An early dispersal of modern humans from Africa to Greece
Analysis of two fossils from a Greek cave has shed
light on early hominins in Eurasia. One fossil is the earliest known
specimen of Homo sapiens found outside Africa; the other is a Neanderthal who lived 40,000 years later.
The origin and early dispersal of Homo sapiens has long been a subject of both popular and scholarly interest1. It is almost universally agreed that H. sapiens
(modern humans) evolved in Africa, with the earliest known fossil
representatives of our species dated to around 315,000 years ago in
Morocco (at a site called Jebel Irhoud)2 and approximately 260,000 years ago in South Africa (at Florisbad)3. Stone tools comparable to those found with both of these fossils have been excavated in Kenya (at Olorgesailie)4 and dated to about 320,000 years ago. Writing in Nature, Harvati et al.5 describe their analysis of a fossil from Apidima Cave in southern Greece that they report to be an early modern H. sapiens
at least 210,000 years old. This fossil is the oldest known modern
human in Europe, and probably in all of Eurasia, and is more than
160,000 years older than the next oldest known European fossil of H. sapiens6.
The Apidima Cave complex was excavated in the late 1970s.
Two partial crania (skulls without the lower jaw), named Apidima 1 and
Apidima 2, were recovered in a single block of a type of rock called
breccia. Neither fossil was previously described in detail. Apidima 2
includes the facial region of the skull and had been identified as a
Neanderthal7.
Apidima 1 consists of only the back of the skull and had not been
previously allocated definitively to a species. Harvati and colleagues
used computed tomography to scan the fossils, and generated a 3D virtual
reconstruction of each specimen. They analysed each fossil to assess
aspects of its shape, and thus to determine the fossils’ similarity to
those of other species.
Apidima 2 is badly damaged owing to
previous breakage and distortion. Analyses of all four generated
reconstructions of the fossil were consistent with it being an early
Neanderthal. Apidima 1 is also damaged, but the specimen is not too
badly distorted, so mirroring its right and left sides yielded a good
reconstruction. The authors’ extensive comparative analysis indicates
that this fossil is an early member of H. sapiens. The posterior part of the cranium is rounded like that of H. sapiens,
and it lacks classic Neanderthal features, such as the distinctive
occipital ‘chignon’ — a bulge at the back of the skull that is shaped
like hair tied in a bun.
Earlier dating8
of a fragment of Apidima 2 using a method called uranium-series
analysis indicated a minimum age of around 160,000 years. Harvati and
colleagues report a more extensive set of uranium-series dating
analyses, which surprisingly reveal that Apidima 1 and Apidima 2 are of
different ages, even though they were found in close proximity. Apidima 2
is around 170,000 years old — well within the age range of other
Neanderthal fossils found across Europe (Fig. 1). Apidima 1 is dated to
be at least 210,000 years old, which is much older than any other widely
accepted H. sapiens fossils found outside Africa.
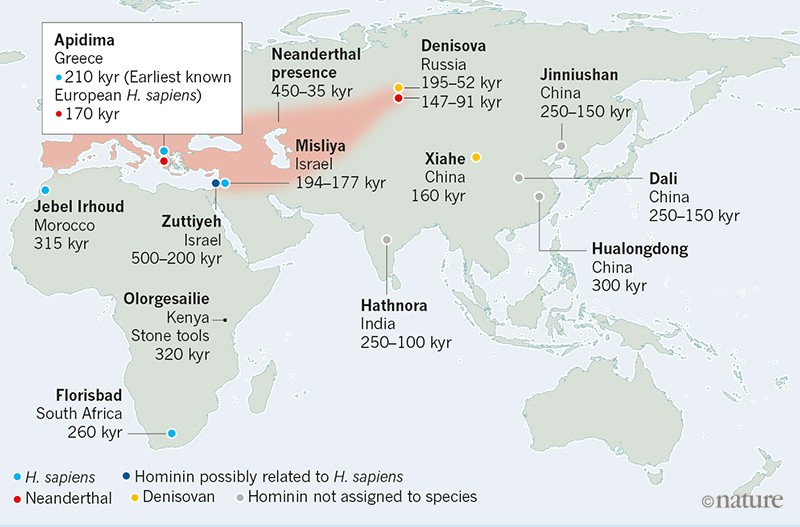
Figure 1 | Some key early fossils of Homo sapiens and related species in Africa and Eurasia. Harvati et al.5 present their analyses of two fossil skulls from Apidima Cave in Greece. They report that the fossil Apidima 1 is an H. sapiens
specimen that is at least 210,000 years old, from a time when
Neanderthals occupied many European sites. It is the earliest known
example of H. sapiens in Europe, and is at least 160,000 years older than the next oldest H. sapiens fossils found in Europe6 (not shown). Harvati and colleagues confirm that, as previously reported7,
Apidima 2 is a Neanderthal specimen, and they estimate that it is at
least 170,000 years old. The authors’ findings, along with other
discoveries of which a selection is shown here, shed light on the timing
and locations of early successful and failed dispersals out of Africa
of hominins (modern humans and other human relatives, such as
Neanderthals and Denisovans). kyr, thousand years old.
This finding reveals that at least two species of hominin
(humans and human relatives from the branch of the family tree after our
split from chimpanzees) inhabited southeastern Europe approximately
200,000 years ago. The discovery of an H. sapiens fossil in Apidima raises questions about what happened to this population. Given that this H. sapiens
existed at a time when there is substantial evidence for a Neanderthal
presence at other European sites, was it part of a population that was
unable to compete successfully with Neanderthals, especially in the
unstable climate of that time? Perhaps one or more times, the two
species replaced each other as the main hominin group present in this
region.
Such patterns of replacement characterize the distribution
of modern humans and Neanderthals in the Levant region of the Middle
East between 250,000 and 40,000 years ago. Homo sapiens replaced Neanderthals across Europe between approximately 45,000 and 35,000 years ago6, eventually giving rise to the ancestral population of Europeans alive today1.
This evidence from Apidima, along with other discoveries, demonstrates
that, on more than one occasion, modern humans kept pushing north and
westwards from Africa and the Levant into Europe. Rather than a single
exit of hominins from Africa to populate Eurasia, there must have been
several dispersals, some of which did not result in permanent
occupations by these hominins and their descendants.
There is
immense interest in understanding the timing and location of both the
successful and failed dispersals of hominins (including modern humans)
from Africa. The first hominin dispersal out of Africa is thought to
have been when members of the species Homo erectus exited some 2
million years ago. The second wave of departures occurred when the
ancestral species that eventually gave rise to Neanderthals moved into
Europe around 800,000–600,000 years ago.
A third group of migrations out of Africa were those of H. sapiens.
Many key fossil discoveries from Israel document early examples of
these dispersals. A fossil that includes the forehead region of a skull
found there, at a site called Zuttiyeh, is dated to between 500,000 and
200,000 years ago, and analysis of the fossil’s shape indicates that it
is either an early Neanderthal or from a population ancestral to both
Neanderthals and H. sapiens9. The Zuttiyeh fossil shows similarities to the Florisbad and Jebel Irhoud fossils9, and an earlier study10 suggested that Zuttiyeh might be an early H. sapiens.
This is a view that I favour, given its similarity to the shape of the
forehead of the Florisbad fossil. Future analysis might reveal that
Zuttiyeh is an even older modern human than Apidima 1; nevertheless, it
is not from Europe.
A jaw of an early modern human from Misliya Cave in Israel has been dated to approximately 194,000–177,000 years ago11. Other early modern human fossils have been found at Skhul and Qafzeh in Israel, dated to around 130,000–90,000 years ago12.
All of these early Eurasian human fossils seem to represent what might
be called ‘failed’ dispersals from Africa — they reached the Middle East
and southeastern Europe, but did not persist in these regions. There is
evidence that these populations were replaced at these or neighbouring
sites by Neanderthals.
Farther east, fossils of early H. sapiens
in Asia, dated from between at least 90,000 and 50,000 years ago, have
been found in regions ranging from Saudi Arabia to Australia13. These Asian fossils, like the European specimens of H. sapiens
from between 50,000 and 40,000 years ago, might have come from
populations that achieved persistent, successful dispersals and
contributed to the ancestry of some living humans.
Given that the
Apidima 1 fossil and those from Misliya and Zuttiyeh are only partial
skulls, some might argue that the specimens are too incomplete for their
status as H. sapiens to be certain. Could molecular approaches
be used to determine the species they are from? It is not always
possible to recover DNA from ancient fossils. However, analysing ancient
proteins preserved in fossils, a method termed palaeoproteomics, is
starting to be used to identify species (see go.nature.com/2xkosom).
Compared with analysis of ancient DNA, palaeoproteomics requires less
specialized handling of the fossil to prevent contamination. It was
recently used14
to analyse a fossilized jaw found in China that is approximately
160,000 years old, enabling the specimen to be identified as an
enigmatic hominin called a Denisovan, whose scarce fossils have also
been found at Denisova Cave in Siberia.
Perhaps palaeoproteomics
can be used to verify the identity of the Apidima fossils. It might also
be possible to apply this method to contemporaneous fossils from Asia
(estimated to be 300,000–150,000 years old) that have not yet been
definitively assigned to a species. These fossils are of interest for
their potential to reveal how many hominin species might have lived
during this time. Perhaps some of them are also H. sapiens, although I doubt it. Among the most complete of these specimens are crania from India at a site called Hathnora15, and from China at Dali16, Jinniushan16 and Hualongdong17.
Until such fossils are studied using palaeoproteomics, analyses such as
those of Harvati and colleagues provide our best handle on the complex
history of our species and our close relatives as these populations
dispersed out of Africa — from the early, unsuccessful dispersals to the
migrations that eventually succeeded.
Nature571, 487-488 (2019)
doi: 10.1038/d41586-019-02075-9
References
1.
O’Shea, N. & Delson, E. Nat. Hist.126(8), 19–22 (2018).
Uma rápida dispersão de humanos modernos da África para a Grécia
A análise de dois fósseis de uma caverna grega esclareceu os primeiros hominídeos na Eurásia.Um fóssil é o mais antigo espécime conhecido do Homo sapiens encontrado fora da África;o outro é um neandertal que viveu 40.000 anos depois.
A origem e a rápida dispersão do Homo sapiens tem sido objeto de interesse popular e acadêmico1. É quase universalmente aceito que o H. sapiens (humanos modernos) evoluiu na África, com os primeiros representantes fósseis conhecidos de nossa espécie datados de cerca de 315.000 anos atrás no Marrocos (em um local chamado Jebel Irhoud) 2 e aproximadamente 260.000 anos atrás no Sul África (em Florisbad) 3. Ferramentas de pedra comparáveis às encontradas nesses dois fósseis foram escavadas no Quênia (em Olorgesailie) 4 e datadas de cerca de 320.000 anos atrás. Escrevendo na revista Nature, Harvati et al.5 descrevem sua análise de um fóssil da Caverna Apidima, no sul da Grécia, que relatam ser um H. sapiens moderno primitivo de pelo menos 210.000 anos de idade. Este fóssil é o mais antigo humano moderno conhecido na Europa, e provavelmente em toda a Eurásia, e é mais de 160.000 anos mais velho do que o mais antigo fóssil europeu conhecido de H. sapiens6.
Leia o artigo: Os fósseis da caverna de Apidima fornecem evidências mais antigas do Homo sapiens na Eurásia
O complexo Apidima Cave foi escavado no final dos anos 70. Dois cranios parciais (crânios sem o maxilar inferior), denominados Apidima 1 e Apidima 2, foram recuperados em um único bloco de um tipo de rocha chamada brecha. Nenhum fóssil foi previamente descrito em detalhes. Apidima 2 inclui a região facial do crânio e foi identificada como Neanderthal7. A apidima 1 consiste apenas na parte de trás do crânio e não foi anteriormente alocada definitivamente a uma espécie. Harvati e seus colegas usaram tomografia computadorizada para escanear os fósseis e geraram uma reconstrução virtual em 3D de cada espécime. Eles analisaram cada fóssil para avaliar aspectos de sua forma e, assim, determinar a similaridade dos fósseis com os de outras espécies.
A Apidima 2 está bastante danificada devido a quebras e distorções anteriores. Análises das quatro reconstruções geradas do fóssil foram consistentes com o fato de ser um neandertal primitivo. Apidima 1 também está danificada, mas a amostra não está muito distorcida, então espelhar seus lados direito e esquerdo produziu uma boa reconstrução. A extensa análise comparativa dos autores indica que este fóssil é um dos primeiros membros do H. sapiens. A parte posterior do crânio é arredondada como a do H. sapiens, e carece de características clássicas do Neandertal, como o distintivo "chignon" occipital - uma protuberância na parte de trás do crânio que tem a forma de cabelo amarrado em um coque.
A datação anterior de um fragmento de Apidima 2 usando um método chamado análise de série de urânio indicava uma idade mínima de cerca de 160.000 anos. Harvati e seus colegas relatam um conjunto mais extenso de análises de datação por séries de urânio, que surpreendentemente revelam que Apidima 1 e Apidima 2 são de diferentes idades, embora tenham sido encontradas nas proximidades. Apidima 2 tem cerca de 170.000 anos - bem dentro da faixa etária de outros fósseis de neandertais encontrados em toda a Europa (Fig. 1). A Apidima 1 é datada de pelo menos 210.000 anos, o que é muito mais antigo do que qualquer outro fóssil amplamente aceito do H. sapiens encontrado fora da África.
Figura 1 | Alguns dos primeiros fósseis do Homo sapiens e espécies relacionadas na África e na Eurásia. Harvati et al.5 apresentam suas análises de dois crânios fósseis da caverna de Apidima, na Grécia. Eles relatam que o fóssil Apidima 1 é um espécime de H. sapiens que tem pelo menos 210.000 anos, de uma época em que os Neandertais ocupavam muitos locais europeus. É o mais antigo exemplo conhecido do H. sapiens na Europa e é pelo menos 160.000 anos mais velho que os fósseis mais antigos do H. sapiens encontrados na Europa6 (não mostrado). Harvati e seus colegas confirmam que, como relatado anteriormente7, Apidima 2 é um espécime de Neanderthal, e estimam que ele tenha pelo menos 170.000 anos de idade. As descobertas dos autores, juntamente com outras descobertas das quais uma seleção é mostrada aqui, esclarecem o momento e a localização das primeiras dispersões fracassadas da África de homininos (humanos modernos e outros parentes humanos, como Neandertais e Denisovanos). kyr, mil anos de idade.
Esta descoberta revela que pelo menos duas espécies de hominina (humanos e parentes humanos do ramo da árvore genealógica após nossa separação dos chimpanzés) habitaram o sudeste da Europa aproximadamente 200.000 anos atrás. A descoberta de um fóssil do H. sapiens em Apidima levanta questões sobre o que aconteceu com essa população. Dado que este H. sapiens existia numa época em que há evidências substanciais de uma presença de Neanderthal em outros locais europeus, fazia parte de uma população que não conseguia competir com sucesso com os neandertais, especialmente no clima instável da época? Talvez uma ou mais vezes as duas espécies se substituíssem como o principal grupo hominíneo presente nesta região.
Tais padrões de substituição caracterizam a distribuição dos humanos modernos e neandertais na região do Levante do Oriente Médio entre 250.000 e 40.000 anos atrás. O Homo sapiens substituiu os neandertais em toda a Europa entre aproximadamente 45.000 e 35.000 anos atrás6, dando origem,
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.

Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.