sequence alignments. BMC Evol Biol. 2010;10:210.PubMedPubMed CentralGoogle Scholar
Price
MN, Dehal PS, Arkin AP. Others. FastTree 2–approximately
maximum-likelihood trees for large alignments. PLoS One. 2010;5:e9490.PubMedPubMed CentralGoogle Scholar
Nguyen
L-T, Schmidt HA, von Haeseler A, Minh BQ. IQ-TREE: a fast and effective
stochastic algorithm for estimating maximum-likelihood phylogenies. Mol
Biol Evol. 2015;32:268–74.Google Scholar
Le
SQ, Dang CC, Gascuel O. Modeling protein evolution with several amino
acid replacement matrices depending on site rates. Mol Biol Evol.
2012;29:2921–36.PubMedGoogle Scholar
Minh BQ, Nguyen MAT, von Haeseler A. Ultrafast approximation for phylogenetic bootstrap. Mol Biol Evol. 2013;30:1188–95.PubMedPubMed CentralGoogle Scholar
Cenci
U, Moog D, Curtis BA, Tanifuji G, Eme L, Lukeš J, et al. Heme pathway
evolution in kinetoplastid protists. BMC Evol Biol. 2016;16. https://doi.org/10.1186/s12862-016-0664-6.
Cenci
U, Ducatez M, Kadouche D, Colleoni C, Ball SG. Was the chlamydial
adaptative strategy to tryptophan starvation an early determinant of
plastid endosymbiosis? Front Cell Infect Microbiol. 2016;6. https://doi.org/10.3389/fcimb.2016.00067.
Maruyama S, Eveleigh RJ, Archibald JM. Treetrimmer: a method for phylogenetic dataset size reduction. BMC Res Notes. 2013;6:145.PubMedPubMed CentralGoogle Scholar
Edgar RC. MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004;32:1792–7.PubMedPubMed CentralGoogle Scholar
Burki
F, Kaplan M, Tikhonekov DV, Zlatogursky V, Minh BQ, Radaykina LV, et
al. Data from: Untangling the early diversification of eukaryotes: a
phylogenomic study of the evolutionary origins of Centrohelida,
Haptophyta, and Cryptista. 2016. doi:https://doi.org/10.5061/dryad.rj87v.
Wang
H-C, Minh BQ, Susko E, Roger AJ. Modeling site heterogeneity with
posterior mean site frequency profiles accelerates accurate phylogenomic
estimation. Syst Biol. 2018;67:216–35.PubMedGoogle Scholar
Kalyaanamoorthy
S, Minh B, Wong T, von Haeseler A, Jermiin L. ModelFinder: fast model
selection for accurate phylogenetic estimates. Nat Methods.
2017;14:587–9.PubMedPubMed CentralGoogle Scholar
de
Vienne DM, Ollier S, Aguileta G. Phylo-MCOA: a fast and efficient
method to detect outlier genes and species in phylogenomics using
multiple co-inertia analysis. Mol Biol Evol. 2012;29:1587–98.PubMedGoogle Scholar
Kim E, Lane CE, Curtis BA, Kozera C, Bowman S, Archibald JM. Complete sequence and analysis of the mitochondrial genome of Hemiselmis andersenii CCMP644 (Cryptophyceae). BMC Genomics. 2008;9:215.PubMedPubMed CentralGoogle Scholar
Kim
JI, Yoon HS, Yi G, Shin W, Archibald JM. Comparative mitochondrial
genomics of cryptophyte algae: gene shuffling and dynamic mobile genetic
elements. BMC Genomics. 2018;19. https://doi.org/10.1186/s12864-018-4626-9.
Nishimura Y, Tanifuji G, Kamikawa R, Yabuki A, Hashimoto T, Inagaki Y. Mitochondrial genome of Palpitomonas bilix : derived genome structure and ancestral system for cytochrome c maturation. Genome Biol Evol. 2016;8:3090–8.PubMedPubMed CentralGoogle Scholar
McFadden GI, Gilson PR, Hill DRA. Goniomonas:
rRNA sequences indicate that this phagotrophic flagellate is a close
relative of the host component of cryptomonads. Eur J Phycol.
1994;29:29–32.Google Scholar
Martin-Cereceda
M, Roberts EC, Wootton EC, Bonaccorso E, Dyal P, Guinea A, et al.
Morphology, ultrastructure, and small subunit rDNA phylogeny of the
marine heterotrophic flagellate Goniomonas aff. amphinema. J Eukaryot Microbiol. 2010;57:159–70.PubMedGoogle Scholar
Stiller
JW, Huang J, Ding Q, Tian J, Goodwillie C. Are algal genes in
nonphotosynthetic protists evidence of historical plastid endosymbioses?
BMC Genomics. 2009;10:484.PubMedPubMed CentralGoogle Scholar
Cavalier-Smith
T. Symbiogenesis: mechanisms, evolutionary consequences, and systematic
implications. Annu Rev Ecol Evol Syst. 2013;44:145–72.Google Scholar
Heiss
AA, Kolisko M, Ekelund F, Brown MW, Roger AJ, Simpson AGB. Combined
morphological and phylogenomic re-examination of malawimonads, a
critical taxon for inferring the evolutionary history of eukaryotes. R
Soc Open Sci. 2018;5:171707.PubMedPubMed CentralGoogle Scholar
Archibald JM. Endosymbiosis and eukaryotic cell evolution. Curr Biol. 2015;25:R911–21.PubMedGoogle Scholar
Burki
F, Okamoto N, Pombert J-F, Keeling PJ. The evolutionary history of
haptophytes and cryptophytes: phylogenomic evidence for separate
origins. Proc R Soc B Biol Sci. 2012;279:2246–54.Google Scholar
Ball
S, Colleoni C, Cenci U, Raj JN, Tirtiaux C. The evolution of glycogen
and starch metabolism in eukaryotes gives molecular clues to understand
the establishment of plastid endosymbiosis. J Exp Bot. 2011;62:1775–801.PubMedGoogle Scholar
O’Neill EC, Trick M, Hill L, Rejzek M, Dusi RG, Hamilton CJ, et al. The transcriptome of Euglena gracilis reveals unexpected metabolic capabilities for carbohydrate and natural product biochemistry. Mol BioSyst. 2015;11:2808–20.PubMedGoogle Scholar
Popper
ZA, Michel G, Hervé C, Domozych DS, Willats WGT, Tuohy MG, et al.
Evolution and diversity of plant cell walls: from algae to flowering
plants. Annu Rev Plant Biol. 2011;62:567–90.PubMedGoogle Scholar
Burns
JA, Pittis AA, Kim E. Gene-based predictive models of trophic modes
suggest Asgard archaea are not phagocytotic. Nat Ecol Evol.
2018;2:697–704.PubMedGoogle Scholar
McKie-Krisberg
ZM, Gast RJ, Sanders RW. Physiological responses of three species of
antarctic mixotrophic phytoflagellates to changes in light and dissolved
nutrients. Microb Ecol. 2015;70:21–9.PubMedGoogle Scholar
Selosse M-A, Charpin M, Not F. Mixotrophy everywhere on land and in water: the grand écart hypothesis. Ecol Lett. 2017;20:246–63.PubMedGoogle Scholar
Tranvik LJ, Porter KG, Sieburth JM. Occurrence of bacterivory in Cryptomonas, a common freshwater phytoplankter. Oecologia. 1989;78:473–6.PubMedGoogle Scholar
Flot
J-F, Hespeels B, Li X, Noel B, Arkhipova I, Danchin EGJ, et al. Genomic
evidence for ameiotic evolution in the bdelloid rotifer Adineta vaga. Nature. 2013;500:453–7.PubMedGoogle Scholar
Zimorski V, Ku C, Martin WF, Gould SB. Endosymbiotic theory for organelle origins. Curr Opin Microbiol. 2014;22:38–48.PubMedGoogle Scholar
Cavalier-Smith
T. Principles of protein and lipid targeting in secondary
symbiogenesis: euglenoid, dinoflagellate, and sporozoan plastid origins
and the eukaryote family tree. J Eukaryot Microbiol. 1999;46:347–66.PubMedGoogle Scholar
Hempel
F, Bullmann L, Lau J, Zauner S, Maier UG. ERAD-derived preprotein
transport across the second outermost plastid membrane of diatoms. Mol
Biol Evol. 2009;26:1781–90.PubMedGoogle Scholar
Sommer
MS, Gould SB, Lehmann P, Gruber A, Przyborski JM, Maier U-G.
Der1-mediated preprotein import into the periplastid compartment of
chromalveolates? Mol Biol Evol. 2007;24:918–28.PubMedGoogle Scholar
Stork
S, Moog D, Przyborski JM, Wilhelmi I, Zauner S, Maier UG. Distribution
of the SELMA translocon in secondary plastids of red algal origin and
predicted uncoupling of ubiquitin-dependent translocation from
degradation. Eukaryot Cell. 2012;11:1472–81.PubMedPubMed CentralGoogle Scholar
Stiller
JW, Schreiber J, Yue J, Guo H, Ding Q, Huang J. The evolution of
photosynthesis in chromist algae through serial endosymbioses. Nat
Commun. 2014;5:5764.PubMedPubMed CentralGoogle Scholar
Baurain
D, Brinkmann H, Petersen J, Rodriguez-Ezpeleta N, Stechmann A, Demoulin
V, et al. Phylogenomic evidence for separate acquisition of plastids in
cryptophytes, haptophytes, and stramenopiles. Mol Biol Evol.
2010;27:1698–709.PubMedGoogle Scholar
Gross
J, Cho WK, Lezhneva L, Falk J, Krupinska K, Shinozaki K, et al. A plant
locus essential for phylloquinone (vitamin K1) biosynthesis originated
from a fusion of four eubacterial genes. J Biol Chem. 2006;281:17189–96.PubMedGoogle Scholar
Emonds-Alt
B, Coosemans N, Gerards T, Remacle C, Cardol P. Isolation and
characterization of mutants corresponding to the MENA, MENB, MENC and
MENE enzymatic steps of 5′-monohydroxyphylloquinone biosynthesis in Chlamydomonas reinhardtii. Plant J. 2016. https://doi.org/10.1111/tpj.13352.PubMedPubMed CentralGoogle Scholar
Chatterjee A, Kundu S. Revisiting the chlorophyll biosynthesis pathway using genome scale metabolic model of Oryza sativa japonica. Sci Rep. 2015;5:14975.PubMedPubMed CentralGoogle Scholar
Bentlage
B, Rogers TS, Bachvaroff TR, Delwiche CF. Complex ancestries of
isoprenoid synthesis in dinoflagellates. J Eukaryot Microbiol.
2016;63:123–37.PubMedGoogle Scholar
Borza T, Popescu CE, Lee RW. Multiple metabolic roles for the nonphotosynthetic plastid of the green alga Prototheca wickerhamii. Eukaryot Cell. 2005;4:253–61.PubMedPubMed CentralGoogle Scholar
Michaud
M, Gros V, Tardif M, Brugière S, Ferro M, Prinz WA, et al. AtMic60 is
involved in plant mitochondria lipid trafficking and is part of a large
complex. Curr Biol. 2016;26:627–39.PubMedGoogle Scholar
Cenci
U, Nitschke F, Steup M, Minassian BA, Colleoni C, Ball SG. Transition
from glycogen to starch metabolism in Archaeplastida. Trends Plant Sci.
2014;19:18–28.PubMedGoogle Scholar
Reyes-Prieto
A, Moustafa A, Bhattacharya D. Multiple genes of apparent algal origin
suggest ciliates may once have been photosynthetic. Curr Biol.
2008;18:956–62.PubMedPubMed CentralGoogle Scholar
Coppin
A, Varré J-S, Lienard L, Dauvillée D, Guérardel Y, Soyer-Gobillard M-O,
et al. Evolution of plant-like crystalline storage polysaccharide in
the protozoan parasite Toxoplasma gondii argues for a red alga ancestry. J Mol Evol. 2005;60:257–67.PubMedGoogle Scholar
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.

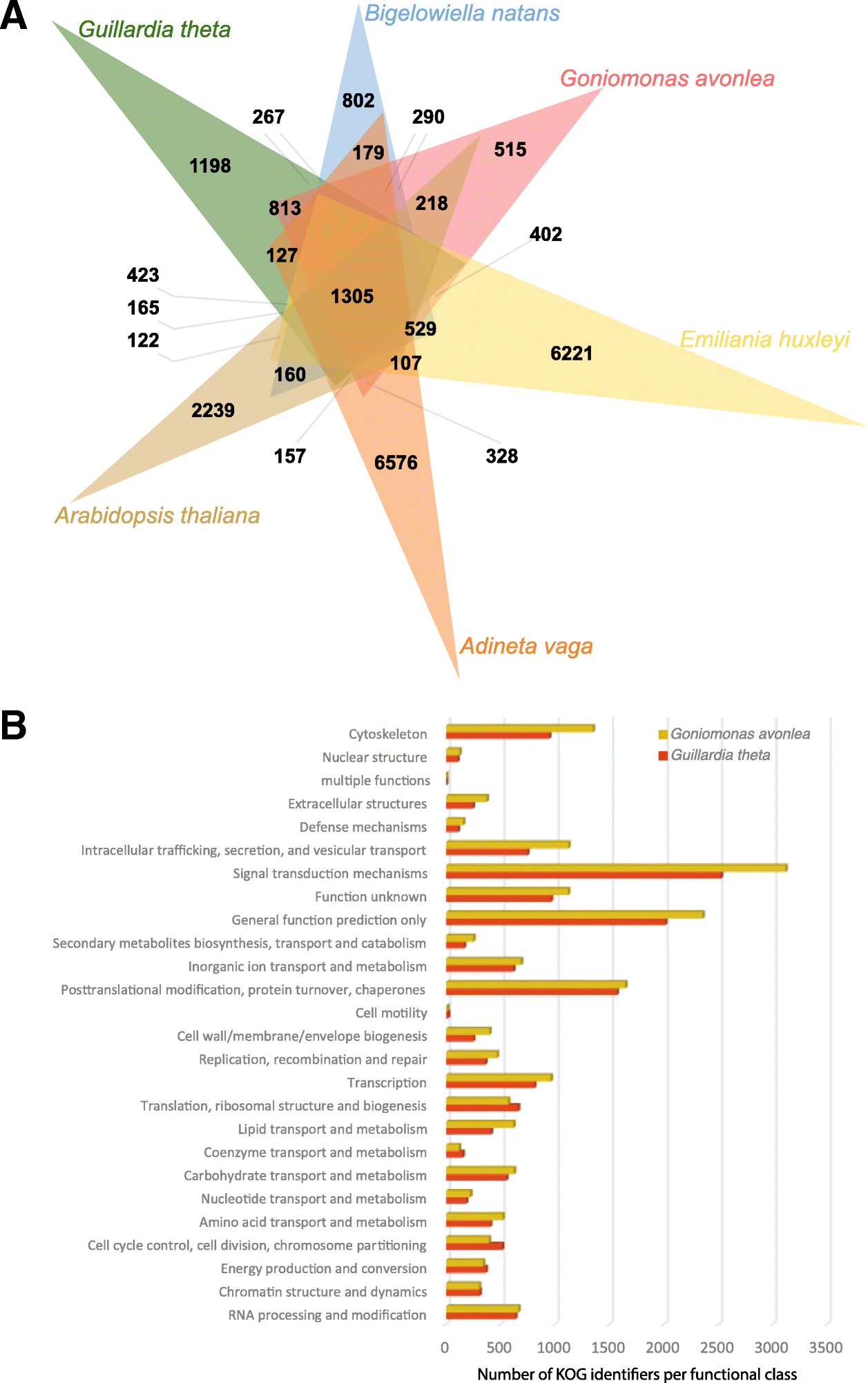


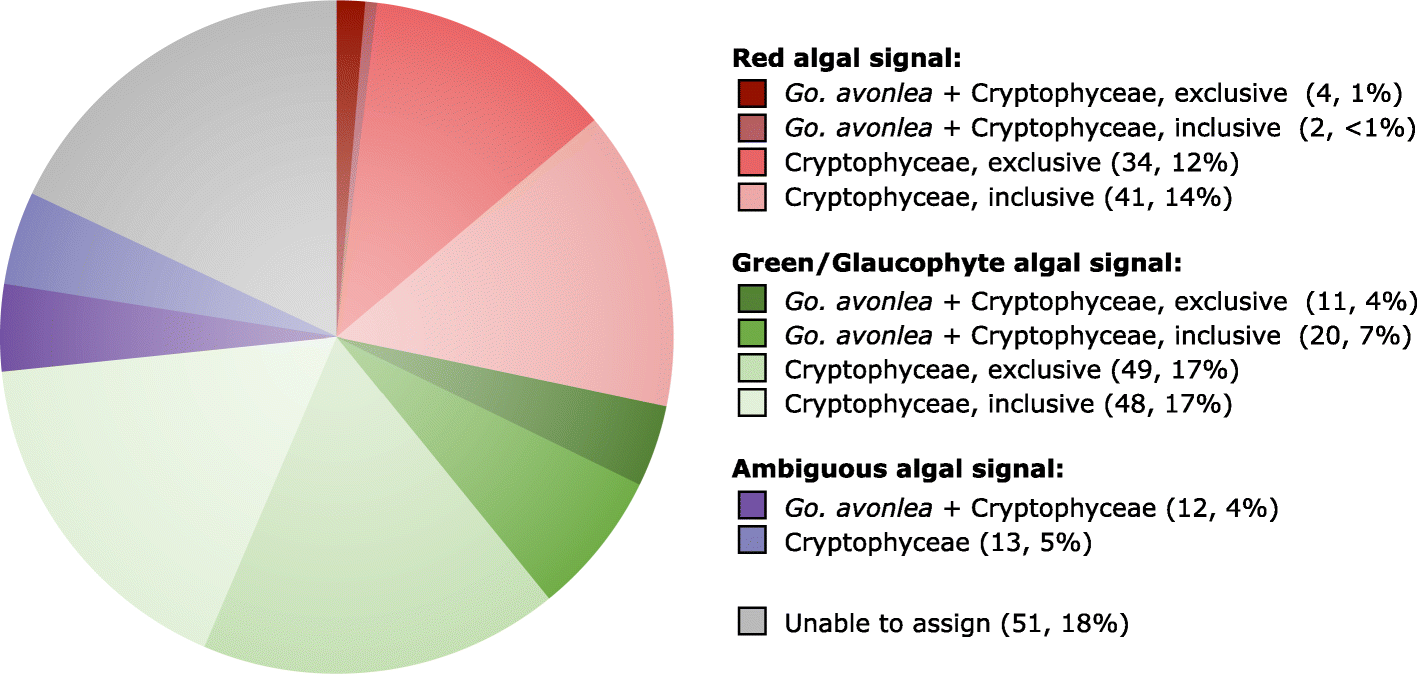
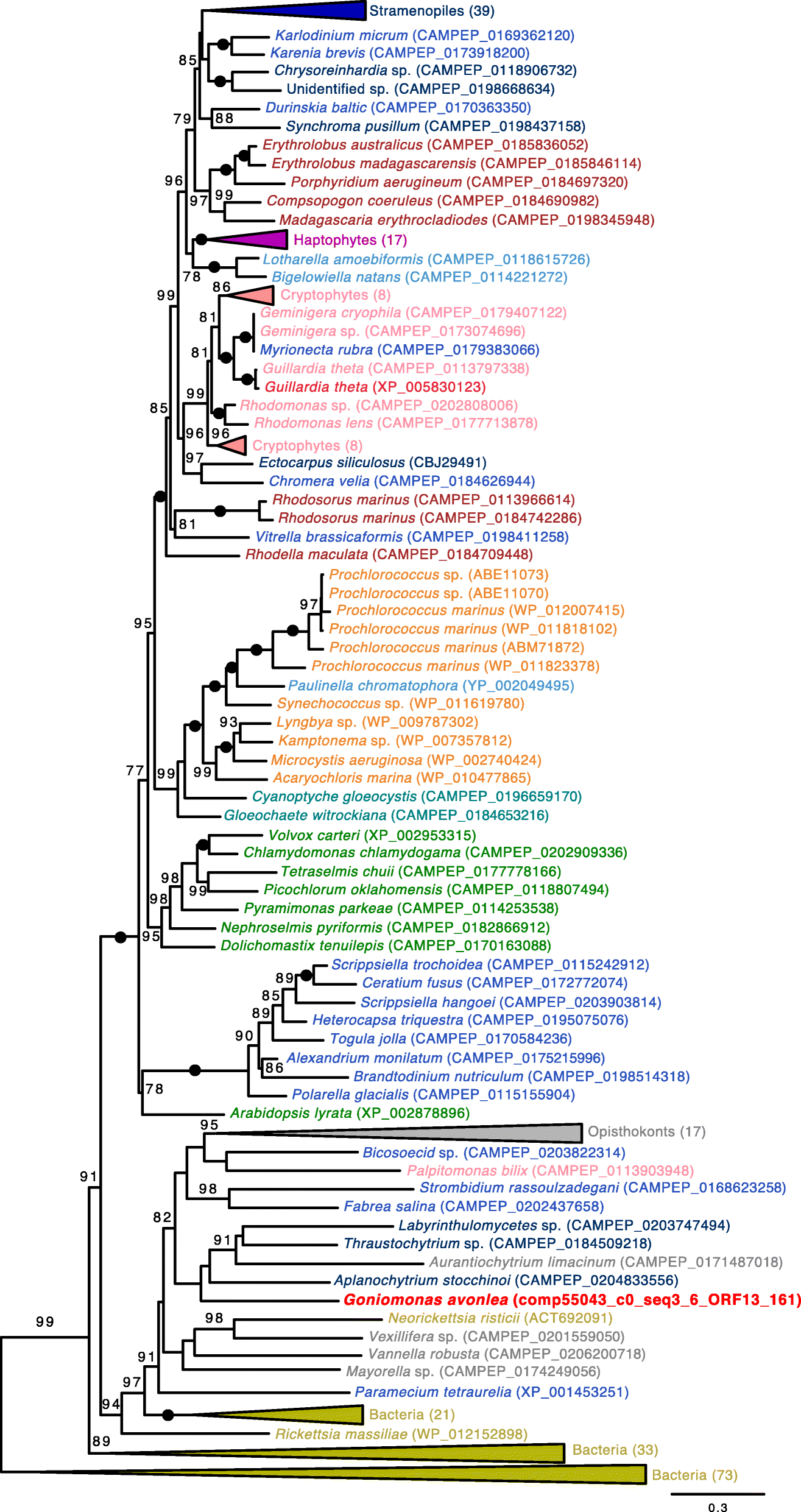

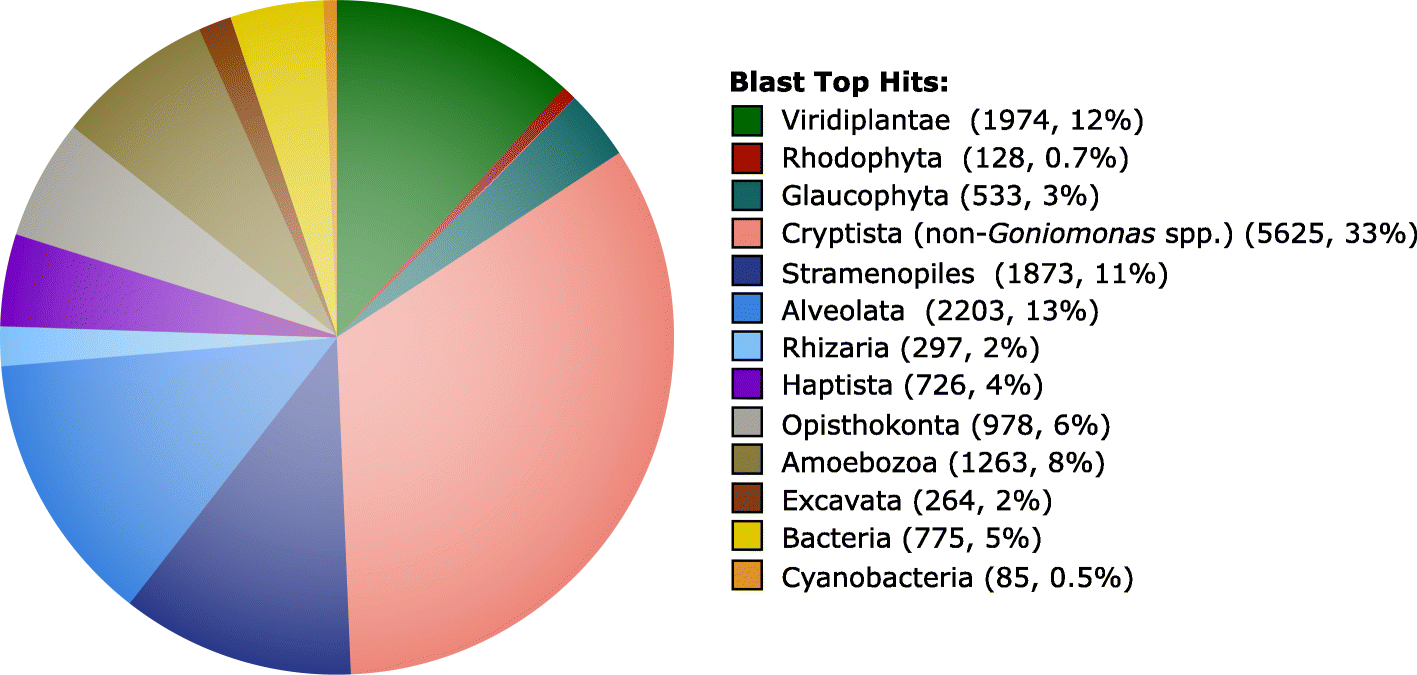

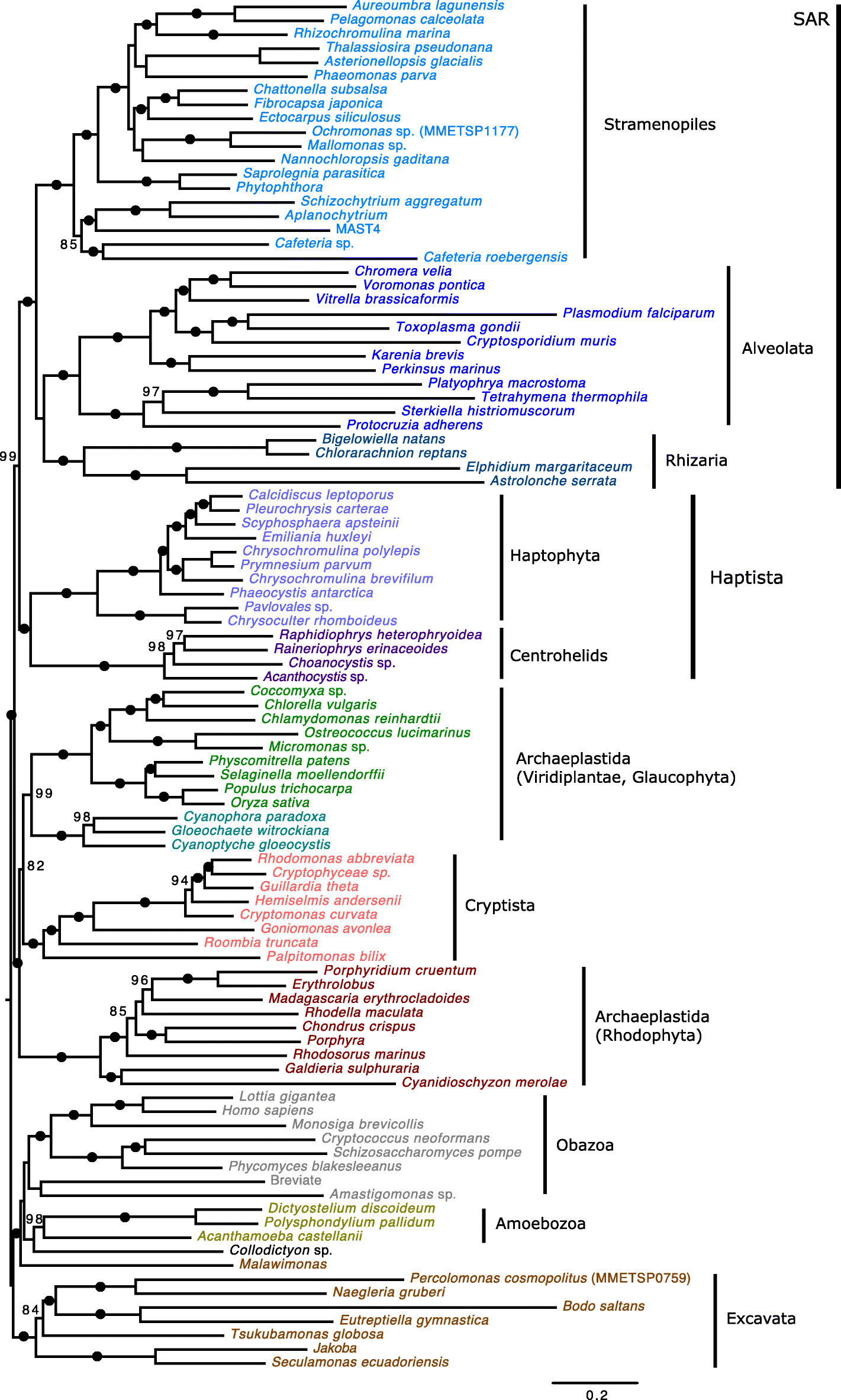

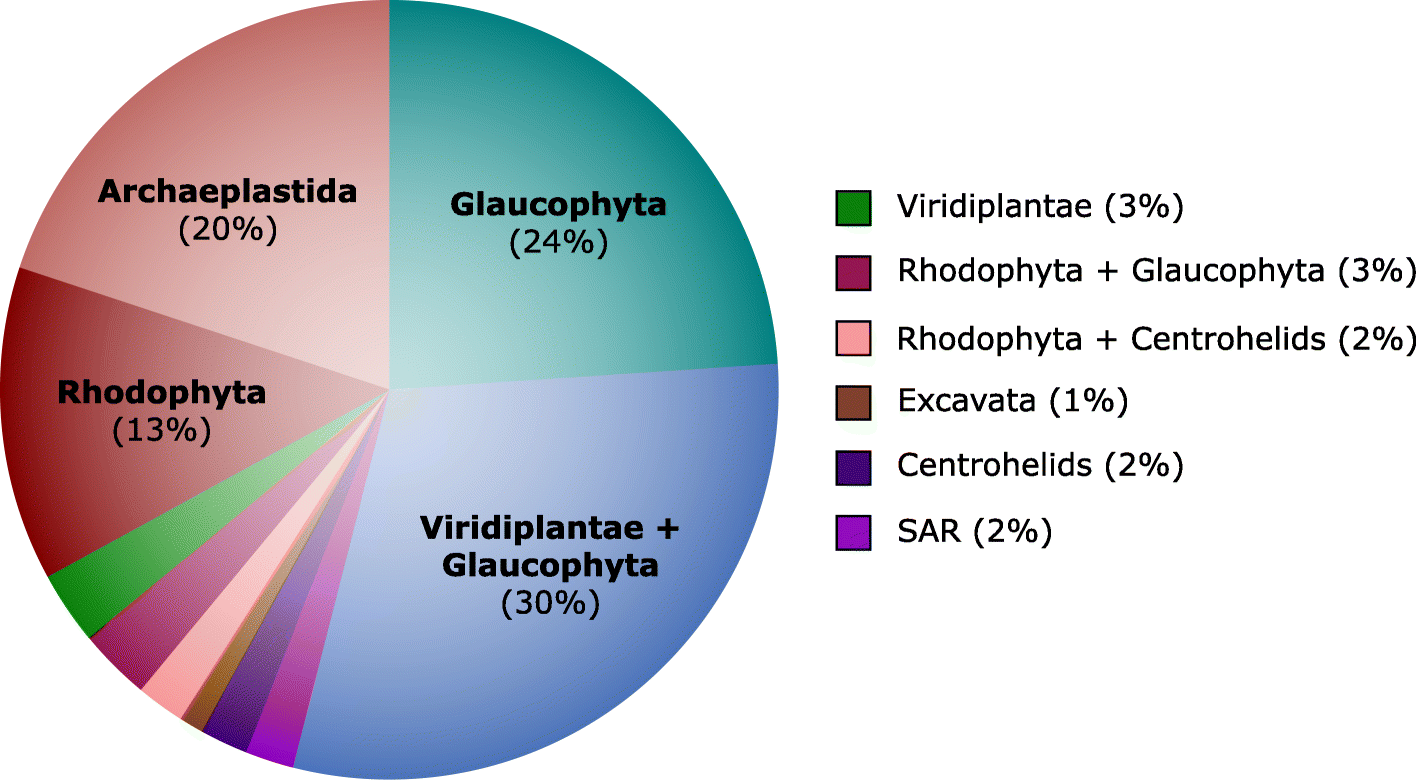

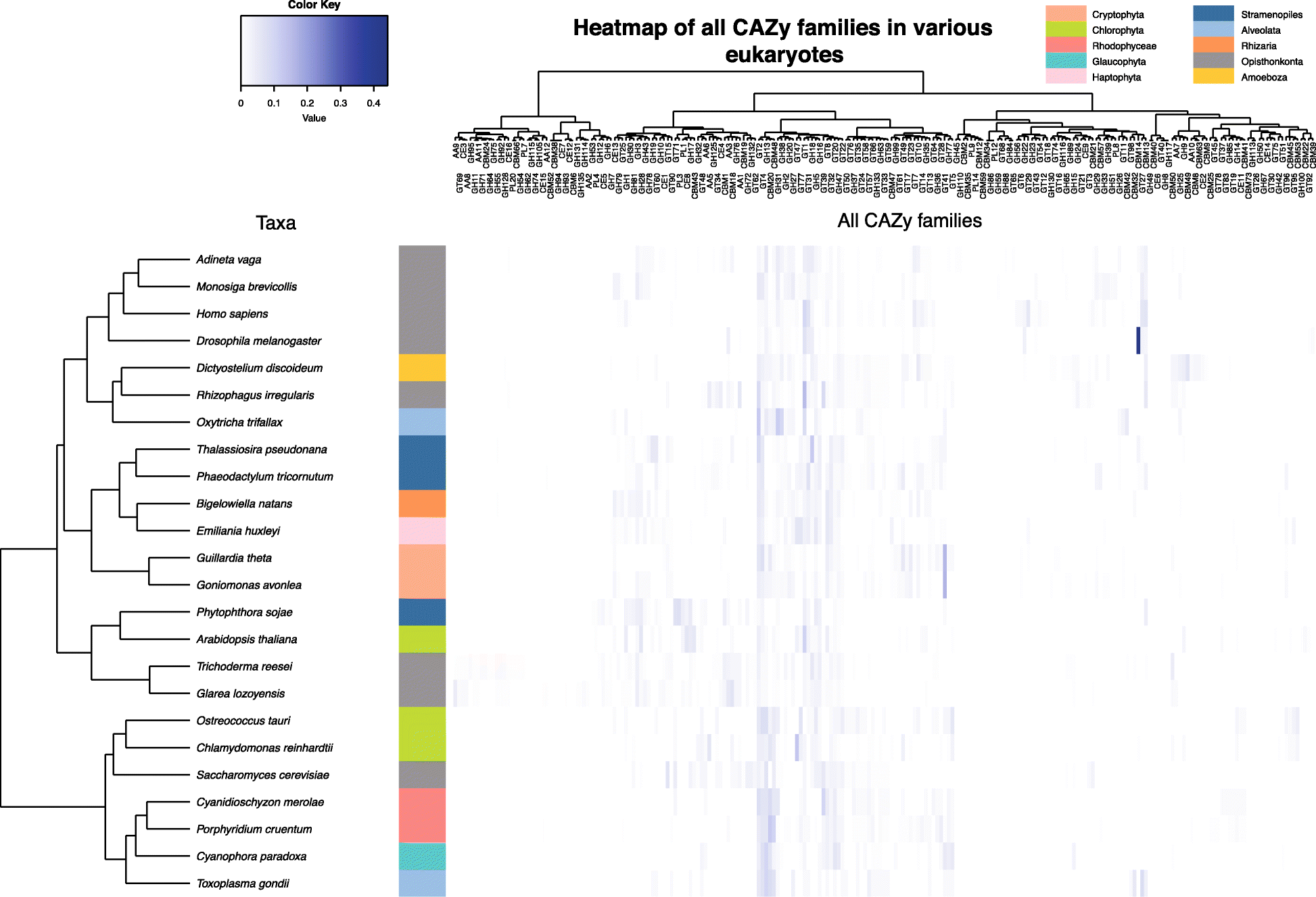

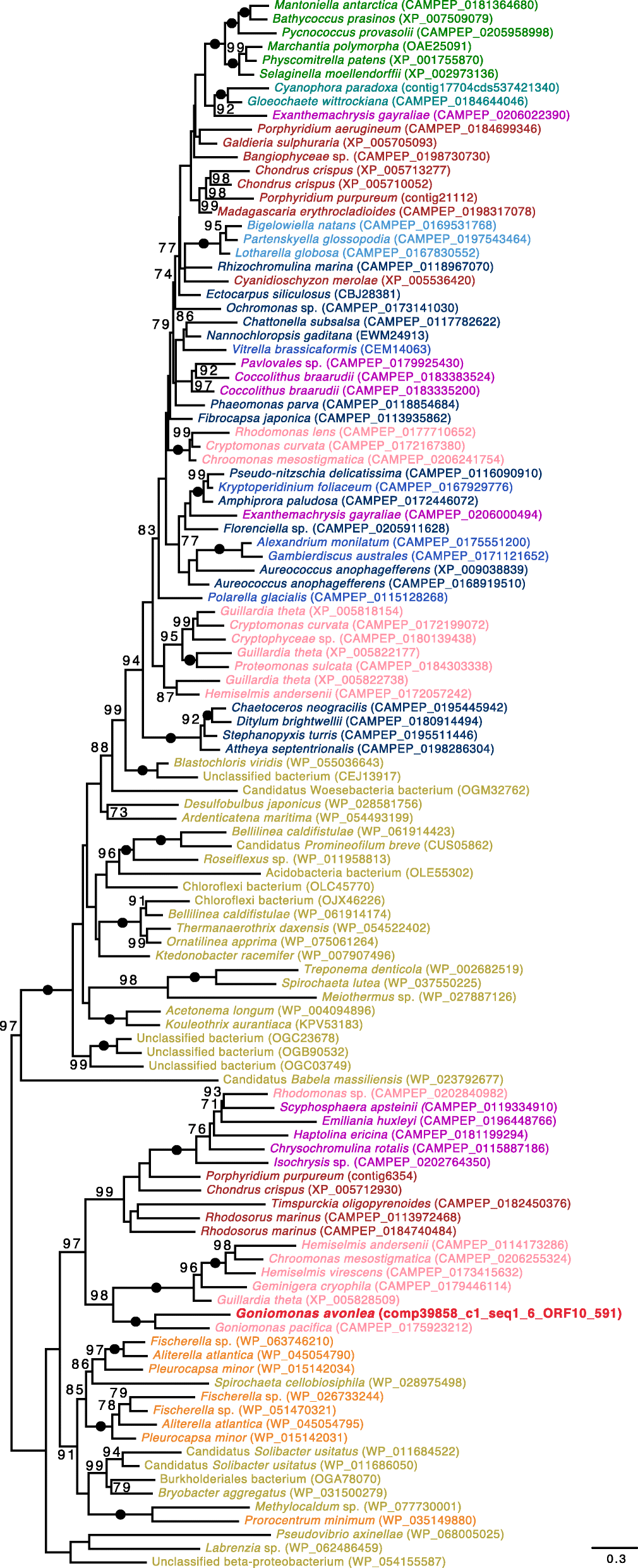

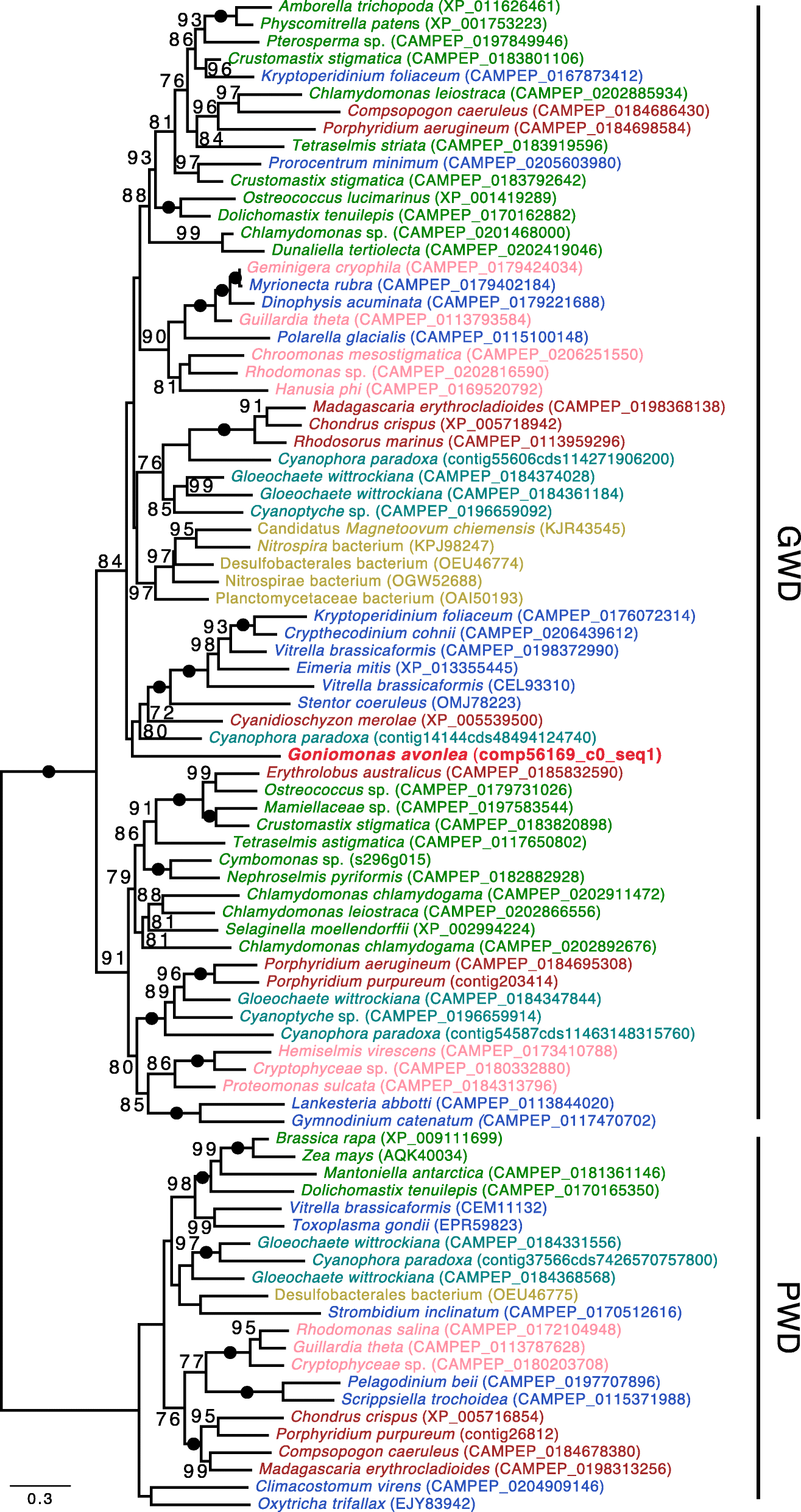
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.