The
55-million-year fossil record of horses (Family Equidae) has been
frequently cited as a prime example of long-term macroevolution. In the
second half of the nineteenth century, natural history museum exhibits
characteristically depicted fossil horses to be a single, straight-line
(orthogenetic) progression from ancestor to descendent. By the beginning
of the twentieth century, however, paleontologists realized that,
rather than representing orthogenesis, the evolutionary pattern of
fossil horses was more correctly characterized by a complexly branching
phylogenetic tree.
We conducted a systematic survey of 20 fossil horse
exhibits from natural history museums in the United States. Our
resulting data indicate that more than half (55%) of natural history
museums today still depict horse evolution as orthogenetic, despite the
fact that paleontologists have known for a century that the actual
evolutionary pattern of the Family Equidae is branching. Depicting
outmoded evolutionary patterns and concepts via museum exhibits, such as
fossils horses exemplifying orthogenesis, not only communicates
outmoded knowledge but also likely contributes to general misconceptions
about evolution for natural history museum visitors.
Fifty million people visit natural history museums in the U.S. each year (MacFadden et al. 2007).
These visitors expect to learn about current science and exciting
discoveries and trust these institutions to communicate correct and
up-to-date information (Falk and Dierking 2000; West 2005; Enseki 2006).
Once inside the museum, many studies have shown that most visitors
spend less than one minute at any given exhibit display (Cone and
Kendall 1978; Donald 1991; Allen 2004),
so museum scientists and exhibit developers are challenged to
communicate science in a highly focused, engaging manner (Bell et al. 2009).
This is oftentimes done with different combinations of specimens and
artifacts, graphic displays, passages of text, and, more recently,
multimedia technology (Hein 1998; Falk and Dierking 2000).
Many
natural history museums and other informal science-learning
institutions seek to communicate science content of societal relevance
to the public, including current hot-button topics and of relevance to
this study, evolution (NAS 2001; Diamond and Scotchmoor 2006).
With regard to evolution, since the second half of the nineteenth
century when many natural history museums were founded in the U.S., the
fossil record of horses has frequently been depicted in exhibits
communicating long-term (macro-) evolution (Gould 2002).
North
America has been home to the horse family (Equidae) over the past 55
million years (since the Eocene epoch), and fossil horses are widespread
on this continent during this time (MacFadden 1992, 2005).
Nineteenth century paleontologists in the U.S. made extensive
collections of fossil horses from the western territories that allowed
them to piece together a sequence depicting horse evolution. As a result
of the abundant fossil record and grand discoveries during the
nineteenth century, fossil horses were prominently displayed in natural
history museums that display evolutionary content (Clark 2008; Dyehouse 2011; Fig. 1). Prominent twentieth century paleontologists such as George Gaylord Simpson (1944, 1953) and Stephen Jay Gould (1988, 2002)
championed fossil horses as a prime example of evolution, and the
popularity and potential impact of this group continues up to the
present day (Franzen 2010).
Fig. 1
Fossil horse
exhibit, which is part of the Darwin travelling exhibit (Denis Finnin
photo, reproduced with permission of the American Museum of Natural
History, NY)
A
century ago, paleontologists typically depicted the evolutionary
pattern of fossil horses in North America as a linear sequence from
smaller ancestor to progressively larger descendent (Fig. 2),
which fit well with a concept popular during the nineteenth century
called orthogenesis, or “straight-line evolution” (“ortho-,” Gr.,
straight, direct; “-genesis,” Gr., origin).
This concept, attributed to
Haacke (1893) and other European scientists (Simpson 1944),
carries along with it notions that evolution is progressive, represents
improvement, and is predestined; these were accepted during that time,
but, in modern scientific thought, they are not considered as part of
the theoretical framework of macroevolution (MacFadden 1992; Gould 2002). Nevertheless, since that time, this pattern of the fossil horse lineage was embodied in museum displays (Clark 2008)
and other media that communicated about evolution. Thus, this sequence
of fossil horses has been widely recognized as a classic, “textbook”
example of evolution (Gould 2002; Dyehouse 2011). More than a half century ago, Simpson (1944, p. 157) stated that: “The most widely cited example of orthogenesis, in any sense of the word, is the evolution of the horse.”
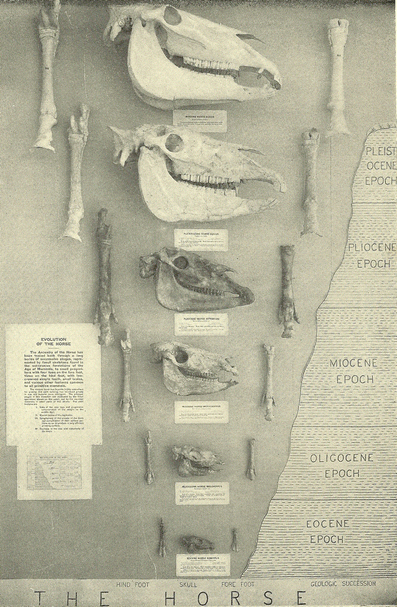
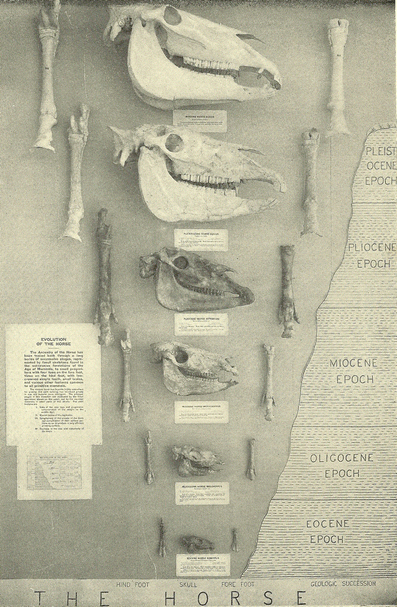
Fig. 2
Classic
straight-line (orthogenetic) depiction of fossil horse evolution in a
natural history museum exhibit in the early twentieth century (Matthew 1926).
In this depiction, fossil specimens are arranged in a temporal sequence
starting with the older fossil species of “eohippus” (Hyracotherium) at the bottom and ending with the genus of modern horse Equus at the top.
In addition to depicting orthogenesis, this graphic display implicitly
communicates nineteenth-century notions of evolution representing
progress or improvement
The
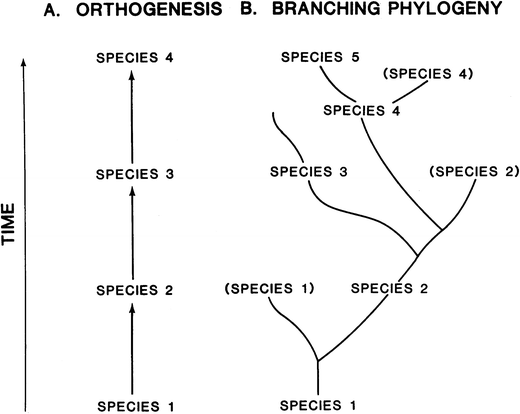
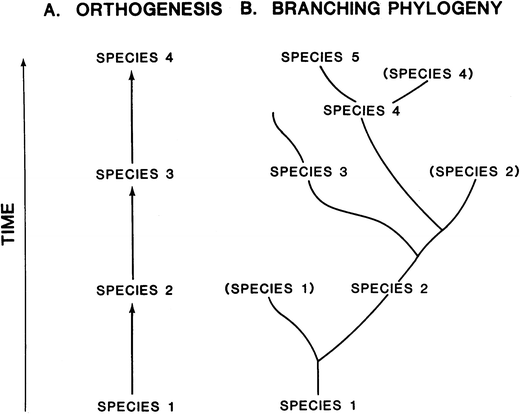
problem with depicting fossil horses as orthogenetic is that, by the
early twentieth century, paleontologists understood that, rather than a
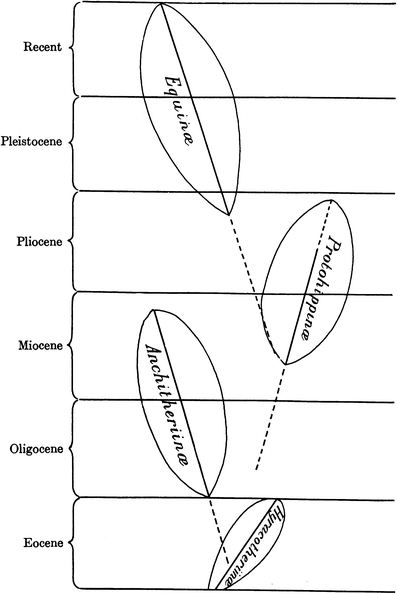
simple, straight-line sequence (Fig. 3A), the actual fossil record of horses was a complexly branching tree (Fig. 3B). The latter is exemplified by Gidley’s (1907; Fig. 4
here) branching diagram of the subfamilies of fossil horses in which
the Anchitheriinae overlaps in time with the Protohippinae during the
late Miocene and the Protohippinae overlaps in time with the Equinae. In
the orthogenetic model (and more recent concept of anagenesis), this
overlap cannot occur as a macroevolutionary pattern because ancestral
and descendant taxa (species) do not overlap in time, i.e., the former
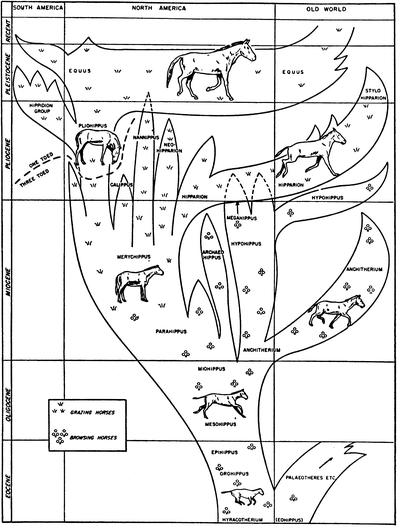
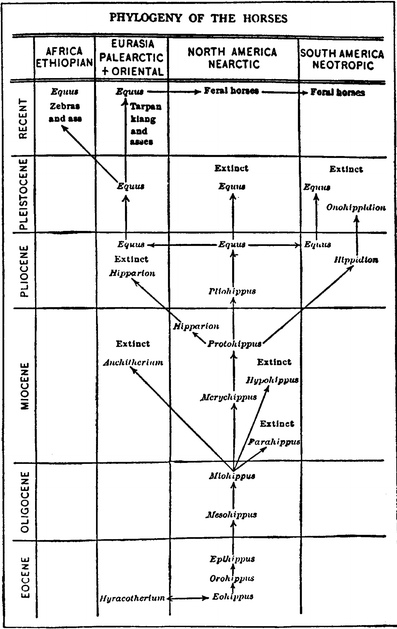
is replaced in time by the latter (Fig. 3). Following on the branching pattern, several decades later, Matthew (1930; Fig. 5)
depicted the evolution of fossil horses with numerous genera and
independent lineages that overlapped in time. This scheme was further
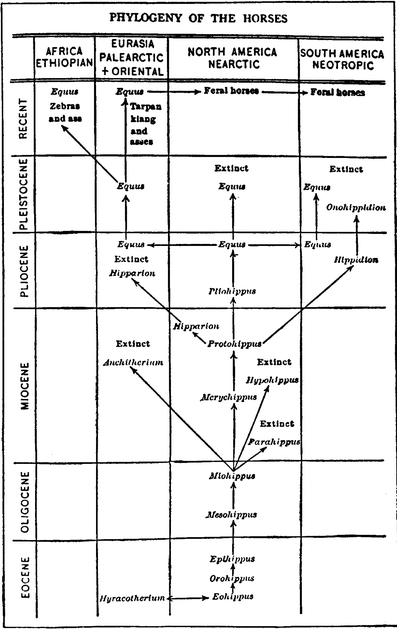
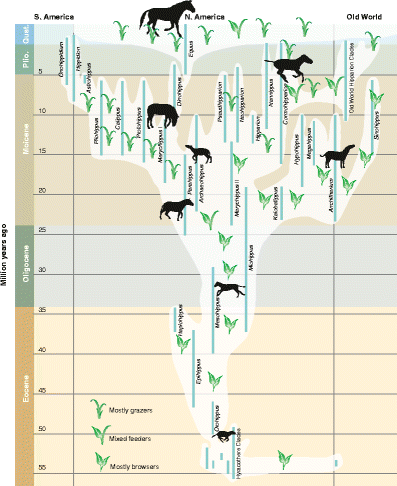
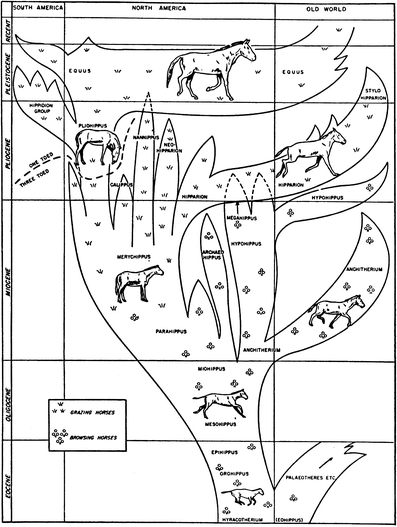
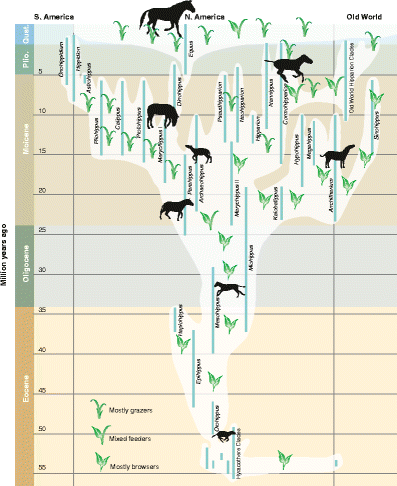
elaborated upon by Simpson’s (1951; Fig. 6) now widely cited horse phylogenetic tree, which continues in its essential pattern to the present day (MacFadden 1992, 2005 [Fig. 7]; Franzen 2010).
Thus, to summarize our current state of knowledge, instead of a linear
sequence in which ancestral species evolve directly into their
descendants, the evolutionary tree of horses is bushy, with many species
overlapping in time, multiple originations, and frequent extinctions.
Fig. 3
A model to show the difference between orthogenesis (A) and branching (B) speciation (or phylogeny; from MacFadden 1992)
Fig. 4
Gidley’s (1907)
phylogeny of the subfamilies of fossil horses from North America
showing some overlap, which thereby negates orthogenesis as an
evolutionary mode
Fig. 5
Matthew’s (1930) horse phylogeny, published in the popular magazine Scientific American, showing significant branching of numerous genera
Fig. 6
Simpson’s (1951)
horse phylogeny that elaborates upon previous workers such as Matthew
and represents the principal pattern of macroevolution of the Family
Equidae that is elaborated upon up to the present day
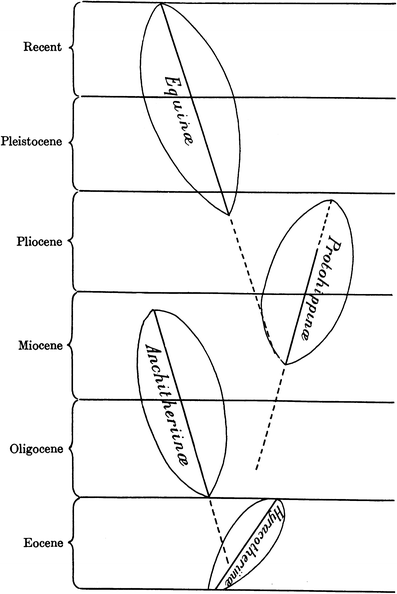
Fig. 7
Phylogeny of the Family Equidae based on a recent updated graphic (MacFadden 2005). Reproduced, with permission from the American Association for the Advancement of Science
Despite
this knowledge, to this day, the classic story of horse evolution in
museums, books, and other media is still oftentimes depicted as
orthogenetic. In addition to being factually incorrect within a modern
scientific context of what we know about evolution, orthogenesis brings
along with it baggage about evolution being largely progressive,
deterministic, and representing improvement (MacFadden 1992; Gould 2002).
These notions associated with orthogenesis therefore likely contribute
to part of the general public’s misunderstanding, or incomplete
knowledge, about fundamental aspects of evolution (MacFadden 1992; Diamond and Scotchmoor 2006; Spiegal et al. 2006; MacFadden et al. 2007). In Jonathan Wells’ (2000) book entitled Icons of Evolution: Science or Myth? Why much of what we teach about evolution is wrong,
the author cites fossil horses as one of ten fundamental examples of
evolution. He also accurately describes some of the problems associated
with the miscommunication and states (Wells 2000,
195): “Since the 1950s, neo-Darwinian paleontologists have been
actively campaigning to replace the old linear picture of horse
evolution with the branching tree.” As we will see below, this campaign
has not been particularly successful.
We
assert that, despite its roots in nineteenth century evolutionary
theory, the outmoded scientific concept of orthogenesis is still widely
communicated to the general public through various media, including
natural history museum exhibits. We use fossil horses to test this
hypothesis because of their widespread use as fundamental evidence for
macroevolution (e.g., Gould 2002).
Based
on a systematic review of natural history and science museums websites,
we contacted staff at 36 institutions with current permanent or
traveling exhibitions referring to evolution or fossil mammals
requesting digital photographs of their horse evolution exhibits.
Seventeen museums confirmed they currently had this type of exhibit. We
received 91 photographs from these museums, including different views of
the same exhibit. We ultimately selected 26 photographs based on
quality and completeness. Three museums had online exhibitions; these
were not contacted for photographs because we were able to access the
exhibits directly. We considered only the graphic representation of
horse evolution, and text descriptions were not included in the coding
scheme because: (a) with only one minute or less for most visitors at a
particular exhibit (Cone and Kendall 1978; Donald 1991),
we assume that the text is not fully read; and (b) studies show
information presented in exhibit text is less easily learned and
recalled (Hooper-Greenhill 1994).
Given what is known about general learning behavior (i.e., the
time-tracking studies cited above), the visual impact of non-text
exhibit components represents a significant factor in science
communication; therefore, we contend that our concentration on graphics
and related three-dimensional content (as opposed to the text) is
justified. Each exhibit was evaluated to determine the number of
components present. A component was defined as each array referring to
horse evolution. For example, a set of skulls, feet, skeletons, or an
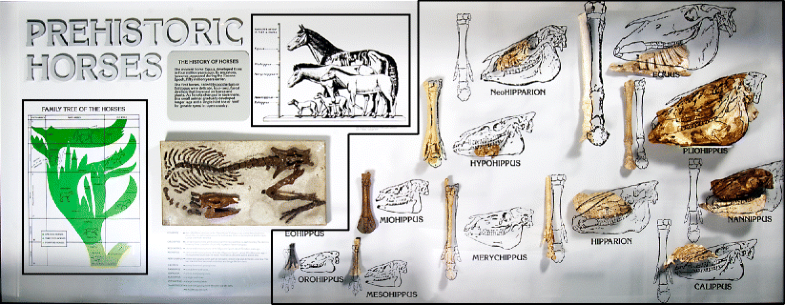
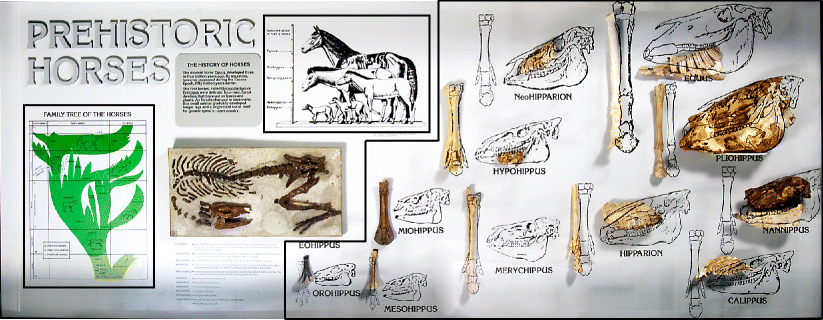
image of horse evolution was considered a single component. Figure 8 shows one exhibit that we divided into three components.
Fig. 8
Horse evolution
exhibit at the Panhandle Plains Historical Museum, Canyon, Texas,
showing the three individual components (indicated in the black boxes) that we coded during our study. Reproduced with permission
We
used Content Analysis as the main methodology for this research because
it is a way to study content in human communications such as text,
images, maps, or symbols (Krippendorf 2004).
A coding guide was developed and tested. A set of images with
evolutionary depictions of species and taxa other than horses was used
to assess reliability during training among two coders (Cohen’s kappa
0.91; Cohen 1960).
In the actual fossil horse exhibit data, coders independently rated
each component as either orthogenetic or branching (Cohen’s kappa 0.94).
Once all elements of the exhibit were coded, each complete exhibit was
then classified using a five-point qualitative scale: orthogenetic, primarily orthogenetic, mixed orthogenetic and branching, primarily branching, or branching. Exhibits, or exhibit components, were coded to be orthogenetic
if there was no depiction of species overlapping in time, and if the
species were depicted in a straight-line scheme, gradually growing in
size and relative complexity (e.g., Fig. 1).
Typically, the number of species displayed in orthogenetic displays was
fewer compared with branching displays. Exhibits, or exhibit
components, were considered to be branching if there was a clear tree with the same emphasis on all species and/or evidence of coexistence of species (e.g., Fig. 7). Exhibit components that mixed these patterns were coded as mixed orthogenetic and branching (e.g., Fig. 9).
Fig. 9
Horse evolution
exhibit currently on display at the International Museum of the Horse,
Kentucky Horse Park, Lexington, KY. This exhibit mixes components that
communicate orthogenesis (the skeletons arranged in the foreground)
along with branching evolution (the phylogenetic tree graphic behind the
skeletons). Reproduced with permission
For
exhibits with more than one component, the approach, size, and location
of each unit were qualitatively assessed by each coder. Intercoder
reliability on the overall assessment was 0.7 using Cohen’s kappa (Cohen
1960), and it was considered appropriate (Landis and Koch 1977; Lombard et al. 2002). Disagreements were discussed and resolved (Lombard et al. 2002).
In total, we analyzed 37 components within 20 physical and online
exhibits depicting fossil horses in the United States (Table 1).
Table 1
Classification of fossil horse exhibits and museums included in the study
Museum
State
Number of displays
Pattern of depiction
Yearly attendance
University of Nebraska State Museum
NE
1
O
86,182
Field Museum of Natural History
IL
2
O
1,212,475
Harvard Museum of Natural History
MA
1
O
150,000
U.S. National Museum of Natural History, Smithsonian Institution
DC
1
O
5,542,000
Darwin (traveling exhibita)
N/A
2
O
N/A
Amherst College, Beneski Museum of Natural History
MA
1
O
25,000
Kansas University Natural History Museum and Biodiversity Research Center
KS
2
O
46,000
American Museum of Natural History
NY
1
O
4,000,000
Carnegie Museum of Natural History
PA
1
PO
497,000
Natural History Museum of Los Angeles County
CA
1
PO
650,000
Peabody Museum of Natural History
CT
4
PO
145,000
International Museum of the Horse
KY
2
M
200,000
University of Michigan Museum of Natural History
MI
2
M
7,000
University of Oregon Museum of Natural and Cultural History
OR
4
M
200,000
University of California Museum of Paleontology-Berkeley
CA
2
M
b
Florida Museum of Natural History
FL
3
PB
275,000
Denver Museum of Nature and Science
CO
1
PB
1,252,300
Santa Barbara Museum of Natural History
CA
1
PB
110,000
The Horse (traveling exhibita)
N/A
1
B
N/A
Panhandle Plains Historical Museum
TX
3
B
75,000
Total summed yearly attendance
14,472,957
The attendance figures for each museum were taken from AAM (2007).
The cumulative attendance figures for the two travelling exhibits were
not available (n/a), and those for the three Web exhibits are not
reported
O orthogenetic, PO primarily orthogenetic, M mixed (equally orthogenetic and branching), PB primarily branching, B branching
aDeveloped by The American Museum of Natural History, New York
b1.2
million Web visitors per month according to University of California at
Berkeley; not included in total summed yearly attendance
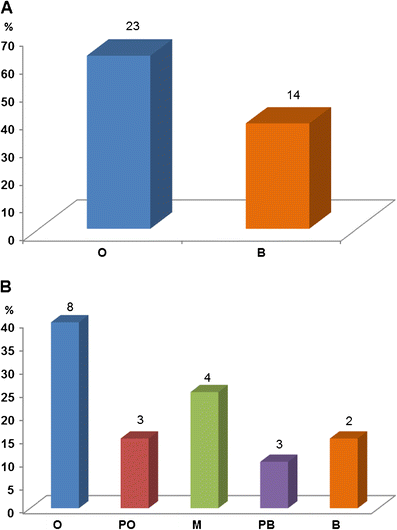
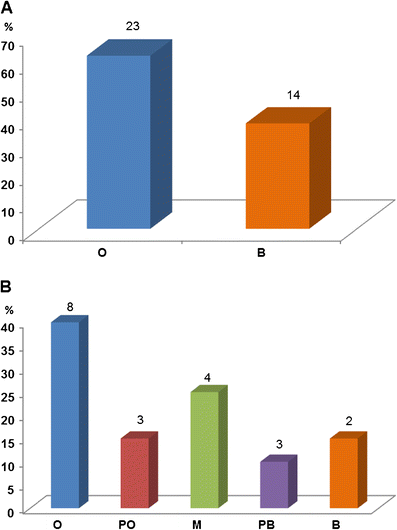
Our
survey data demonstrate that within fossil horse exhibits, the
orthogenetic pattern is the most common type of depiction in both
individual exhibit components (62%) as well as complete exhibits (40%,
Fig. 10). Moreover, when orthogenetic and primarily orthogenetic exhibits are grouped, they represent 55% of the total sample; exhibits considered to be mixed orthogenesis and branching represent 20%; and the branching and primarily branching
groups combined represent the remaining 25%. We therefore conclude from
these data that orthogenesis is a widespread depiction in fossil horse
exhibits in U.S. natural history museums today.
Fig. 10
Distribution of
fossil horse displays in natural history museums communicating
orthogenesis versus branching patterns of evolution. a Orthogenesis versus branching in the 37 individual components analyzed. b Overall pattern of evolution depicted in multi-component exhibits from 20 museums (see Table 1). Abbrev: O, orthogenetic; PO, primarily orthogenetic; M, mixed O and B; PB, primarily branching; B, branching. Numbers at the top of each bin represent sample size (N)
Given
these results, why is orthogenesis so prevalent in current fossil horse
exhibits, when paleontologists have known for a century that it is an
incorrect way to depict the pattern of horse evolution? We contend that
the answer to this question is complex and likely involves multiple
factors, including:
1.
An inability and inertia of
researchers studying fossil horses to effectively communicate to other
scientists (particularly outside of their discipline), museum curators,
exhibit designers, text-book authors, and the public. As demonstrated
above, knowledge that the evolutionary pattern of fossil horses is
branching has been current for a century, but this fact has not been
effectively communicated outside the narrow sphere of practicing
professional paleontologists.
2.
Museum exhibits are expensive to build, particularly in grand exhibition halls costing millions of dollars. For example, the Hall of Florida Fossils: Evolution of Life and Land
at the Florida Museum of Natural History was completed in 2004 at a
cost of about $2.5 million dollars, which equates to about $600 per
square foot. This is fairly typical for a multimedia, specimen-rich
major exhibition hall. Accordingly, and from a pragmatic point of view,
after the rush of activity passes with newly opened exhibit halls,
museum professionals typically move on to the next project, and
therefore physical renovations and scientific updates to prior projects
are slow to be implemented. It therefore is a challenge to incorporate
the most current scientific concepts into physical exhibits (although
with modern multimedia displays and cyberexhibits, this inertia has the
potential to change in the future). Moreover, as Dyehouse (2011)
has shown, even when museum curators are sensitive to the issue of
public perception of orthogenesis, this pattern or schema may
unintentionally become incorporated into updated physical exhibits.
3.
With only a minute or less for
learning opportunities at individual exhibits, museum interpretive
content graphics are frequently presented in a simplified manner, e.g.,
with orthogenesis rather than a branching schema. Studies of museum
visitors, however, show that, when abstract concepts are oversimplified,
misconceptions can result (Bishop and Anderson 1990; Matuk and Uttal 2008).
Thus, within the context of our study, we assert that orthogenetic
depictions provide incorrect information for the sake of simplicity and
also lead to fundamental misconceptions about the pattern of
macroevolution.
4.
Research from the cognitive and
learning sciences demonstrates that common ways of visually representing
evolution contribute to fundamental misconceptions, especially among
people with less well-developed backgrounds in science, i.e., where
intuition results in a naïve conception framework for learning. Of
relevance to the current study, depictions that utilize vertical space
with more recent developments placed at the top imply progress or
improvement and lead individuals to conceive of evolution as a
teleological (purpose-driven) process (Tversky 2011).
Furthermore, linear depictions encourage anagenic (direct sequence from
ancestor to descendent) interpretations of speciation (Catley et al. 2010; Novick et al. 2011).
Other
research has shown that branching-tree depictions that accurately
represent macroevolutionary patterns are difficult for students and
museum visitors to grasp (Gregory 2008; Evans et al. 2010).
The solution, we contend, is not to settle for simpler, more accessible
museum displays. Inappropriate depictions of evolution in museums
should be no less accurate than their textbook counterparts (Catley and
Novick 2008). The 15 institutions surveyed here that depict horse evolution as either orthogenetic, primarily orthogenetic, or mixed have an estimated visitation of more than 12 million people per year (Table 1)
to their physical exhibits (when cyberexhibits are added, this figure
would be greater). While we do not assert that every visitor to each of
these institutions saw the horse evolution exhibit, the potential exists
for scientific miscommunication on a large scale, which likely
contributes to the low public acceptance, and generally poor
understanding, of evolutionary concepts in the U.S (Miller et al. 2006).
We
have used fossil horse exhibits as an example of orthogenesis, yet the
impact of this cognitive framework is pervasive and extends to other
organisms, both living and extinct, including humans (Scott and Guisti 2006).
The common quip among the non-believing general public, “Don’t tell me
I’m descended from a monkey,” derives from an orthogenetic framework in
which humans evolved directly from apes. The impact of orthogenesis also
transcends museum exhibits and can be found in many other media that
communicate about science, including textbooks, newspapers, magazines,
documentaries, and the enormous access provided by the Web. This lack of
public understanding of evolution in the U.S. has profound consequences
for overall understanding within the life and natural sciences and also
impacts applied aspects of society, such as human medicine.
While
the scope of our study concentrated on natural history museums in the
U.S., anecdotal evidence (MacFadden, personal observations; also see,
e.g., Franzen 2010,
but no comprehensive list is currently available) suggests that
orthogenetic frameworks are widespread in museums in other countries,
and this pattern is not just found in fossil horses exhibits, but in
many other kinds of depictions, particularly involving evolutionary
sequences and interpreted phylogenies of groups with a fossil record.
The magnitude and pervasiveness of this style of content presentation
thus results in a worldwide challenge for science communication and
science literary.
Science
evolves with the advent of new discoveries and ongoing research, as is
the case for the classic story of fossil horse evolution that has
developed over the past 150 years. Despite the scientific discoveries
that paleontologists have made “in the trenches,” these advances in
knowledge have been slow to enter into the general body of scientific
knowledge about evolution. One consequence of this inertia has been that
natural history museum exhibits have been slow to incorporate recent
scientific advances into the content of their exhibits. Misinformation
communicated in this manner likewise contributes to the museum visitors’
misunderstanding about fundamental concepts and examples of
evolutionary theory. If science literacy is a priority in modern society
and the public needs to make informed decisions in their everyday lives
that depend upon understanding evolution, then it is incumbent on
scientists and museum professionals to more effectively communicate in
media such as museum exhibits.
We
thank our contacts at the museums surveyed for providing exhibit
photographs. This research was partially supported by U.S. National
Science Foundation (NSF 09-66884, OISE, EAR, OISE), Vertebrate
Paleontology Fund, Florida Museum of Natural History, and student
scholarships provided by the Fossil Club of Lee County and Southwest
Florida Fossil Club.
AAM
(American Association of Museums). The Official Museum Directory, 38th
Edition. New Providence New Jersey: National Register Publishing; 2007.Google Scholar
Allen S. Designs for learning: studying science museum exhibits that do more than entertain. Sci Educ. 2004;88 Suppl 1:S17–33.View ArticleGoogle Scholar
Bell
P, Lewenstein B, Shouse AE, Feder MA, editors. Learning science in
informal environments: people, places, and pursuits. Washington DC:
National Academies Press; 2009.Google Scholar
Bishop B, Anderson C. Student conceptions of natural selection and its role in evolution. J Res Sci Teach. 1990;27:415–27.View ArticleGoogle Scholar
Catley
KM, Novick LR. Seeing the wood for the trees: an analysis of
evolutionary diagrams in biology textbooks. BioScience 2008; 58:976–987.Google Scholar
Catley
KM, Novick LR, Shade CK. Interpreting evolutionary diagrams: when
topology and process conflict. J Res Sci Teach 2010; 47:861–882.Google Scholar
Clark CA. God—or gorilla: images of evolution in the Jazz Age. Baltimore: The Johns Hopkins University Press; 2008.Google Scholar
Cohen J. A coefficient of agreement for nominal cases. Educ Psychol Meas. 1960;20:137–46.View ArticleGoogle Scholar
Cone
CA, Kendall K. Space, time, and family interaction: visitor behavior at
the Science Museum of Minnesota. Curator. 1978;21:245–58.View ArticleGoogle Scholar
Donald JG. The measurement of learning in the museum. Can J Educ. 1991;16:371–82.View ArticleGoogle Scholar
Dyehouse
J. “A textbook case revisited”: visual rhetoric and series patterning
in the American Museum of Natural History’s horse evolution displays.
Tech Commun Quart. 2011;20:327–46.View ArticleGoogle Scholar
Enseki C. Public trust and accountability. New Standard. Washington DC: American Association of Museums; Summer 2006.Google Scholar
Evans
M, Frazier B, Hazel A, Kiss A, Lane JD, Spiegal A, Diamond J. Tree
thinking: do pictorial representations of phylogenetic relationships
help or hinder museum visitors’ understanding of evolution? Abstract of
talk presented at the Tree of Life Conference, Carnegie Museum of
Natural History (August 2010).Google Scholar
Falk J, Dierking L. Learning from museums. Walnut Creek, CA: AltaMira Press; 2000.Google Scholar
Franzen JL. The rise of horses. Baltimore: The Johns Hopkins University Press; 2010.Google Scholar
Gidley JW. Revision of the Miocene and Pliocene Equidae of North America. Bull Am Mus Nat Hist. 1907;23:865–934.Google Scholar
Gould SJ. The case of the creeping fox terrier clone. Nat Hist. 1988;97:16–24.Google Scholar
Gould SJ. The structure of evolutionary theory. Cambridge, MA: Harvard University Press; 2002.Google Scholar
Gregory TR. Understanding evolutionary trees. Evo Edu Outreach. 2008;1:121–37.View ArticleGoogle Scholar
Haacke W. Gestaltung und Verebung. Leipzig: Weigel; 1893.Google Scholar
Hein GE. Learning in the museum. New York: Routledge; 1998.Google Scholar
Hooper-Greenhill E. Museums and their visitors. New York: Routledge; 1994.Google Scholar
Krippendorf K. Content analysis. An introduction to its methodology. Thousand Oaks, CA: Sage Publications; 2004.Google Scholar
Landis R, Koch G. The measurement of observer agreement for categorical data. Biometrics. 1977;33:159–74.View ArticlePubMedGoogle Scholar
Lombard
MJ, Snyder-Duch J, Campanella C. Content analysis in mass
communication. Assessment and reporting of intercoder reliability. Human
Commun Res. 2002;28:587–604.View ArticleGoogle Scholar
MacFadden
BJ. Fossil horses: systematics, paleobiology, and evolution of the
Family Equidae. Cambridge: Cambridge University Press; 1992.Google Scholar
National Academy of Sciences. Science, evolution, and creationism. Washington, DC: NAS; 2001.Google Scholar
Novick
LR, Shade CK, Catleyb KM. Linear versus branching depictions of
evolutionary history: implications for diagram design. Top Cogn Sci.
2011;3:536–59.View ArticlePubMedGoogle Scholar
Scott M, Guisti E. Designing human evolution exhibitions: insights from exhibitions and audiences. Mus Soc Issues. 2006;1:49–67.View ArticleGoogle Scholar
Simpson GG. Tempo and mode in evolution. New York: Columbia University Press; 1944.Google Scholar
Simpson
GG. Horses: the story of the horse family in the modern world and
through sixty million years of history. New York: Oxford University
Press; 1951.Google Scholar
Simpson GG. The major features of evolution. New York: Columbia University Press; 1953.Google Scholar
Spiegal AN, Evans M, Graham W, Diamond J. Museum visitors’ understanding of evolution. Mus Soc Issues. 2006;1:69–85.View ArticleGoogle Scholar
Wells
J. Icons of evolution: science or myth? Why much of what we teach about
evolution is wrong. Washington DC: Regnery Publishing; 2000.Google Scholar
West
R. The lay of the land: the current context for communicating evolution
in natural history museums. Rep Natl Cent Sci Educ. 2005;25:21–5.Google Scholar
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.




Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.