Dinosaur
embryos are among the rarest of fossils, yet they provide a unique
window into the palaeobiology of these animals. Estimating the
developmental stage of dinosaur embryos is hindered by the lack of a
quantitative method for age determination, by the scarcity of material,
and by the difficulty in visualizing that material. Here we present the
results of a broad inquiry, using 3D reconstructions from X-ray computed
tomography data, into cranial ossification sequences in extant saurian
taxa and in well-preserved embryos of the early branching sauropodomorph
dinosaur Massospondylus carinatus. Our findings support
deep-time conservation of cranial ossification sequences in saurians
including dinosaurs, allowing us to develop a new method for estimating
the relative developmental percentage of embryos from that clade. We
also observe null-generation teeth in the Massospondylus carinatus
embryos which get resorbed or shed before hatching, similar to those of
geckos. These lines of evidence allow us to confidently estimate that
the Massospondylus carinatus embryos are only approximately 60%
through their incubation period, much younger than previously
hypothesized. The overall consistency of our results with those of
living saurians indicates that they can be generalized to other extinct
members of that lineage, and therefore our method provides an
independent means of assessing the developmental stage of extinct,
in-ovo saurians.
Os embriões de dinossauros estão entre os fósseis mais raros, mas fornecem uma janela única para a paleobiologia desses animais. A estimativa do estágio de desenvolvimento dos embriões de dinossauros é dificultada pela falta de um método quantitativo para determinação da idade, pela escassez de material e pela dificuldade em visualizá-lo. Apresentamos os resultados de uma ampla pesquisa, usando reconstruções 3D de dados de tomografia computadorizada de raios-X, em sequências de ossificação craniana em táxons saurianos existentes e em embriões bem preservados do dinossauro sauropodomorfo, Massospondylus carinatus. Nossas descobertas apóiam a conservação em tempo real de sequências de ossificação craniana em saurianos, incluindo dinossauros, permitindo desenvolver um novo método para estimar a porcentagem relativa de desenvolvimento de embriões desse clado. Também observamos dentes de geração nula nos embriões de Massospondylus carinatus que são reabsorvidos ou eliminados antes da eclosão, semelhantes aos das lagartixas. Essas linhas de evidência nos permitem estimar com segurança que os embriões de Massospondylus carinatus são de apenas aproximadamente 60% durante o período de incubação, muito mais jovens do que se supunha anteriormente. A consistência geral de nossos resultados com os de saurianos vivos indica que eles podem ser generalizados para outros membros extintos dessa linhagem e, portanto, nosso método fornece um meio independente de avaliar o estágio de desenvolvimento de saurianos in-ovo extintos.
Introduction
Birds
are the sole living dinosaur lineage and are therefore often used as a
modern analogue when studying dinosaurian evolution, especially where
fossilized remains are lacking, such as in developing eggs1,2,3,4,5,6.
Embryology has revealed that some non-avian dinosaurs display both
plesiomorphic developmental characteristics (such as a slow incubation
period) as well as derived developmental characteristics of birds (such
as skeletal anatomy and egg macro- and microstructure)7.
Recent research has also shown that the shape of the bird skull can be
explained by paedomorphic retention of juvenile, non-avian dinosaur
features4.
Despite these findings, little is known about the cranial ossification
sequence of dinosaurian embryos, and how this sequence compares to other
saurians8,9.
As aves são a única linhagem de dinossauros vivos e, portanto, são frequentemente usadas como um análogo moderno no estudo da evolução dos dinossauros, especialmente onde faltam restos fossilizados, como no desenvolvimento de ovos1,2,3,4,5,6. A embriologia revelou que alguns dinossauros não aviários exibem características de desenvolvimento plesiomórficas (como um período de incubação lento) e características de desenvolvimento derivadas de aves (como anatomia esquelética e macro e microestrutura de ovo) 7. Pesquisas recentes também mostraram que a forma do crânio do pássaro pode ser explicada pela retenção pedomórfica de características juvenis de dinossauros não aviários4. Apesar desses achados, pouco se sabe sobre a sequência de ossificação craniana de embriões dinossauros e como essa sequência se compara a outros saurianos8,9.
Ossification
patterns during embryonic development have been studied using clearing
and staining for a variety of saurian taxa including: several species of
galliform10,11, palaeognath12, anseriform11, and passeriform birds11; crocodilians13; testudines14,15,16,17,18; and squamates19,20,21. More recently, X-ray micro-computed tomography (µCT) has also been used as a method to look at these patterns22,23.
In birds, the general ossification sequence of skeletal elements in
embryos has been found to be conserved within species and to a certain
degree between species and groups, regardless of altriciality or
precociality24,25 in hatchlings. However, some heterochronies can occur with the relative timing of these ossification events varying slightly11,12.
In non-avian reptiles, there is very slight interspecific and
intraspecific variability in both the pattern and timing of cranial
ossification13,14,18,19,20,21,26.
Understanding these ossification sequences as well as the phylogenetic
relationships between the taxa is important for the clarification of
heterochronic processes in macroevolution.
In extant taxa,
embryonic stages are commonly determined by the appearance of
morphological traits such as somites, cartilaginous processes,
pigmentation, muscles, brain development, etc.15,27,28,29,30.
The level of cranial ossification has been previously mentioned for
some of these stages (for example in crocodiles and some squamates), but
the ossification sequence of the bones has not been explicitly used nor
universally applied as a criterion for assessing developmental stage23,29,30. However, a broader study using event-pair cracking31, found that there are five modules in the cranial ossification sequence across large phylogenetic distances32. These consist of the jaw bones, the palatal bones, the bones forming the orbit, the skull roof bones and the braincase bones.
In
extinct saurians, dinosaurs have the most abundant embryonic record,
with in-ovo fossils having been found across all three major lineages33,34,35,36,37,38,39,40,41.
Most dinosaur embryonic material includes cranial bones. Past studies
have used the degree of cranial sutural closure to estimate the level of
maturity of these dinosaur embryos42,43,44. The underlying assumption of these studies is that, as animals mature, the sutures become narrower and eventually close8,9. This method is somewhat compromised by post-mortem disarticulation and slow growth rates of some reptiles9. The progressive closure of sutures is also not observed in all extant taxa. For example, Alligator mississippiensis has cranial sutures that widen throughout ontogeny, possibly due to feeding mechanics8. Finally, the degree of sutural closure is difficult to assess in very immature specimens such as embryos.
Other
methods of assessing developmental stages in fossil embryos have also
been explored. The size of the embryo in proportion to its egg was used
to infer maturity in an enantiornithine and in Massospondylus carinatus33,34,41. A study of therizinosauroid embryos compared the postcranial patterns of ossification to that of alligators38.
Several studies have compared dinosaur embryonic postcranial and
cranial ossification levels to extant birds in order to determine the
developmental stage (eg. titanosaurs, oviraptorid, troodontids)35,36,39.
Incubation periods are a complicating factor for such studies, as they
vary greatly between living saurians (i.e., birds, crocodilians, and
turtles)7,
making it challenging to determine which clade is the best proxy for
dinosaurs. To our knowledge, there is no study that compares
ossification levels of individual cranial bones in dinosaurs to a broad
sample of saurian embryos.
In 1976, a clutch of seven subspherical
eggs (BP/1/5347a) was discovered by Prof. James Kitching in the early
Jurassic upper Elliot Formation of Golden Gate Highlands National Park,
South Africa34,45,46,47. Two partially exposed embryos in the clutch were quickly identified as being dinosaurian47,
making them among the oldest known dinosaur eggs and embryos in the
world. These eggs were later identified as belonging to the basal
sauropodomorph dinosaur species Massospondylus carinatus34
and the visible embryonic remains were described. The size of the
embryos relative to their respective eggs, along with general
observations about the level of ossification, and the presence of a
stapes and a fourth trochanter lead to the hypothesis that they were
nearing the end of development and close to hatching46.
Here, we take a new look at embryonic cranial ossification patterns in the Massospondylus carinatus
embryos (BP/1/5347a). We reconstruct the ossified portions of their
embryonic skulls, and we compare them to ossified cranial bones in a
growth series of four living saurian taxa (Gallus gallus, Crocodylus niloticus, Centrochelys sulcata and Pogona vitticeps) using synchrotron radiation X-ray micro-computed tomography (SRµCT) imaging methods48 and published literature23.
We develop a numerical method for coding the ossification stage of each
bone in each specimen, and use a dissimilarity matrix to assess the
relative developmental percentage of the Massospondylus carinatus embryos.
Methods
The
seven eggs preserved in the clutch BP/1/5347A were characterised at the
ID19 beamline of the European Synchrotron Radiation Facility (ESRF,
Grenoble, France) using propagation phase contrast SRµCT. Each egg was
first imaged individually using a setup providing an isotropic voxel
size of 13.11µm. In a second experiment, we focused on the two visible
embryonic skulls and increased the resolution using a setup providing an
isotropic voxel size of 2.98µm (details of both setups are provided in
S1). The bones were digitally reconstructed in VG Studio MAX 3.2 (Volume
Graphics, Heidelberg, Germany). The best exemplars of each bone in the Massospondylus carinatus
embryos were then extracted as surface mesh (.stl) files and combined
in order to reconstruct an articulated skull for visualization purposes
(Fig. 1). Surface files are available on the online repository Morphosource (https://www.morphosource.org/MyProjects/Dashboard/dashboard/select_project_id/798).
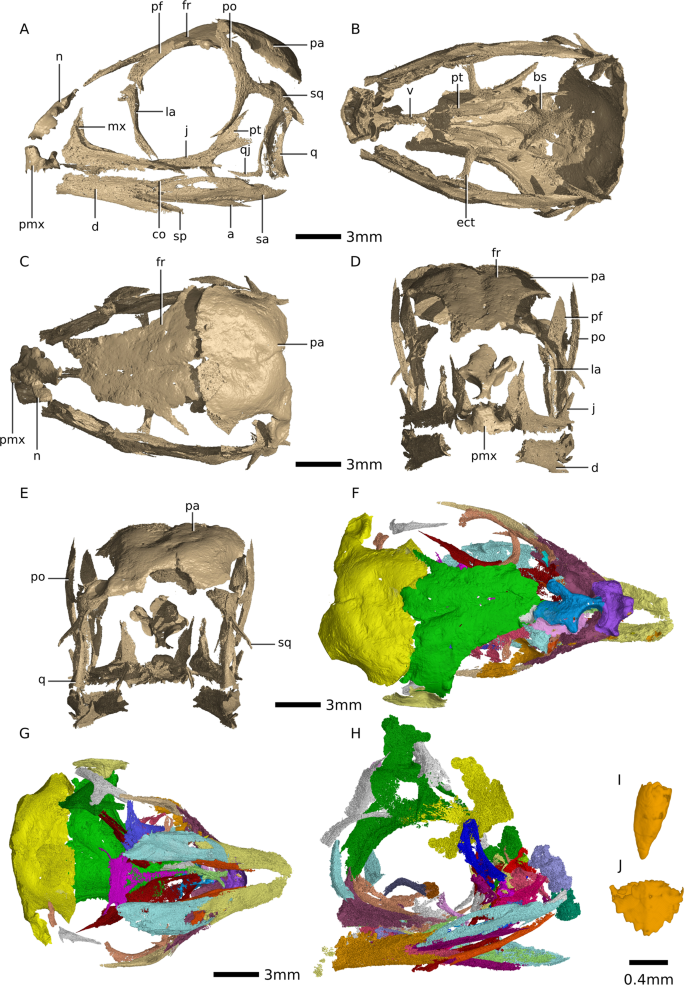
Figure 1
Digital reconstructions of Massospondylus carinatus embryonic skulls based on SRµCT data. (A) composite of Massospondylus carinatus embryonic skulls in left lateral view. (B) composite of Massospondylus carinatus embryonic skulls in ventral view. (C) composite of Massospondylus carinatus embryonic skulls in dorsal view. (D) composite of Massospondylus carinatus embryonic skulls in anterior view. (E) composite of Massospondylus carinatus embryonic skulls in posterior view. (F) Massospondylus carinatus Embryo 2 skull reconstruction as preserved in dorsal view. (G) Massospondylus carinatus Embryo 2 skull reconstruction as preserved in ventral view. (H) Massospondylus carinatus Embryo 1 skull reconstruction as preserved in lateral view. (I) Massospondylus carinatus embryonic null-generation tooth. (J) Massospondylus carinatus
functional embryonic tooth. Abbreviations: a, angular; bs,
basisphenoid; d, dentary; ect, ectopterygoid; fr, frontal; j, jugal; la,
lacrimal; mx, maxilla; n, nasal; pa, parietal; pf, prefrontal; po,
postorbital; pmx, premaxilla; pt, pterygoid; sa, surangular; sp,
splenial; sq, squamosal; q, quadrate; qj, quadratojugal; v, vomer.
Tomographical data of Centrochelys sulcata, Gallus gallus and Crocodylus niloticus embryos were obtained from the ESRF database (http://paleo.esrf.eu; acquisition parameters provided in Table S2).
Each sampled developmental percentage in that
dataset (i.e., approximately x% through the incubation period, depending
on taxon and specimen availability) was reconstructed for each taxon.
Data for Pogona vitticeps was obtained from literature23
that used µCT scans. It is important to note that this taxon was
analysed using the illustrations provided to test our method on
published data, that it does not include all developmental percentages
in the incubation period, and therefore that it needs to be considered
with caution.
The degree of ossification of individual bones was
used as an indicator of developmental percentage. We quantified our
observations of ossification level across taxa and ontogenetic stages
using the following numerical system: stage-code 0 = absence of
ossification in that bone; stage-code 1 = beginning of ossification,
usually in the form of a small amorphous pellet; stage-code 2 = bone is
immediately recognizable in shape, but many of its processes and rami
are incompletely ossified, often ending in somewhat ragged projections;
and stage-code 3 = bone shape strongly resembles the juvenile condition
short of complete expansion (see Table S3).
Bones
that are “lost” in some taxa, either through fusion to other bones or
lack of complete ossification, were coded as NA in our matrix, although
these may have embryonic ossification centres (e.g., the postorbital and
prefrontal of Gallus gallus49).
It was not possible to fully track the level of ossification of these
(i.e., as stage-code 2 or 3) across the full embryonic sequence.
In two of the Massospondylus carinatus
embryos, the parietals, frontals and nasals are extremely thin sheets
of bone preserved on the surface of the specimen (i.e., they have been
freed from the encasing matrix by preparation). The phase retrieval
algorithm used for the tomographic reconstruction often obscures the
margins of bones located at the surface, making it difficult to
digitally reconstruct them (see Fig. 1).
They can, however, clearly be seen on the specimen and we have coded
them in our ossification level scheme with the aid of visual inspections
under a microscope (in this case, as a level 2). As an additional
sensitivity analysis, we also coded them as a level 3 and present the
results in the supplement (see Table S4).
A
matrix was created with each row representing a different relative
developmental percentage (days in incubation) of each taxon during
incubation (i.e., between the start and end of embryonic ossification,
see Table S3) and
each column representing a different cranial bone. A dissimilarity
distance matrix containing all pairwise comparisons of developmental
percentages across all comparable bones using the “Gower” method was
computed using R Studio Max v1.1.45350 and the package vegan51.
The “Gower” method was selected as it corrects for missing data (in
this case bones which are not present across all taxa) as NAs are not
considered when calculating distances52.
The matrix was used to quantitatively compare dissimilarity between
individual stages in extant saurian ontogenetic sequences and the
ossification pattern observed in the Massospondylus carinatus embryos (see Tables 1 and S5
for distance matrix results using S3, and see S6 for sensitivity
analysis distance matrix results using S4). The distance matrix (Table S5) was then subjected to a principal coordinate analysis (PCoA) using the R package ape53 (see Table S7 and Fig. S8).
Table 1 Dissimilarity distance matrix pairwise comparisons of Massospondylus carinatus
embryo terminals using the “Gower” method. Specimens in bold represent
the shortest distance between Massospondylus carinatus embryos and the
extant taxon in question.
As an additional sensitivity analysis, the stage-code matrix (Table S5) was replaced by a simple presence|absence matrix (binary 0 and 1 stage-codes, see Table S9), and a dissimilarity matrix using the same parameters as above was generated (see Table S10).
Results
Phase contrast was successful in detecting embryonic remains in BP/1/5347a, as in other studies of fossilized embryonic remains48.
Our analyses of the clutch using this method revealed skeletal material
in only three of the eggs: in the fully prepared embryo in lateral view
(here considered to be the bottom left egg), in the prepared embryo in
dorsal view, and in the broken egg in the top right corner of the clutch
(which included a few cranial bones). Throughout the manuscript,
“Embryo 1” will refer to the embryo prepared in lateral view, “Embryo 2”
will refer to the embryo prepared in dorsal view and “Embryo 3” will
refer to the partial embryo in the top right egg (see Fig. S11).
Embryo
1 is mostly articulated but is missing the anterior tip of its snout.
Among its cranial bones, it preserves the maxilla, nasal, jugal,
lacrimal, frontal, parietal, postorbital, quadrate, quadratojugal,
pterygoid, ectopterygoid, palatine, vomer, basisphenoid, angular,
surangular, prearticular, splenial, coronoid and dentary (see Fig. 1 and Table S12).
Embryo
2 is mostly articulated and among its cranial bones preserves the
premaxilla (that appears to have been damaged during preparation),
maxilla, nasal, jugal, lacrimal, frontal, parietal, postorbital,
quadratojugal, pterygoid, ectopterygoid, palatine, vomer, basisphenoid,
angular, surangular, prearticular, splenial, coronoid and dentary (see
Fig. 1 and Table S12).
Embryo 3 is disarticulated and among its cranial bones only preserves a right maxilla, right postorbital and frontal (see Fig. S13 and Table S12).
In
general, the bones that are approximately fully ossified (stage-code 3)
in all of the embryos are the bones of the snout (maxilla, premaxilla),
the mandible (dentary, coronoid, splenial, surangular, angular,
prearticular), the postorbital, prefrontal, quadratojugal, squamosal, as
well as some of the bones of the palate (pterygoid, ectopterygoid). The
bones of the skull roof (frontal, parietal, nasals) are recognizable in
shape but have incompletely ossified margins (stage-code 2), as are the
palatine, the vomers, jugals, quadrates, and the lacrimals, in which
the full shapes have not been realized.
The only ossified
braincase bone in our sample is the basisphenoid (stage-code 2). In
Embryo 1, it is a flat sheet of bone with an anteriorly extending
cultriform process and a tube-like, partially ossified right
basipterygoid process. The left basipterygoid process as well as the
basal tubera have not yet ossified. The basisphenoid of Embryo 2 is a
flat sheet of bone with a cultriform process but no ossification of the
basal tubera or basipterygoid processes. The quadrate is partially
ossified in Embryo 1 (portions of the midshaft and proximal portions of
the pterygoid and quadratojugal rami have ossified) but is absent in
Embryo 2.
All three embryos have teeth in the maxillae, dentaries
and premaxilla (where preserved). The teeth have two distinct
morphologies: small, simple and conical; and large, broad, and serrated
(see Fig. 1).
The latter morphology only has crowns or partial crowns formed with no
roots. These two morphologies do not appear in any identifiable pattern
and can sometimes be found in the same alveolus or in adjacent alveoli
(see Fig. S14). Crocodylus niloticus, Gallus gallus domesticus, Centrochelys sulcata and Pogona vitticeps have similar ossification sequences to each other (Tables S3 andS5).
The bones of the snout are first to ossify, including the premaxilla,
maxilla and dentary. They are followed by the remaining bones of the
mandible, excluding the articular, and most of the lateral bones of the
face (except for the quadrate and parietal) and the palatal bones
(pterygoid, ectopterygoid when present, palatine and vomers). The
coronoid (when present) ossifies next, along with the supraoccipital,
otoccipital, quadrate, and the basisphenoid. This is also when teeth
begin to form (when present). Following this, the basioccipital and
prootic begin to ossify, followed by the parietal and laterosphenoid
(when present). Finally, the articular and the palpebral (when present)
are the last bones to begin ossifying at approximately 70% through the
incubation period.
The dissimilarity matrix (see Tables 1 and S5) indicates that the Massospondylus carinatus embryos are most similar to the Crocodylus niloticus embryos at 48 days, the Gallus gallus embryos at 14 days, the Centrochelys sulcata embryos at 61 days of the incubation, and the Pogona vitticeps embryos at 48 days of the incubation (see Fig. 2). The distance between the Massospondylus carinatus embryos and Pogona vitticeps
at 48 days is relatively large compared to the others, however this
comparison is of lower resolution due to the gap in developmental
percentages included in the analysis (no Pogona vitticeps embryos between 55% and 74% through the incubation period were illustrated in the published study).
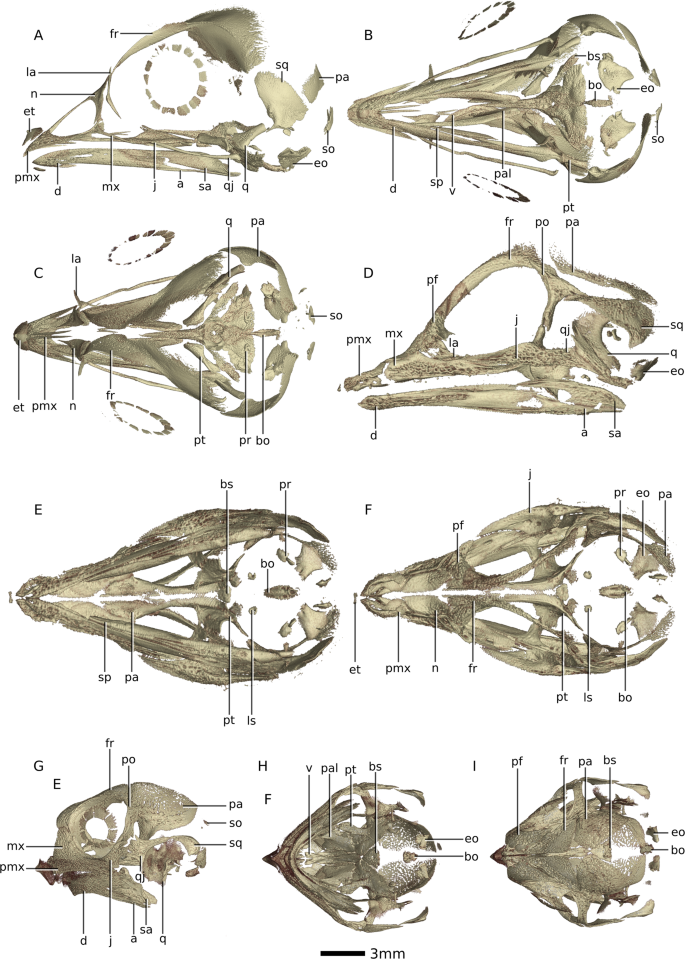
Figure 2
Digital reconstructions of saurian embryo skulls based on SRµCT data. (A) Gallus gallus at 14 days in incubation period in left lateral view. (B) Gallus gallus at 14 days in incubation period in ventral view. (C) Gallus gallus at 14 days in incubation period in dorsal view. (D) Crocodylus niloticus at 48 days in incubation period in left lateral view. (E) Crocodylus niloticus at 48 days in incubation period in ventral view. (F) Crocodylus niloticus at 48 days in incubation period in dorsal view. (G) Centrochelys sulcata at 61 days in incubation period in left lateral view. (H) Centrochelys sulcata at 61 days in incubation period in ventral lateral view. (I) Centrochelys sulcata
at 61 days in incubation period in dorsal lateral view. Abbreviations:
a, angular; bo, basioccipital; bs, basisphenoid; d, dentary; eo,
exoccipital; et, egg tooth; fr, frontal; j, jugal; la, lacrimal; ls,
laterosphenoid; mx, maxilla; n, nasal; pa, parietal; pal, palatine; pf,
prefrontal; po, postorbital; pmx, premaxilla; pr, prootic; pt,
pterygoid; sa, surangular; so, supraoccipital; sp, splenial; sq,
squamosal; q, quadrate; qj, quadratojugal; v, vomer.
Changing the stage-codes of the Massospondylus carinatus embryo frontals and parietals to stage-code 3 does not alter the general results of the dissimilarity matrix (see Table S6). The distance values change, but the smallest distances between Massospondylus carinatus
and the respective extant taxa remain at the same developmental
percentages. The sensitivity analysis using a simple binary
presence|absence matrix cannot estimate a developmental percentage for
the Massospondylus carinatus embryos because too many of the distances between developmental percentages are equal (see Table S10).
Discussion
Our
results show that both the sequence and to a certain extent, the
relative timing of cranial ossification are conserved between Gallus gallus, Crocodylus niloticus, Centrochelys sulcata and Pogona vitticeps. This sequence agrees with previous work using different methods done on larger phylogenetic samples (including mammals)31,32,54. In general, this order follows the aforementioned modules described in the event-pair cracking study32
(i.e. jaw bones, the palatal bones, the bones forming the orbit, the
skull roof bones and the braincase bones), however some bones ossify
later than the rest of their respective modules (such as the coronoid,
the laterosphenoid, the articular and the palpebral).
Based on the
level of ossification of the basisphenoid, frontals, parietals, palate,
and quadrate, as well as the absence of the remaining braincase bones
and the articular, we hypothesize that the Massospondylus carinatus embryos are approximately 60% through their incubation period (56% or 48 days out of 85 for crocodiles55;
67% or 12 days out of 21 for chickens; 61% or 61 days out of 100 for
spurred tortoises and 74% or 48 days out of 65 for bearded dragons56) (see Fig. 2).
In our analysis, the bearded dragon was not scored between 36 days and
48 days in the incubation period (or 55–74%), as this gap was not
illustrated in the literature. This comparison therefore has lower
precision than the others and is considered with caution. Although at
this relative age we would expect the Massospondylus carinatus
embryos to have an onset of ossification in the rest of the braincase
bones (i.e notably exoccipital and basioccipital but also possibly the
supraoccipital, prootic and laterosphenoid), the extant taxa present a
small ossification centre for these bones at this developmental
percentage. It is therefore possible that these had not started to
ossify yet in the dinosaur embryos, that the small ossification centres
did not preserve, or that they were below the resolution threshold in
our phase -contrast -based SRµCT data. This indicates that the embryos
are certainly not much more developed than 60% through their incubation
period, as these braincase bones would otherwise be more ossified and
visible. In Gallus gallus, Crocodylus niloticus,Centrochelys sulcata, and
Pogona vitticeps, these braincase bones are all at stage-code 2 at
approximately 70–75% through the incubation period (day 15, day 55, day
70 and day 60 respectively, see Table S3). This puts an upper limit on the developmental percentage of the Massospondylus carinatus embryos because stage-code 2 ossification is readily seen in our scan in other bones of the skull.
Our hypothesized relative developmental percentage of the Massospondylus carinatus embryos indicates that they are earlier in development than previously thought46.
This makes them some of the ontogenetically youngest dinosaur embryos
known. All other dinosaur embryos in the literature with ontogenetic age
estimates are hypothesized as being in the last third of their
development or near hatching35,36,37,38,39.
However, analysing the latter using micro computed tomography scans and
our stage-code method could reveal that some of them are younger than
the Massospondylus carinatus embryos presented here.
Our hypothesis for the developmental percentage of the Massospondylus carinatus embryos is corroborated by the presence of both null-generation teeth and crowns with adult tooth morphology (see Fig. 1). Although many dinosaur embryos have been found to have teeth36,37,38,40,
in all cases the reported morphology is most similar to adult teeth. To
our knowledge, null-generation teeth have not been reported in
dinosaurs, nor have null-generation and adult teeth been reported in a
single embryo. Troodon teeth have cylindrical roots,
linguobuccaly compressed and mesiodistally elongated crowns. These were
hypothesized as being early developmental casts of unossified teeth35. Maiasaura
embryos preserve different generations of teeth, however the budding
teeth are hypothesized to grow in the form of the larger teeth
preserved. These are therefore replacement teeth and are similar in
morphology57.
Null-generation teeth form during embryonic development in several reptile species58,59,60.
These non-functional teeth are small, unicuspid (even if adult tooth
morphology is multicuspid), and possess little or no enamel. They are
either resorbed into the jaw or shed into the oral cavity59.
Little information is available on null-generation teeth in living
saurian taxa. However, in geckos, the null-generation teeth appear at
about 23% through the incubation period. By 66% into the incubation
period, null-generation teeth are present in half of the tooth
positions, while the other half are formed by the first functional teeth
(null-generation teeth are therefore not a prerequisite for the
formation of adult tooth morphologies). By the end of the incubation
period, the first set of functional teeth have started to be resorbed
while the second set of functional teeth start moving orally58. All null-generation teeth are replaced by functional teeth during embryonic development61. The simple conical tooth morphology seen in the Massospondylus carinatus embryos probably represent null-generation teeth for Massospondylus carinatus,
which will get resorbed or shed into the oral cavity before hatching.
The abundance of these null-generation teeth (approximately half of the
teeth preserved, see Fig. S14) correlate well with the 60% in incubation period estimated by the bone ossification.
Our
stage-code method provides a simple, relatively precise, repeatable
estimate of the developmental percentage in other extinct and extant
saurian embryos. Such estimates are of broad utility when including
embryos in broader ontogenetic studies. For example, the early
developmental percentage of the Massospondylus carinatus embryos
suggests caution against uncritical use of limb measurements as part of
allometric studies, e.g., postural determination. Coupling these
developmental percentage estimates with incubation period7
also provides better insight into the life histories of dinosaurs and
could potentially enable more precise future studies of how birds, for
example, decreased their incubation periods.
Although Reisz et al.
(2010) identified embyronic skeletal material in five of the eggs that
comprise BP/1/5347a, our results show that only three of the seven eggs
contain embryonic material. Several factors have been hypothesized to
have an effect on clutch viability in extant taxa including
environmental conditions, eggshell structure, predation and microbial
contamination62,63. However the patterns of embryonic mortality are poorly understood63.
There are many potential reasons for fossil eggs being empty,
including: a high number of infertile eggs; high levels of early
mortality before ossification of the skeleton; leakage from broken eggs
during fossilisation; or that the clutch represents two or more clutches
laid at different times (Deeming, pers. com.). Given our sample size,
we cannot assess any of these critically at this time. However, the
three preserved embryos do not differ substantially in their
ossification, indicating that they are therefore at similar
developmental percentages and are probably from the same clutch. The
first and second embryos show some slight differences in level of
ossification. The Embryo 1 has a partial basipterygoid process that has
started to ossify as well as a quadrate. Embryo 2 does not have either
of these elements ossified. Despite these, they are highly synchronous
in their development and it is not possible to speculate as to the
nature of the depositional time in the nest, if these embryos would have
asynchronous hatching, or if these differences represent intraspecific
variation in the timing or rate of embryonic ossification11.
A
strong, highly conserved pattern of cranial ossification is seen in
saurians. Both this pattern and the level of ossification at the
different developmental percentages are enough to correlate and
approximate the Massospondylus carinatus embryos. However,
several complicating factors need to be taken into account with regards
to the comparative extant embryonic datasets. Because X-ray µCT imaging
of extant embryos can involve sacrificing the specimens, the series we
present for these taxa do not track the ossification sequence of one
single individual, but rather of several individuals (one for each
developmental percentage represented). Previous research has found that
there is slight inter-individual variations in the timing of
ossification11.
However, given the strength of the overall pattern, we do not consider
the variance introduced by these minor differences to compromise our
overarching result. The datasets for Crocodylus niloticus, Centrochelys sulcata and Pogona vitticeps
do not include individuals for every day in the incubation period, but
rather individuals which are several days apart (especially in Pogona vitticeps).
This reduces precision for tracking the ossification sequence,
complicates understanding ossification rates in individual bones, and
represents a fertile area for further study.
Conclusion
Cranial
ossification sequence is conserved in saurians, including in dinosaurs,
even across large phylogenetic distances. Using this observation and a
stage-code based method recording the ossification level of each cranial
bone at each developmental percentage during the incubation allows for
the determination of the developmental percentage of fossil embryos.
Based on 3D reconstructions from SRµCT data, we find that the famous
embryos of Massospondylus carinatus (BP/1/5347a) are
approximately 60% through their incubation period, much earlier than
previously hypothesized. This is corroborated by the presence of
formerly unreported null-generation teeth. This research provides
potential to elucidate broader patterns of macroevolution and suggests
that heterochronic shifts in ossification timing are likely not a major
mechanism for how different shaped skulls evolved.
Allen,
V., Bates, K. T., Li, Z. & Hutchinson, J. R. Linking the evolution
of body shape and locomotor biomechanics in bird-line archosaurs. Nature497, 104 (2013).
Heers,
A. M., Dial, K. P. & Tobalske, B. W. From baby birds to feathered
dinosaurs: incipient wings and the evolution of flight. Paleobiology40, 459–476 (2014).
Nudds, R. L. & Dyke, G. J. Forelimb posture in dinosaurs and the evolution of the avian flapping flight‐stroke. Evolution: International Journal of Organic Evolution63, 994–1002 (2009).
Bhullar, B. A. et al.
A molecular mechanism for the origin of a key evolutionary innovation,
the bird beak and palate, revealed by an integrative approach to major
transitions in vertebrate history. Evolution; international journal of organic evolution69, 1665–1677, https://doi.org/10.1111/evo.12684 (2015).
Hutchinson,
J. R., Bates, K. T., Molnar, J., Allen, V. & Makovicky, P. J. A
computational analysis of limb and body dimensions in Tyrannosaurus rex with implications for locomotion, ontogeny, and growth. PLoS One6, e26037 (2011).
Erickson,
G. M., Zelenitsky, D. K., Kay, D. I. & Norell, M. A. Dinosaur
incubation periods directly determined from growth-line counts in
embryonic teeth show reptilian-grade development. Proceedings of the National Academy of Sciences114, 540–545 (2017).
Bailleul,
A. M., Scannella, J. B., Horner, J. R. & Evans, D. C. Fusion
patterns in the skulls of modern archosaurs reveal that sutures are
ambiguous maturity indicators for the Dinosauria. PLoS One11, e0147687 (2016).
Sampson,
S. D., Ryan, M. J. & Tanke, D. H. Craniofacial ontogeny in
centrosaurine dinosaurs (Ornithischia: Ceratopsidae): taxonomic and
behavioral implications. Zoological Journal of the Linnean Society121, 293–337 (1997).
Maxwell, E. E. Comparative embryonic development of the skeleton of the domestic turkey (Meleagris gallopavo) and other galliform birds. Zoology111, 242–257 (2008).
Mitgutsch,
C., Wimmer, C., Sánchez-Villagra, M. R., Hahnloser, R. & Schneider,
R. A. Timing of ossification in duck, quail, and zebra finch:
intraspecific variation, heterochronies, and life history evolution. Zoological science28, 491 (2011).
Maxwell, E. E. Comparative ossification and development of the skull in palaeognathous birds (Aves: Palaeognathae). Zoological Journal of the Linnean Society156, 184–200 (2009).
Rieppel, O. Studies on skeleton formation in reptiles. V. Patterns of ossification in the skeleton of Alligator mississippiensis Daudin (Reptilia, Crocodylia). Zoological Journal of the Linnean Society109, 301–325 (1993).
Rieppel, O. Studies on skeleton formation in reptiles: patterns of ossification in the skeleton of Chelydra serpentina (Reptilia, Testudines). Journal of Zoology231, 487–509 (1993).
Greenbaum, E. & Carr, J. L. Staging criteria for embryos of the spiny softshell turtle, Apalone spinifera (Testudines: Trionychidae). Journal of morphology254, 272–291 (2002).
Tulenko, F. J. & Sheil, C. A. Formation of the chondrocranium of Trachemys scripta (Reptilia: Testudines: Emydidae) and a comparison with other described turtle taxa. Journal of morphology268, 127–151 (2007).
Danielson, S. C. & Sheil, C. A. Patterns of chondrification and ossification in the skull of Graptemys pseudogeographica, the false map turtle (Emydidae). Journal of morphology278, 1739–1753 (2017).
Rieppel,
O. Studies on skeleton formation in reptiles. III. Patterns of
ossification in the skeleton of Lacerta vivipara Jacquin (Reptilia,
Squamata). Fieldiana (Zool.) N. S68, 1–25 (1992).
Rieppel, O. Studies on skeleton formation in reptiles. II. Chamaeleo hoehnelii (Squamata: Chamaeleoninae), with comments on the homology of carpal and tarsal bones. Herpetologica, 66–78 (1993).
21.
Rieppel, O. Studies on skeleton formation in reptiles. Patterns of ossification in the skeleton of Lacerta agilis exigua Eichwald (Reptilia, Squamata). Journal of Herpetology, 145–153 (1994).
Werneburg, I., Polachowski, K. M. & Hutchinson, M. N. Bony skull development in the Argus monitor (Squamata, Varanidae, Varanus panoptes) with comments on developmental timing and adult anatomy. Zoology118, 255–280 (2015).
Ollonen,
J., Da Silva, F. O., Mahlow, K. & Di-Poï, N. Skull development,
ossification pattern, and adult shape in the emerging lizard model
organism Pogona vitticeps: a comparative analysis with other squamates. Frontiers in physiology9, 278 (2018).
Starck,
J. M. Quantitative design of the skeleton in bird hatchlings: does
tissue compartmentalization limit posthatching growth rates? Journal of morphology222, 113–131 (1994).
Maxwell,
E. E., Harrison, L. B. & Larsson, H. C. Assessing the phylogenetic
utility of sequence heterochrony: evolution of avian ossification
sequences as a case study. Zoology113, 57–66 (2010).
Rieppel, O. Studies on skeleton formation in reptiles. I. The postembryonic development of the skeleton in Cyrtodactylus pubisulcus (Reptilia: Gekkonidae). Journal of Zoology227, 87–100 (1992).
Vieira, L. et al. Description of embryonic stages in Melanosuchus niger (Spix, 1825)(Crocodylia: Alligatoridae). Journal of Morphological Science28, 11–22 (2011).
Magnusson, W. E. & Taylor, J. A. A description of developmental stages in Crocodylus porosus, for use in aging eggs in the field. Wildlife Research7, 479–485 (1980).
Jeffery,
J. E., Richardson, M. K., Coates, M. I. & Bininda-Emonds, O. R.
Analyzing developmental sequences within a phylogenetic framework. Systematic biology51, 478–491 (2002).
Balanoff,
A. M., Norell, M. A., Grellet-Tinner, G. & Lewin, M. R. Digital
preparation of a probable neoceratopsian preserved within an egg, with
comments on microstructural anatomy of ornithischian eggshells. Naturwissenschaften95, 493–500 (2008).
Reisz,
R. R., Scott, D., Sues, H.-D., Evans, D. C. & Raath, M. A. Embryos
of an Early Jurassic prosauropod dinosaur and their evolutionary
significance. Science309, 761–764 (2005).
Varricchio, D. J., Horner, J. R. & Jackson, F. D. Embryos and eggs for the Cretaceous theropod dinosaur Troodon formosus. Journal of Vertebrate Paleontology22, 564–576 (2002).
Garcia,
R. A. & Cerda, I. A. Dentition and histology in titanosaurian
dinosaur embryos from Upper Cretaceous of Patagonia, Argentina. Palaeontology53, 335–346 (2010).
Horner, J. R. & Currie, P. J. In Dinosaur eggs and babies (eds. Kenneth Carpenter, Hirsch, K. F., & Horner, J. R.) 312–337 (Cambridge University Press, 1994).
38.
Kundrát,
M., Cruickshank, A. R., Manning, T. W. & Nudds, J. Embryos of
therizinosauroid theropods from the Upper Cretaceous of China: diagnosis
and analysis of ossification patterns. Acta Zoologica89, 231–251 (2008).
Norell,
M. A., Clark, J. M. & Chiappe, L. M. An embryonic oviraptorid
(Dinosauria: Theropoda) from the Upper Cretaceous of Mongolia. American Museum Novitates3315, 1–17 (2001).
Chiappe, L. M., Salgado, L. & Coria, R. A. Embryonic skulls of titanosaur sauropod dinosaurs. Science293, 2444–2446, https://doi.org/10.1126/science.1063723 (2001).
Varricchio,
D. J., Balanoff, A. M. & Norell, M. A. Reidentification of avian
embryonic remains from the cretaceous of Mongolia. PLoS One10, e0128458, https://doi.org/10.1371/journal.pone.0128458 (2015).
Longrich, N. R. & Field, D. J. Torosaurus is not Triceratops: ontogeny in chasmosaurine ceratopsids as a case study in dinosaur taxonomy. PloS one7, e32623 (2012).
Rauhut, O. W. M. Braincase structure of the Middle Jurassic theropod dinosaur Piatnitzkysaurus. Canadian Journal of Earth Sciences41, 1109–1122 (2004).
Reisz,
R. R., Evans, D. C., Roberts, E. M., Sues, H. D. & Yates, A. M.
Oldest known dinosaurian nesting site and reproductive biology of the
Early Jurassic sauropodomorph Massospondylus. Proceedings of the National Academy of Sciences of the United States of America109, 2428–2433, https://doi.org/10.1073/pnas.1109385109 (2012).
Reisz, R. R., Evans, D. C., Sues, H.-D. & Scott, D. Embryonic skeletal anatomy of the sauropodomorph dinosaur Massospondylus from the Lower Jurassic of South Africa. Journal of Vertebrate Paleontology30, 1653–1665 (2010).
Kitching,
J. W. Preliminary report on a clutch of six dinosaurian eggs from the
Upper Triassic Elliot Formation, Northern Orange Free State. Palaeontologia africana22, 41–45 (1979).
Fernandez, V. et al. Phase contrast synchrotron microtomography: Improving noninvasive investigations of fossil embryos in ovo. Microscopy and microanalysis18, 179–185 (2012).
Smith-Paredes, D. et al. Dinosaur ossification centres in embryonic birds uncover developmental evolution of the skull. Nature ecology & evolution2, 1966 (2018).
Sánchez‐Villagra,
M. R., Goswami, A., Weisbecker, V., Mock, O. & Kuratani, S.
Conserved relative timing of cranial ossification patterns in early
mammalian evolution. Evolution & development10, 519–530 (2008).
Fergusson, R. A. Nile crocodile Crocodylus niloticus. Crocodiles. Status Survey and Conservation Action Plan, 3rd edn (eds. Manolis, S. C. & Stevenson, C.), 84–89 (2010).
56.
Boyer, T. H. Diseases of Bearded Dragons. In Proceedings of the Pacific Veterinary Conference (2015).
57.
Horner, J. R. Cranial osteology and morphology of the type specimen of Maiasaura peeblesorum (Ornithischia: Hadrosauridae), with a discussion of its phylogenetic position. Journal of Vertebrate Paleontology3, 29–38 (1983).
Zahradnicek,
O., Horacek, I. & Tucker, A. S. Tooth development in a model
reptile: functional and null generation teeth in the gecko Paroedura picta. Journal of anatomy221, 195–208 (2012).
Westergaard, B. & Ferguson, M. W. J. Development of the dentition in Alligator mississipiensis:
upper jaw dental and craniofacial development in embryos, hatchlings,
and young juveniles, with a comparison to lower jaw development. The American Journal of Anatomy187, 393–421 (1990).
Westergaard, B. & Ferguson, M. Development of the dentition in Alligator mississippiensis. Later development in the lower jaws of embryos, hatchlings and young juveniles. Journal of Zoology212, 191–222 (1987).
Sire,
J. Y., Davit‐Beal, T., Delgado, S., Van Der Heyden, C. & Huysseune,
A. First‐generation teeth in nonmammalian lineages: evidence for a
conserved ancestral character? Microscopy research and technique59, 408–434 (2002).
Cedillo-Leal, C. et al. Eggshell structure in Caiman latirostris eggs improves embryo survival during nest inundation. Proceedings of the Royal Society B: Biological Sciences284, 20162675 (2017).
Deeming, D. & Wadland, D. Observations on the patterns of embryonic mortality over the laying season of pheasants. British poultry science42, 569–573 (2001).
We
acknowledge the European Synchrotron Radiation Facility for provision
of synchrotron radiation facilities and we would like to thank Paul
Tafforeau for assistance in using beamline ID19. We thank Céline for her
kind hospitality during data collection. We thank Josef Stiegler for
interesting input. Funding for this research was provided by the DST-NRF
African Origins Platform grants (98800 and 118794 to JNC), the DST-NRF
Centre of Excelle
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.