Previously unknown human species found in Asia raises questions about early hominin dispersals from Africa
Excavations in southeast Asia have unearthed a previously unreported hominin species named Homo luzonensis. The discovery has implications for ideas about early hominin evolution and dispersal from Africa.
Homo sapiens is the only living species of
a diverse group called hominins (members of the human family tree who
are more closely related to each other than they are to chimpanzees and
bonobos).
Most extinct hominin species are not our direct ancestors, but
instead are close relatives with evolutionary histories that took a
slightly different path from ours. Writing in Nature, Détroit et al.1
report the remarkable discovery of one such human relative that will no
doubt ignite plenty of scientific debate over the coming weeks, months
and years. This newly identified species was found in the Philippines
and named Homo luzonensis after Luzon, the island where bones and teeth from individuals of this species were excavated from Callao Cave. Specimens of H. luzonensis
were dated to minimum ages of 50,000 and 67,000 years old, which
suggests that the species was alive at the same time as several other
hominins belonging to the genus Homo, including Homo sapiens, Neanderthals, Denisovans and Homo floresiensis.
Rapidly changing knowledge about hominin evolution in Asia
is forcing the re-examination of ideas about early hominin dispersals
from Africa to Eurasia. Hominins appear in the fossil record about 6
million to 7 million years ago in Africa, and the earliest hominin
fossils in Eurasia are about 1.8 million years old2. Explanations for the earliest hominin dispersals from Africa fall under what is known as the Out of Africa I paradigm3. Modern humans only come into focus in the Out of Africa II paradigm, which refers to the early dispersals of H. sapiens from Africa to Eurasia that first occurred in the past 200,000 years4.
Ever since Homo erectus
was discovered in the early 1890s in Indonesia on the island of Java,
this species has essentially been the only character of interest in the
Out of Africa I dispersal events. The conventional viewpoint is that
this intrepid hominin began to stride gallantly towards far-off places
around 1.5 million to 2 million years ago (a dispersal that enabled it
to ultimately occupy territory across Africa and Eurasia)3.
Meanwhile, the other hominin species around at that time stayed in
Africa, living on borrowed time and facing imminent extinction. Compared
with H. erectus, these species — for example, other early Homo species such as Homo habilis, as well as the australopiths (hominins not in the genus Homo), which include Paranthropus and Australopithecus
— had smaller brains and an anatomy that is less similar to that of
modern humans. Indeed, as this simplified old story goes, how could such
species possibly compete given the anatomical and probable behavioural
advantages that H. erectus possessed?
Certain discoveries have challenged these entrenched ideas by raising the possibility that other hominins besides H. erectus dispersed from Africa to Eurasia during the Early Pleistocene (a period that occurred 2.58 million to 0.78 million years ago)5. Stone tools found in China are 2.1 million years old6, but no fossils as old as that have been attributed to H. erectus. It is also under debate whether H. erectus is the ancestor of H. floresiensis7, a species that lived on the Indonesian island of Flores. H. luzonensis provides yet more evidence that hints that H. erectus might not have been the only globe-trotting early hominin.
Détroit and colleagues have assigned seven teeth, two hand bones, three foot bones and one thigh bone to H. luzonensis. These remains, including one bone that was found previously8,
came from at least two adults and one child. The teeth include two
premolars and three molars from an individual’s upper jaw (Fig. 1).
Overall, these teeth and bones have a striking combination of
characteristics never before reported together in a hominin species.
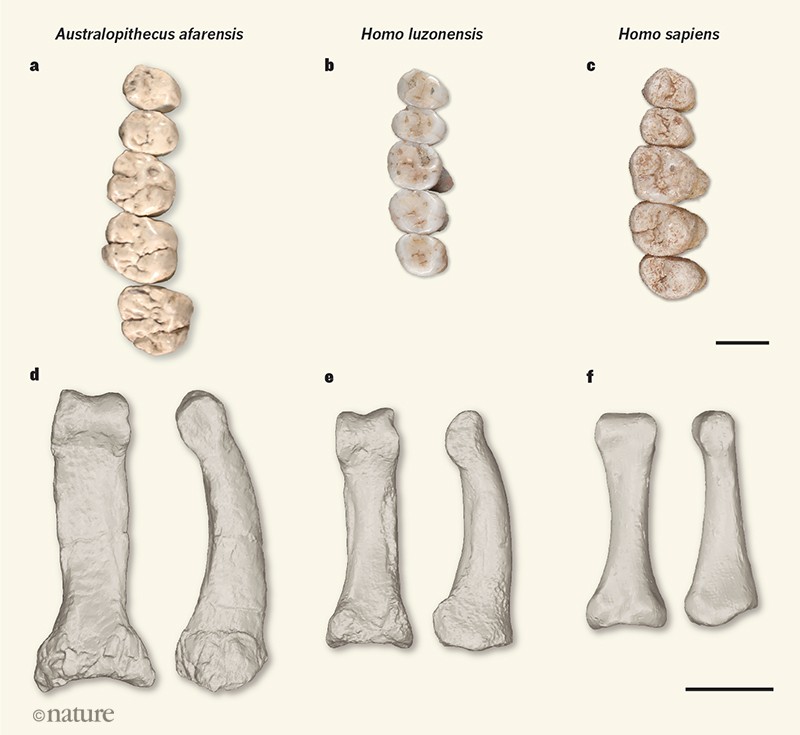
Figure 1 | Hominin teeth and toe bones. Détroit et al.1 report the discovery of a previously unknown extinct human species named Homo luzonensis, identified from teeth and bones excavated in the Philippines and dated to at least 50,000 years old. a–c, H. luzonensis premolar and molar teeth (b)
have a general pattern of a small tooth size and low tooth surface
complexity that is typical of premolars and molars from the genus Homo, including the teeth of Homo sapiens (c; a roughly 177,000-year-old specimen from Israel4).
This pattern contrasts with the larger and more complex surfaces of
premolars and molars of some early hominins that do not belong to the
genus Homo, such as Australopithecus afarensis (a; a cast of an approximately 3.4-million-year-old specimen from Ethiopia). The teeth in b are from the right side of the jaw; those in a and c are from the left side but are shown mirrored as from the right side for comparison with b. d–f, A 3D scan of a toe bone from H. luzonensis (e; bottom and side views shown) is strikingly similar to that of A. afarensis (d;
a 3D scan of an approximately 3.2-million-year-old specimen from
Ethiopia) in overall shape and curvature, and unlike the straighter and
more slender anatomy of H. sapiens toe bones (f; a 3D scan of a specimen from the Philippines from the 1800s). The interesting mix of features observed in H. luzonensis
raises key questions about the species’ ancestry and its relationships
with other human species. Scale bars represent 1 centimetre.
When compared with the molars of other hominin species, H. luzonensis
molars are astonishingly small, and the simplified surfaces of their
crowns and their low number of cusps are features that look similar to
the molar crowns and cusps of H. sapiens. Yet the shapes of H. luzonensis teeth share similarities with the teeth of H. erectus from Asia, and the size ratio of H. luzonensis premolars to molars is similar to that of Paranthropus,
species of which are known for their massive jaws and teeth. The
authors also used 3D imaging to examine the enamel–dentine junction
(EDJ), which is an internal region of the tooth where dentine material
meets the enamel layer. H. luzonensis premolar EDJs have a form that is distinct from that of hominins other than H. floresiensis. However, depending on the specific H. luzonensis molar that was analysed, the EDJs either look like those of H. erectus from Asia or like those of H. sapiens.
This strange juxtaposition of features in a single individual’s jaw is
completely unexpected and clouds our ability to reasonably assess, at
least for now, the exact evolutionary relationships between H. luzonensis and other hominin species.
Although attempts to extract DNA from H. luzonensis specimens have so far been unsuccessful, the anatomy of the foot and hand bones of H. luzonensis
strengthens the case that these remains represent a previously unknown
hominin species. A foot bone called the third metatarsal has an anatomy
in H. luzonensis that is distinct from that of other hominin species, including H. sapiens8. The authors’ 3D shape analyses of a H. luzonensis toe bone show that its shape is essentially indistinguishable from the toe bones of Australopithecusafarensis and Australopithecus africanus (Fig. 1), despite the separation of these australopiths from H. luzonensis by at least 2 million to 3 million years of evolution. Similar analyses found that a H. luzonensis finger bone most resembles the finger bones of australopiths and species of early Homo. Finally, the H. luzonensis
finger and toe bones are curved, which suggests that climbing was an
important part of this species’ behavioural repertoire, as was also the
case for many species of early hominin9.
The discovery of H. floresiensis raised the question of whether it evolved from a species of early Homo7,10,
for which evidence of its existence outside Africa has yet to be
documented. Scientists are still struggling to answer this definitively11,12. The question is more pressing with the discovery of H. luzonensis, given the need to explain yet another hominin species that, like H. floresiensis, was around during part of the Late Pleistocene (defined as a period of time 126,000 to 11,700 years ago), had Homo-like teeth but australopith-like hands and feet, and that lived on an island only reachable after a major sea crossing. Perhaps H. floresiensis and H. luzonensis are both descendants of H. erectus populations that evolved separately on their respective islands for hundreds of thousands of years, if not more13,14. However, explaining the many similarities that H. floresiensis and H. luzonensis share with early Homo
species and australopiths as independently acquired reversals to a more
ancestral-like hominin anatomy, owing to evolution in isolated island
settings, seems like a stretch of coincidence too far15.
Given the rich history of the Out of Africa I paradigm, unsurprisingly, H. erectus
has been the centre of attention in ideas about early hominin evolution
and dispersals in Asia. Nevertheless, it is worth considering how
different these ideas might be if, in the 1890s, H. floresiensis or H. luzonensis had been discovered rather than H. erectus. Because H. luzonensis provides the first glimpse of a second hominin species living on a distant island at a time when H. sapiens
populations from Africa were beginning to spread across the world, one
thing can be said for certain — our picture of hominin evolution in Asia
during the Pleistocene just got even messier, more complicated and a
whole lot more interesting.

Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.