Cetiosaurus Owen, 1841 (Dinosauria, Sauropoda): proposed conservation of usage by designation of Cetiosaurus oxoniensis Phillips, 1871 as the type species Paul Upchurch Department of Earth Sciences, University College London, Gower Street, London WC1E 6BT, U.K. (e-mail: p.upchurch@ucl.ac.uk)
John Martin 6 The Nook, Great Glen, Leicester LE8 9GQ, U.K.
(e-mail:
Johnmartin424@aol.com)
Michael P. Taylor
Palaeobiology Research Group, School of Earth & Environmental Sciences, Burnaby Building, Burnaby Road, University of Portsmouth, Portsmouth, PO1 3QL, U.K. (e-mail: dino@miketaylor.org.uk)
Abstract. The purpose of this application, under Article 81.1 of the Code, is to preserve stability in the taxonomy of sauropod dinosaurs by designating Cetiosaurus oxoniensis as the type species of Cetiosaurus. The genus Cetiosaurus (including the species C. medius and C. oxoniensis) was established during the earliest period of research on sauropod dinosaurs, and is historically significant. The name Cetiosaurus was fixed to the type species
Cetiosaurus medius, a sauropod of indeterminate affinities; however, the fragmentary nature of the type material of C. medius,
combined with the subsequent description of much more complete Middle Jurassic
sauropod material as Cetiosaurus oxoniensis, has meant that subsequent literature has
overwhelmingly adopted C. oxoniensis over C. medius as the primary exemplar of
Cetiosaurus. Stability would be best served by designating Cetiosaurus oxoniensis
as the type species of the genus Cetiosaurus in place of the current type species,
C. medius.
Keywords. Nomenclature; taxonomy; Dinosauria; Sauropoda; CETIOSAURIDAE;
Cetiosaurus; Cetiosaurus oxoniensis; Cetiosaurus medius; England; Europe; Middle
Jurassic.
1. The generic name Cetiosaurus was first published by Owen (1841, p. 457) without any associated specific name. It was based primarily on material found by John Kingdon in 1825, but no specimen numbers were given. Cetiosaurus was among the first named sauropod dinosaurs and, as a result, has become a ‘wastebasket’ taxon, with much material indiscriminately referred to it. As detailed by Upchurch &
Martin (2003, p. 208), the stratigraphic range of Cetiosaurus, if all referrals were
supported, would extend from the Bajocian (Middle Jurassic) to the Barremian
(Early Cretaceous), a range of about 45 million years.
2. The first published species of Cetiosaurus were C. hypoolithicus Owen, 1842 and
C. epioolithicus Owen, 1842, both published in the same report (Owen, 1842a, p. 12).
The type material was not specified for either species, and neither was illustrated or
diagnosed; therefore both species are nomina nuda, and are ineligible for fixation as
the type species (Upchurch & Martin, 2003, p. 209).
3. The next published species of Cetiosaurus were C. brevis, C. brachyurus, C.
medius and C. longus, all described in a single report by Owen (1842b, pp. 94, 100a,
100b, 101). Since descriptions of all four species were furnished, they are not nomina
nuda and are thus available names for fixation as the type species. Although Owen
did not explicitly designate any of these species as the type, he did note of the C.
medius material that ‘it is principally on these bones, with others subsequently
discovered and in the collection of Mr. Kingdon, that the characters of the
Cetiosaurus were first determined’ (Owen, 1842b, pp. 100–101). C. medius is thus the
type species of Cetiosaurus under Article 69.1.1 of the Code (Type species by
subsequent designation), an interpretation endorsed by, for example, Steel (1970,
p. 64).
4. Cetiosaurus oxoniensis was described by Phillips (1871, p. 291) from a large series
of remains from the Forest Marble (Bathonian, Middle Jurassic of Oxfordshire),
among which no type specimen was formally designated. Phillips (1871, pp. 290–291)
wrote only ‘I propose for the species found in the immediate vicinity of Oxford and
elsewhere, the only one for which sufficient materials are collected to serve for
determining its characters, the title Oxoniensis’, so that Phillips’s specimens found in
this area form a syntype series. Among these specimens, the largest of three
individuals from Bletchingdon Station, near Enslow Bridge, is the most complete and
diagnostic; it was therefore designated as the lectotype (OUMMNH J13605–13613,
J13615–16, J13619-J13688, J13899 in the Oxford University Museum of Natural
History) by Upchurch & Martin (2003, p. 216). The material represents a nonneosauropod
eusauropod. It is important due to its historical significance as the first
sauropod known from adequate remains, illustrating the basic sauropod body plan,
and also because of the light it casts on the evolution of sauropods, being one of
the most derived taxa outside the clade Neosauropoda (Upchurch, 1998, fig. 19;
Upchurch et al., 2004, fig. 13.18). Cetiosaurus is the type genus of the family
CETIOSAURIDAE Lydekker, 1888 and was used as a specifier in the phylogenetic
definition of the clade CETIOSAURIDAE (Upchurch et al., 2004, p. 301).
5. Several further species of Cetiosaurus have also been erected. As summarised by
Upchurch & Martin (2003, p. 215), of the thirteen named species, three are nomina
nuda, two are junior objective synonyms, four are nomina dubia, and four are
diagnosable taxa (C. brevis, C. oxoniensis, C. glymptonensis Phillips, 1871, p. 291 and
C. humerocristatus Hulke, 1874, p. 17). These last four cannot be congeneric as they
represent several different sauropod groups.
6. Under a strict application of the Code, Cetiosaurus medius is the type species of
Cetiosaurus. However, the name Cetiosaurus has invariably been associated with the
species C. oxoniensis, and specifically the Bletchingdon Station material (e.g. Owen,
1875; Hatcher, 1903; Huene, 1904, 1927; Fraas, 1908; Janensch, 1914, 1929;
Matthew, 1915; Coombs, 1975; Wild, 1978; Bonaparte, 1986, 1999; Martin, 1987;
Upchurch, 1998; Casanovas et al., 2001; Upchurch & Martin, 2002, 2003; Liston,
2004; Day et al., 2004; Upchurch et al., 2004; Sánchez-Hernández, 2005; Wedel, 2005, 2007; Yates, 2006, 2007; Galton & Knoll, 2006; Moser et al., 2006; Naish & Martill,
2007; Taylor & Naish, 2007). The stability of use of this species in the literature as
representing Cetiosaurus is indicated by the fact that no other generic name has ever
been proposed for C. oxoniensis; nor has it ever been referred to as ‘Cetiosaurus’
oxoniensis. Enforcing the strict application of the Code would lead to considerable
nomenclatural confusion and inconsistency.
6. For this reason, Upchurch & Martin (2003, p. 215) informally treated
Cetiosaurus oxoniensis as the type species of Cetiosaurus pending a promised petition
to the Commission.
7. The International Commission on Zoological Nomenclature is accordingly
asked:
(1) to use its plenary power to set aside all previous fixations of type species for the
nominal genus Cetiosaurus Owen, 1841 and to designate Cetiosaurus oxoniensis
Phillips, 1871 as the type species;
(2) to place on the Official List of Generic Names in Zoology the name Cetiosaurus
Owen, 1841 (gender: masculine), type species Cetiosaurus oxoniensis Phillips,
1871, as ruled in (1) above;
(3) to place on the Official List of Specific Names in Zoology the name oxoniensis
Phillips, 1871, as published in the binomen Cetiosaurus oxoniensis (specific
name of the type species of Cetiosaurus Owen, 1841), as ruled in (1) above.
References
Bonaparte, J.F. 1986. Les dinosaures (Carnosaures, Allosauridés, Sauropodes, Cétiosauridés)
du Jurassique moyen de Cerro Cóndor (Chubut, Argentina). Annales de Paléontologie, 72:
325–386.
Bonaparte, J.F. 1999. Evolucion de las vertebras presacras en Sauropodomorpha. Ameghiniana,
36: 115–187.
Casanovas, M.L., Santafé, J.V. & Sanz, J.L. 2001. Losillasaurus giganteus, un neuvo
saurópodo del tránsito Jurásico-Cretácico de la cuenca de ‘Los Serranos’ (Valencia,
España). Paleontologia i Evolució, 32–33: 99–122.
Coombs, W.P. 1975. Sauropod habits and habitats. Palaeogeography, Palaeoclimatology,
Palaeoecology, 17: 1–33.
Day, J.J., Norman, D.B., Gale, A.S., Upchurch, P. & Powell, H.P. 2004. A Middle Jurassic
dinosaur trackway site from Oxfordshire, UK. Palaeontology, 47: 319–348.
Fraas, E. 1908. Ostafrikanische Dinosaurier. Palaeontographica, 55: 105–144.
Galton, P.M. & Knoll, F. 2006. A saurischian dinosaur braincase from the Middle Jurassic
(Bathonian) near Oxford, England: from the theropod Megalosaurus or the sauropod
Cetiosaurus? Geological Magazine, 143: 905–921.
Hatcher, J.B. 1903. Osteology of Haplocanthosaurus with description of a new species, and
remarks on the probable habits of the Sauropoda and the age and origin of the
Atlantosaurus beds. Memoirs of the Carnegie Museum, 2: 1–72, pls. I–V.
Huene, F.v. 1904. Dystrophaeus viaemalae Cope in neuer Beleuchtung. Neues Jahrbuch für
Mineralogie, Geologie, und Paläontologie Abhandlungen, 19: 319–333.
Huene, F.v. 1927. Sichtung der Grundlagen der jetzigen Kenntnis der Sauropoden. Eclogae
Geologicae Helvetiae, 20: 444–470.
Hulke, J.W. 1874. Note on a very large saurian limb-bone adapted for progression upon land,
from the Kimmeridge Clay of Weymouth, Dorset. Quarterly Journal of the Geological
Society, 30: 16–17 and Plate II.
Janensch, W. 1914. Übersicht über der Wirbeltierfauna der Tendaguru-Schichten nebst einer
kurzen Charakterisierung der neu aufgefuhrten Arten von Sauropoden. Archiv fur
Biontologie, 3: 81–110.
Janensch, W. 1929. Material und Formengehalt der Sauropoden in der Ausbeute der
Tendaguru-Expedition. Palaeontographica (Suppl. 7), 2: 1–34.
Liston, J.J. 2004. A re-examination of a Middle Jurassic sauropod limb bone from the
Bathonian of the Isle of Skye. Scottish Journal of Geology, 40: 119–122.
Lydekker, R. 1888. Catalogue of the fossil Reptilia and Amphibia in the British Museum
(Natural History), Cromwell Road, S.W. Part 1. Containing the orders Ornithosauria,
Crocodylia, Dinosauria, Squamata, Rhynchocephalia, and Proterosauria. British Museum
(Natural History), London.
Martin, J. 1987. Mobility and feeding of Cetiosaurus (Saurischia: Sauropoda) – why the long
neck? Occasional Papers of the Tyrrell Museum of Palaeontology (Fourth Symposium on
Mezozoic Terrestrial Ecosystems), 3: 154–159.
Matthew, W.D. 1915. Dinosaurs, with special reference to the American Museum collections.
American Museum of Natural History, New York.
Moser, M., Mathur, U.B., Fürsich, F.T., Pandey, D.K. & Mathur, N. 2006. Oldest camarasauromorph
sauropod (Dinosauria) discovered in the Middle Jurassic (Bajocian) of the
Khadir Island, Kachchh, western India. Paläontologische Zeitschrift, 80: 34–51.
Naish, D. & Martill, D.M. 2007. Dinosaurs of Great Britain and the role of the Geological
Society of London in their discovery: basal Dinosauria and Saurischia. Journal of the
Geological Society, London, 164: 493–510.
Owen, R. 1841. A description of a portion of the skeleton of the Cetiosaurus, a gigantic extinct
saurian reptile occurring in the oolitic formations of different portions of England.
Proceedings of the Geological Society of London, 3: 457–462.
Owen, R. 1842a. Second rapport sur les reptiles fossiles de la Grande-Bretagne. L’Institut,
Journal général des Sociétés et Travaux Scientifique de la France et de l’ Étranger, 10:
11–14.
Owen, R. 1842b. Report on British fossil reptiles, Part II. Reports of the British Association for
the Advancement of Science, 11: 60–204.
Owen, R. 1875. Monograph of the Mesozoic Reptilia, part 2: Monograph on the genus
Cetiosaurus. Palaeontolographical Society Monograph, 29: 27–43.
Phillips, J. 1871. Geology of Oxford and the valley of the Thames. Clarendon Press, Oxford.
Steel, R. 1970. Handbuch der Paläoherpetologie. Part 14. Saurischia. Gustav Fischer Verlag,
Stuttgart.
Sánchez-Hernández, B. 2005. Galveosaurus herreroi, a new sauropod dinosaur from Villar del
Arzobispo Formation (Tithonian-Berriasian) of Spain. Zootaxa, 1034: 1–20.
Taylor, M.P. & Naish, D. 2007. An unusual new neosauropod dinosaur from the Lower
Cretaceous Hastings Beds Group of East Sussex, England. Palaeontology, 50: 1547–1564.
Upchurch, P. 1998. The phylogenetic relationships of sauropod dinosaurs. Zoological Journal
of the Linnean Society, 124: 43–103.
Upchurch, P. & Martin, J. 2002. The Rutland Cetiosaurus: The anatomy and relationships of
a Middle Jurassic British sauropod dinosaur. Palaeontology, 45: 1049–1074.
Upchurch, P. & Martin, J. 2003. The anatomy and taxonomy of Cetiosaurus (Saurischia,
Sauropoda) from the Middle Jurassic of England. Journal of Vertebrate Paleontology, 23:
208–231.
Upchurch, P., Barrett, P.M. & Dodson, P. 2004. Sauropoda. Pp. 259–322 in Weishampel, D.B.,
Dodson, P. & Osmólska, H. (Eds.), The Dinosauria, 2nd edition. University of California
Press, Berkeley and Los Angeles.
Wedel, M.J. 2005. Postcranial skeletal pneumaticity in sauropods and its implications for mass
estimates. Pp. 201–228 in Wilson, J.A. & Curry Rogers, K. (Eds.), The sauropods:
evolution and paleobiology. University of California Press, Berkeley.
Wedel, M.J. 2007. What pneumaticity tells us about ‘prosauropods’, and vice versa. Pp.
207–222 in Barrett, P.M. & Batten, D.J. (Eds.), Special Papers in Palaeontology 77:
evolution and palaeobiology of early sauropodomorph dinosaurs. The Palaeontological
Association, U.K.
Wild, R. 1978. Ein Sauropoden-Rest (Reptilia, Saurischia) aus dein Posidonienschiefer (Lias,
Toarcium) von Holzmaden. Stuttgarter Beiträge zur Naturkunde, Serie B (Geologie und
Paläontologie), 41: 4–15.
Yates, A.M. 2006. Solving a dinosaurian puzzle: the identity of Aliwalia rex Galton. Historical
Biology, 19: 93–123.
Yates, A.M. 2007. The first complete skull of the Triassic dinosaur Melanorosaurus Haughton
(Sauropodomorpha: Anchisauria). Pp. 9–55 in Barrett, P.M. & Batten, D.J. (Eds.),
Special Papers in Palaeontology 77: Evolution and palaeobiology of early sauropodomorph
dinosaurs. The Palaeontological Association, U.K.
Acknowledgement of receipt of this application was published in BZN 65: 162.
Comments on this case are invited for publication (subject to editing) in the Bulletin; they
should be sent to the Executive Secretary, I.C.Z.N., c/o Natural History Museum, Cromwell
Road, London SW7 5BD, U.K. (e-mail: iczn@nhm.ac.uk).





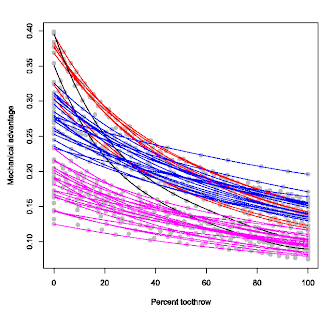 It may occur to some that maybe this profile would be consistent across theropod taxa; after all, they all seem to have longish tooth rows with little variation in tooth row morphology. This is true to some extent that the majority of the profile morphology is relatively similar, in that they are all simple parabolic curves. However, there are major differences in the vertical positioning of the curves, or the absolute values of the mechanical advantages (see figure to the left). This graph is from an older version so it's not exactly the same one in the final paper, but the overall pattern is the same; you see a vertical separation in biting profiles, albeit along a continuum (kind of like a smear I guess). Nonetheless, there are prominently unique profiles. For instance the ones in black, Coelophysis and Syntarsus. These profiles criss-cross the entire vertical spectrum from very high mechanical advantages at the back of the tooth row to very low mechanical advantages at the front of the tooth row. Other theropods do not have this extreme combination.
It may occur to some that maybe this profile would be consistent across theropod taxa; after all, they all seem to have longish tooth rows with little variation in tooth row morphology. This is true to some extent that the majority of the profile morphology is relatively similar, in that they are all simple parabolic curves. However, there are major differences in the vertical positioning of the curves, or the absolute values of the mechanical advantages (see figure to the left). This graph is from an older version so it's not exactly the same one in the final paper, but the overall pattern is the same; you see a vertical separation in biting profiles, albeit along a continuum (kind of like a smear I guess). Nonetheless, there are prominently unique profiles. For instance the ones in black, Coelophysis and Syntarsus. These profiles criss-cross the entire vertical spectrum from very high mechanical advantages at the back of the tooth row to very low mechanical advantages at the front of the tooth row. Other theropods do not have this extreme combination.













{kind=link}