The
capture and enslavement of eukaryotic algae by unicellular predators to
acquire photosynthesis was a major driving force in early eukaryotic
diversification. A genome presented in BMC Biology
provides a glimpse of how such a tiny predator might have preyed on red
algae and detained them to create new lineages of photosynthetic
organisms.
Few biologists will have heard of Goniomonas,
though they could find this minute, colorless biflagellate in just
about any stream or inlet they cared to look in. But why should they
care? Well, Goniomonas is
an important piece in the puzzle of how the colorful flags of
photosynthesis came to be daubed across numerous different and unrelated
branches of the tree of eukaryotes [1].
Goniomonas
first came to my attention 25 years ago when friend and cryptomonad
aficionado David Hill showed me a little ‘bug’ he had retrieved from the
Yarra River in Melbourne. Once I had grasped what I was viewing, I knew
we had to get its DNA and try to unravel how Goniomonas
is related to the photosynthetic cryptomonads. Cryptomonads, you see,
are the ‘smoking gun’ of a remarkable and massively influential
evolutionary process known as secondary endosymbiosis.
Plastids—the
subcellular structures in plants and algae where photosynthesis
occurs—first arose by endosymbiosis of a cyanobacterium. The
cyanobacterium was engulfed by a eukaryotic phagotroph, but instead of
being digested for food as normally would happen to a phagotroph’s prey,
it was detained as a solar cell to perform the metabolic magic of using
light energy to smash water, release electrons, and string carbon atoms
together to create the fuel of life—glucose. This initial
endosymbiosis, referred to as the primary endosymbiosis,
spawned the red algae, the glaucophytes, and the green algae and their
eventual descendants, the plants. The plastids in these
photosynthesizers have two bounding membranes, now clearly understood to
derive from the pair of membranes that enclose your typical,
Gram-negative cyanobacterium.
But
primary endosymbiosis was merely the first chapter in the acquisition
of photosynthesis by eukaryotes. Once the cyanobacterium was tamed, it
was transplanted horizontally into several disjunct eukaryotic lineages
by multiple secondary endosymbioses. These secondary
endosymbioses—sometimes involving a red alga and sometimes involving a
green alga, but curiously never a glaucophyte—resulted in the eukaryotic
alga becoming the detainee in one or another phagotroph to create a
metaorganism comprising a relic cyanobacterium inside a eukaryote inside
yet another eukaryote. How do we know this with certainty?
Cryptomonads. These little algae are a classic Darwinian transition form
in this shell game, retaining all the shells of the two sequential
engulfment processes, plus a few tell-tale bits and pieces of the
swallowed ones.
The
crucial piece in cryptomonads is the vestigial nucleus of the engulfed
red alga, which we now refer to as the nucleomorph in recognition of
Dennis Greenwood’s seminal observation that it had the morphology of a
little nucleus sequestered between the shells (membranes) of the
metaorganism (references in [2]).
Nucleomorph genomes confirm that this little structure is indeed the
relic nucleus of the red algal endosymbiont and prove unequivocally that
secondary endosymbiosis did happen, and that cryptomonads are one
result [3].
Nucleomorphs give credence to all the other putative secondary
endosymbioses where the nucleomorph, and even some of the membrane
shells, have apparently disappeared with reduction of the endosymbiont
through increased integration into the metaorganisms.
But back to Goniomonas. The German phycologist Georg Fresenius recognized Goniomonas
as a cryptomonad 160 years ago on the basis of its asymmetric shape,
presence of ejectisomes (enigmatic ejectile structures that burst like
party streamers from the cell under duress), and the two unequal
flagella emerging from a deep, complicated anterior invagination [4] (Fig. 1). Where Goniomonas
differs to most cryptomonads—which are either blueish-greenish or
reddish-brownish depending on which of the endosymbiont’s phycobilin
pigments they retain—is it lacks any color, and apparently any plastid [5].
This raises the question of whether it had a plastid and carelessly
lost it, or whether it is a representative of the cryptomonads before
they entered into their complex secondary endosymbiotic relationship
with a red alga? A first step would be to infer how Goniomonas relates to photosynthetic cryptomonads, and DNA sequences were the obvious approach.
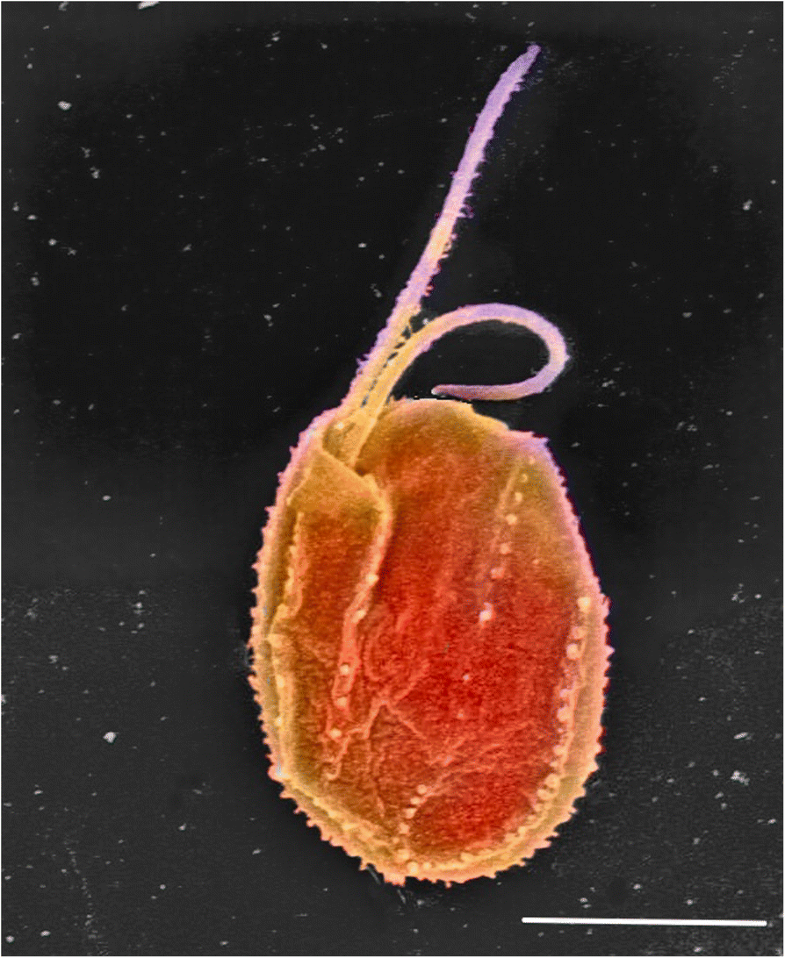
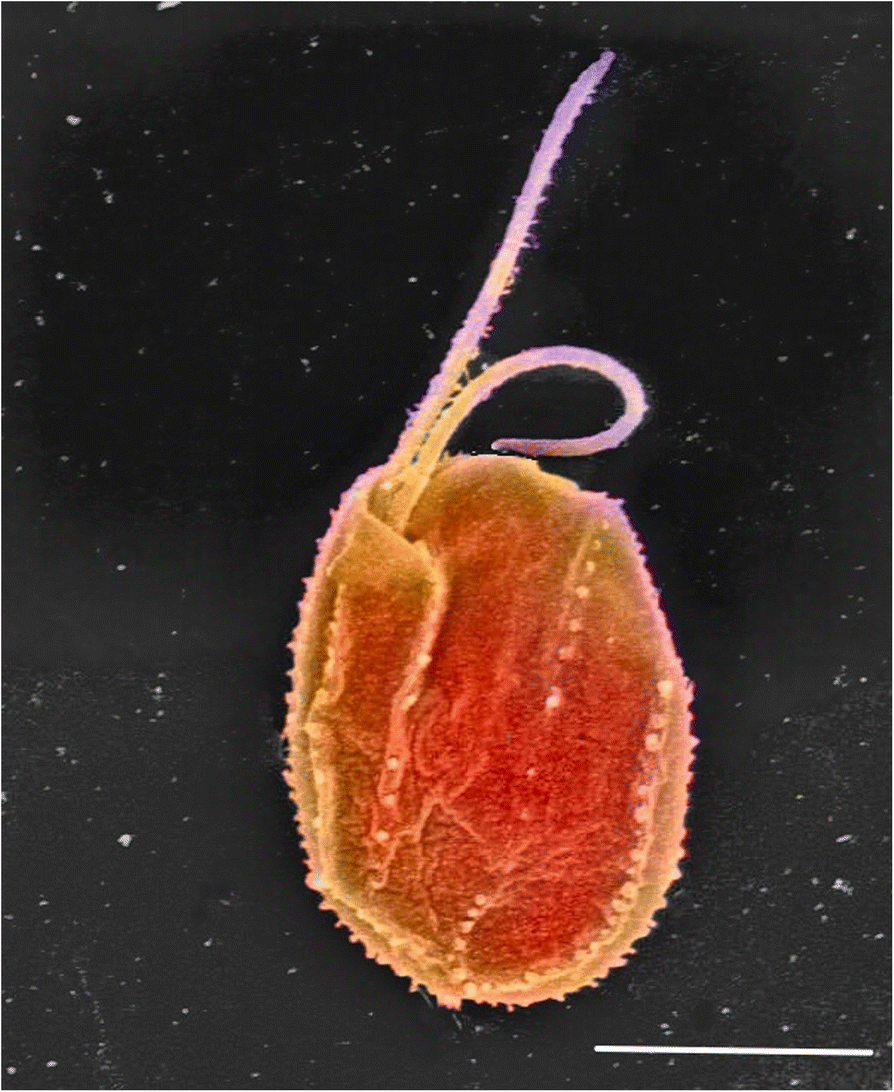
Fig. 1.
Scanning electron micrograph of the tiny predator Goniomonas avonlea
(colourised). Typical cryptomonad features such as the flat, asymmetric
shape, two anterior flagella emerging from a complex invagination, and
the light armour plating are visible. Photo credit E. Kim. Bar = 5 μm
To get DNA sequences from organisms one typically needs a goodly amount of them. Goniomonas
is a predator, so when Hill and I wanted to grow it up in large
numbers, we opted to provide it with loads of bacteria to engulf and
digest—this was as simple as me popping a grain of wheat into several
flasks of culture media to support bacterial growth, whilst Hill
arduously plucked out individual Goniomonas
cells with a micropipette and placed them in solitary confinement with
only food for company. Hill and I got a clonal culture and a ribosomal
RNA sequence from Goniomonas truncata, and could show it was sister to photosynthetic cryptomonads [6].
In their recent article in BMC Biology, Cenci et al. report the entire genome of a related species, Goniomonas avonlea [7], which hails from the setting of Lucy Montgomery’s novel Anne of Green Gables. The G. avonlea
genome paints the first complete picture of how cryptomonads might have
functioned before they crossed the tracks and made a pact with an alga
to become autotrophic. It is a fascinating window into how things
probably were with cryptomonads before they switched lifestyles.
As genomes go, the G. avonlea
blueprint is still a bit of a roughie, remaining in ~ 32,000 unjoined
pieces. That’s because the genome is huge for such a tiny organism, with
a final size approaching 100 megabases; the effort to get it fully
assembled and polished would be formidable. Gene count, at ~ 18,000
non-redundant proteins, is also impressive, and these genes are, on
average, interrupted by about five introns each. But numbers aside, what
does the genome tell us about how G. avonlea makes its living, now and in the past? Quite a bit as it happens. A concerted search turned up no convincing evidence for G. avonlea
now having, or ever having had, a plastid. Thus, we can be pretty
certain this type of cryptomonad is ancestrally heterotrophic. Metabolic
pathways typically taken care of by the plastid in plants and
algae—namely fatty acid synthesis, isoprenoid precursor synthesis, iron
sulfur cluster generation, and heme synthesis—are apparently done in the
cytosol of G. avonlea
using canonical eukaryotic machinery not related to cyanobacteria. Thus,
there are no traces of plastid-type metabolisms lurking in the genome,
and Goniomonas appears to be a living representative of the pre-secondary-endosymbiosis cryptomonads.
G. avonlea
also seems well equipped gene-wise to digest its prey, having a panoply
of lysozymes to chew through the cell walls of those bacteria
unfortunate enough to end up going down the gullet of the predator and
into its food vacuoles. For me though, the jewel-in-the-crown of the G. avonlea
genome are glycan hydrolases belonging to the GH50 family, which cleave
β-1,4 glycosidic bonds of agarose, a principal component of red algal
cell walls. G. avonlea thus
seems equipped to be algivorous, capable of digesting the walls of red
algal prey cells. In today’s oceans and streams, most red algae are
multicellular and too large to be preyed upon by a miniscule flagellate
like G. avonlea, but unicellular red algae small enough for a Goniomonas cell to engulf are not uncommon and were perhaps more so in earlier times. Thus, if G. avonlea
does indeed engulf and digest red algae, it is not too much of a
stretch to imagine a scenario where the prey cell is detained and not
digested—exactly the kind of event predicted to have occurred at the
outset of a secondary endosymbiosis creating the photosynthetic
cryptomonads [2, 3].
Goniomonas
is thus a nice fit for the ancestral phagotroph that was routinely
capturing and digesting red algae and could have commenced a longer and
more sustained relationship with its prey to embark on the acquisition
of photosynthesis through a secondary endosymbiosis. But what does Goniomonas tell us about the other, non-cryptomonad eukaryotes that also have reduced red algal endosymbionts for plastids?
Major
eukaryote groups including the heterokonts/stramenopiles (algae like
brown kelps, diatoms, and golden flagellates), haptophytes (abundant
limestone-armoured phytoplankton whose dead ancestors comprise most of
the white cliffs of Dover), dinoflagellates (including the symbionts of
corals crucial for reef building and the toxic basis of certain red
tides), and alveolates (parasites of animals and protists that cause
diseases such as malaria and toxoplasmosis and possess relic,
non-photosynthetic plastids) all harbour secondary endosymbionts of red
algal origin. A long-standing debate about whether all these different
types of organisms gained their red algal endosymbiont in one
event—perhaps akin to the one discussed here in which a Goniomonas-like
phagotroph captured and retained a red alga—or whether each of them
descends from a separate capture of a red alga by different ancestors
remains wide open, despite 40 years of investigation [8, 9, 10].
The genome of Goniomonas
doesn’t yet resolve this debate, but it gives us an extant model with
which to explore what the phagotrophic partner in the extraordinary
amalgam that led to at least one group of complex algae, the
cryptomonads, was like. We might just have extant descendants of the two
partners—predator and prey—with which to better understand how a great
swathe of eukaryotic diversity originated through secondary
endosymbiosis. Maybe we should feed our Goniomonas on red algae rather than bacteria.
GIM is supported by an Australian Research Council Laureate Fellowship.
Availability of data and materials
Not applicable.
Author’s contributions
GIM wrote the manuscript. GIM read and approved the final manuscript.
Competing interests
The author declares that he has no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Open AccessThis article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/),
which permits unrestricted use, distribution, and reproduction in any
medium, provided you give appropriate credit to the original author(s)
and the source, provide a link to the Creative Commons license, and
indicate if changes were made. The Creative Commons Public Domain
Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
Douglas
S, Zauner S, Fraunholz M, Beaton M, Penny S, Deng L-T, Wu X, Reith M,
Cavalier-Smith T, Maier U-G. The highly reduced genome of an enslaved
algal nucleus. Nature. 2001;410:1091–6.View ArticleGoogle Scholar
Fresenius G. Beiträge zur Kenntniss mikroskopischer Organismen. Abh Senckenberg Naturforsch Ges. 1858;2:1–34.Google Scholar
Kim
E, Archibald JM. Ultrastructure and molecular phylogeny of the
cryptomonad Goniomonas avonlea sp. nov. Protist. 2013;164:160–82.View ArticleGoogle Scholar
McFadden
GI, Gilson PR, Hill DRA. Goniomonas—rRNA sequences indicate that this
phagotrophic flagellate is a close relative of the host component of
cryptomonads. Eur J Phycol. 1994;29(1):29–32.View ArticleGoogle Scholar
Cenci
U, Sibbald S, Curtis B, Kamikawa R, Eme L, Moog D, Henrissat B,
Marechal E, Chabi M, Djemel C, et al. Nuclear genomne sequene of the
plastid lacking cryptomonad Goniomonas avonlea: phylogenomic and
metabolic insights into the evolution of secondary plastids. BMC Biol.
2018. https://doi.org/10.1186/s12915-018-0593-5.
Cavalier-Smith
T. The kingdom Chromista: origin and systematics. In: Round F, Chapman
D, editors. Progress in phycological research, vol. 3. Bristol:
Biopress; 1986. p. 309–47.Google Scholar
Gould SB, Maier UG, Martin WF. Protein import and the origin of red complex plastids. Curr Biol. 2015;25(12):R515–21.View ArticleGoogle Scholar
Waller
RF, Gornik SG, Koreny L, Pain A. Metabolic pathway redundancy within
the apicomplexan-dinoflagellate radiation argues against an ancient
chromalveolate plastid. Commun Integr Biol. 2016;9(1):e1116653.View ArticleGoogle Scholar
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.