Os tecidos moles de muitos vertebrados fósseis preservam evidências de melanossomos - organelas em escala micro que informam sobre coloração tegumentar e estratégias de comunicação. Nos vertebrados existentes, no entanto, os melanossomas também ocorrem nos tecidos internos. Assim, os melanossomas fósseis podem não derivar apenas do tegumento e seus apêndices. Aqui, analisando os sapos existentes e fósseis, mostramos que os melanossomas não tegumentares têm alto potencial de fossilização, superam largamente os da pele, e potencialmente dominam os filmes de melanossomos preservados em alguns vertebrados fósseis. Os nossos experimentos de decaimento mostram que os melanossomas não tegumentares geralmente permanecem in situ desde que as carcaças não sejam perturbadas. O estudo de fósseis em escala de mícron, no entanto, demonstra que os melanossomas não tegumentares podem redistribuir através de partes do corpo se as carcaças forem perturbadas pelas correntes. Coletivamente, esses dados indicam que os melanossomas fósseis nem sempre estão relacionados à coloração tegumentar. Melanossomas tegumentar e não tegumentar podem ser discriminados usando a geometria e distribuição do melanossomo. Isto é essencial para reconstruções precisas das cores tegumentares dos vertebrados fósseis.
Introduction
Melanin is a key component of visual signals in animals through its incorporation into integumentary patterning1.
In vertebrates, melanin occurs in skin and its derivatives as discrete
micron-sized membrane-bound organelles termed melanosomes2.
Physical and chemical evidence of melanin has been used to infer the
plumage colours of fossil birds and feathered dinosaurs (reviewed in
ref.3) and integumentary coloration in fossil marine reptiles4 (but see refs.5,6,7).
Reconstructions of original skin colour rely on evidence that fossil
microstructures are melanosomes (and not, for example, bacteria5)
and that fossil melanosomes derive solely from the integument.
Melanosomes occur, however, in various internal organs in extant
vertebrates8,9,10
and thus melanosomes preserved in the body outline of fossils will not
always indicate integumentary colour. Given the high recalcitrance of
melanin and melanosomes11,
non-integumentary melanosomes (defined here as those from internal
organs and tissues and excluding the eyes) may persist during
fossilization, but this remains to be demonstrated; the decay
microenvironment within carcasses and the external environment often
differ markedly in chemistry12,13.
If non-integumentary melanosomes are decay-resistant and abundant in
vivo, reconstruction of integumentary coloration based on the
presence/absence, shape and distribution of fossil melanosomes requires
that melanosomes from different tissue sources can be discriminated.
Recent studies of colour reconstruction14,15
have opted to avoid sampling the torso, in attempts to avoid sampling
non-integumentary melanosomes.
This does not, however, guarantee that
melanosomes from the limbs and tail are integumentary because
non-integumentary melanosomes may be redistributed throughout parts of
the body during decay. In key biotas such as Jehol, fossils are often
partly disarticulated by the action of bottom currents close to the lake
floor and can preserve evidence for rupturing of organs and
redistribution of their contents throughout the body16.
Intuitively, such disturbance of exposed carcasses by current activity
could also potentially distribute melanosomes from degraded organs
inside the body. Whether such redistribution is likely in the absence of
physical disturbance (e.g., when organs decompose or the carcass
ruptures from gas pressure) is unknown. Resolving these critical issues
requires a systematic and comprehensive test of (1) whether
non-integumentary melanosomes are abundant and can survive decay and (2)
under what taphonomic scenarios are they likely to redistribute.
Aqui, abordamos essas questões usando histologia, microscopia de fluorescência, microscopia eletrônica de varredura (SEM) e oxidação de hidróxido alcalino (AHPO) 11 de tecidos de sapos existentes recentemente mortos e experimentalmente decompostos, juntamente com dados de fósseis. As rãs são um caso de teste ideal, uma vez que melanossomos foram relatados em órgãos internos de espécies sobreviventes8,9,10 e são conhecidos exemplos de fósseis que preservam melanossomas13. Nossos dados mostram que os melanossomas não-tegumentares são abundantes em alguns vertebrados existentes, têm um alto potencial de fossilização e podem se redistribuir pelo corpo se perturbados por correntes de fundo, e devem ser considerados em estudos de cor fóssil baseados em melanossomas preservados. Melanossomas não-integumentários e integumentários podem, no entanto, ser discriminados com base em sua geometria e distribuições espaciais.
Results
Abundance of non-integumentary melanosomes
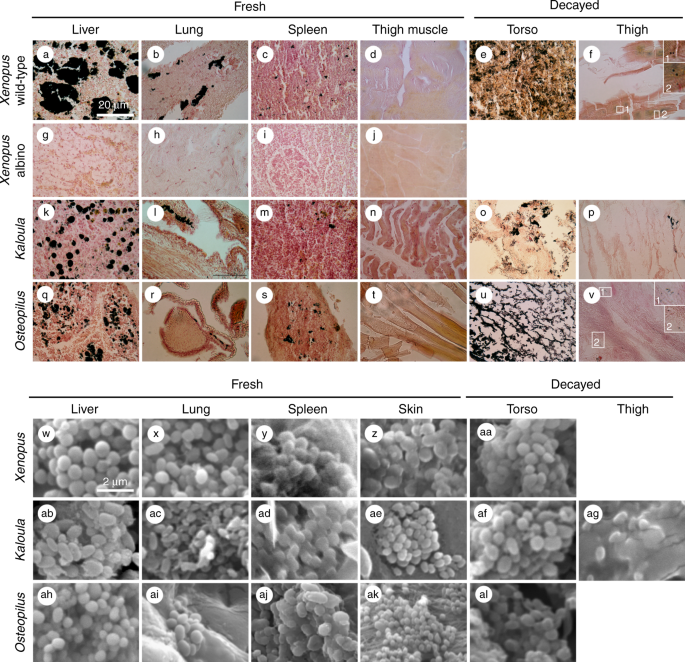
Fontana-Masson histological staining is an established technique for the identification of melanin in histological sections17: melanosomes are stained black, and other tissue components, hues of pink to yellow (Fig. 1a–v).
Fontana-Masson, however, can also stain argentaffin granules, which can
occur in liver tissue and in the epithelial lining of the digestive
tract. To discriminate between melanosomes and argentaffin granules, we
prepared histological sections of tissues from an albino specimen of Xenopus laevis (albino animals lack melanosomes but can possess argentaffin granules18). The results (Fig. 1g–j)
reveal no evidence for argentaffin granules. The regions stained black
in the histological sections of the wild-type (melanin-bearing)
specimens thus correspond to melanin.
Fig. 1
Melanosomes in extant frogs. a–v Histological sections stained with Fontana-Masson; melanosomes (and aggregates of melanosomes) appear black. Insets in f and v show isolated melanosomes. w–al Scanning electron micrographs. Scale bar in a, 20 µm, and same scale in a–v; scale bar in w, 2 µm, and same scale in w–al
Histological
sections and scanning electron microscopy of tissue samples confirm
that melanosomes are present in the lung, liver and spleen (but not the
thigh; Fig. 1d, n, t) of all three extant taxa (Fig. 1a–c, k–m, q–s). These data are supported by AHPO analyses of X. laevis
tissue, which confirm the presence of pyrrole-2,3,5-tricarboxylic acid
(PTCA), pyrrole-2,3-dicarboxylic acid (PDCA) and
pyrrole-2,3,4,5-tetracarboxylic acid (PTeCA) in the liver, lung and
spleen (Table 1). These are diagnostic biomarkers for eumelanin11,19,20. 4-Amino-3-hydroxyphenylalanine (4-AHP), a specific biomarker for pheomelanin21,
was also detected after hydroiodic acid hydrolysis. The amounts of
4-AHP are much lower than those of PTCA and PTeCA, indicating that the
melanin produced in those internal organs is eumelanic.
Table 1 AHPO quantification of melanin markers in extant and fossil frogs
Volumes
of non-integumentary melanosomes were calculated as follows. First, the
volume of each organ was measured in the laboratory. The volume of
melanosomes in each organ was then estimated using histological
sections, whereby the area of tissue occupied by melanin in each section
was converted to a volume using the known thickness of the histological
sections; see Methods). Volumes of non-integumentary melanosomes are
1.7 cm3 for Xenopus, 0.36 cm3 for Kaloula and 0.01 cm3 for Osteopilus (Table 2);
these values correspond to 3.9%, 0.6% and 0.1% of body volume,
respectively. For all taxa, melanosomes are most abundant in the liver
(84–98% of total melanosomes) and least abundant in the spleen
(0.0005–1.45% of total melanosomes; Table 2).
Table 2 Abundance and volume of melanosomes in extant Xenopus, Kaloula and Osteopilus
Models
of integumentary colour in fossils should consider the potential of
contributions from non-integumentary melanosomes if the latter are
abundant in extant vertebrates. The abundance of non-integumentary
melanosomes was assessed as follows. The layer of non-integumentary
melanosomes in the Libros frogs has a mean thickness of ca. 25 µm
(measured in SEM from polished vertical sections)12. The volume of melanosomes required to produce a layer of this thickness over the body outline is 0.126 cm3 for Xenopus, 0.038 cm3 for Kaloula and 0.028 cm3 for Osteopilus (Table 2). Remarkably, Xenopus and Kaloula
therefore possess sufficient non-integumentary melanosomes to define a
layer over the entire body that is 338 μm thick and 234 μm thick,
respectively, i.e., far in excess of the thickness of the layer in the
fossils. Osteopilus has sufficient non-integumentary melanosomes
to define a 25-μm-thick layer over the torso, but not the torso plus the
limbs. In extant frogs, integumentary melanosomes typically form patchy
layers <10 clusters="" dominant="" in="" integumentary="" m="" melanophores="" melanosomes="" occur="" of="" often="" source="" sup="" the="" thick="">9
. Thus, in the fossil frogs, and in extant Xenopus and Kaloula, non-integumentary melanosomes vastly outnumber those from the skin and should dominate the preserved melanosomes.
The fate of non-integumentary melanosomes during decay
Scanning
electron micrographs of the tissues of the experimentally degraded
frogs reveal abundant rod-shaped and spherical to oblate microstructures
(Figs. 1aa, af, ag, al and 2a–c).
The rod-shaped microstructures are interpreted as decay bacteria as no
rod-shaped structures were identified in the tissues in untreated
specimens. The spherical to oblate microstructures, however, are not
definitively coccoid bacteria, as they and melanosomes cannot be
discriminated in scanning electron micrographs. In fluorescence images
of the degraded tissues, rod-shaped microstructures stain green (Fig. 2d–f) and can be identified as bacteria (in stationary growth phase) on the basis of their fluorescence response22,
rod-shaped morphology, size (1.5–2.5 µm long), spatial association with
identical rod-shaped microstructures that stain orange-red
(representing bacteria in active growth phase), and location within
regions of filamentous material, presumably bacterial extracellular
polymeric saccharides. The spherical, micron-sized structures in the
fluorescence images comparable to those visible under SEM quench
fluorescence and appear dark (Fig. 2d–f), as with melanosomes in extant animals23,21,25.
This plus the melanin-rich nature of our tissue samples and the
histological evidence for melanosomes in the degraded tissues from the
torso (Fig. 1e, o, u) and thigh (Fig. 1f, p, v),
the dark microstructures in the fluorescence images can be most
parsimoniously interpreted as melanosomes. Melanin can fluoresce when
strongly oxidized using hydrogen peroxide25,26, but that procedure was not used in this study.
Fig. 2
Decayed tissues of extant frogs. a–c Scanning electron micrographs of rod-shaped decay bacteria within internal tissues of the torso in Kaloula (a), Xenopus (b) and Osteopilus (c). d–f Fluorescent micrographs of melanosomes (d) and rod-shaped bacteria (e) in the torso, and of rod-shaped bacteria in the thigh (f). Melanosomes appear black in fluorescent images. Bacteria appear green and red. Scale bars, 2 μm (a–c), 20 μm (d–f)
Our
laboratory experiments thus confirm that non-integumentary melanosomes
survive the decay process; as with melanosomes from the skin, they show
no visible morphological alteration during decay. Our results also show
that melanosomes usually remain dispersed through, but inside, in the
torso during early decay. After 12 weeks, non-integumentary melanosomes
are abundant in degraded tissue from the torso (Fig. 1e, o, u, aa, af, al). They also occur, albeit rarely, in the thigh region of all three taxa (Fig. 1f, p, v, aa, ag).
Given that our experiments did not involve agitation or other
disturbance of carcasses, our data suggest that substantial
redistribution of melanosomes in fossils is unlikely where carcasses are
deposited prior to extensive decay and not subjected to disturbance
while resting on the sediment surface, e.g., by bottom currents. More
prolonged pre-depositional decay and/or post-depositional disturbance of
carcasses, however, is likely to result in more extensive
redistribution of melanosomes inside the body. This hypothesis can be
tested using fossil material (see below).
Non-integumentary melanosomes in fossil amphibians
Previous
study of the soft tissues of frogs from the Late Miocene Libros biota
revealed the presence of carbonaceous microbodies (Fig. 3g–i)12. The microbodies were originally interpreted as fossil bacteria12, but reinterpreted as fossil melanosomes based on geochemical data13.
This interpretation is further supported herein by our AHPO analysis,
which confirms the presence of PTCA and PTeCA (a marker for
diagenetically altered eumelanin11) in the fossil soft tissues (Table 1).
Fig. 3
Fossil amphibian soft tissues. a–c Frogs from Libros (Miocene, Spain) (a MNCN 63776, Museu Nacional de Ciencias Naturales, Madrid), Bechlejovice (Oligocene, Czech Republic) (b NMP 39449, National Museum Prague), and Messel (Eocene, Germany) (c
SMF-ME-00978, Forschungsinstitut und Naturmuseum Senckenberg,
Frankfurt). Torsos show dark patches of non-integumentary melanosomes.
Inset in b shows melanosomes from the torso. d, e Tadpoles from Libros. Lungs are dark patches in the torso (d MNCN 63848); the notochord, two closely spaced parallel lines (d, e NHM 49999, Natural History Museum, London, UK). f
Salamander from Daohuguo (Middle Jurassic; CNU-V-1264, Capital Normal
University, Beijing, China). Torso shows conspicuous dark patches. g–j Scanning electron micrographs of melanosomes in frogs (g–i) and tadpoles (j) from Libros. g Non-integumentary melanosomes. h Integumentary melanosomes. i, j Size-specific layers of melanosomes. i Integumentary melanosomes (top right) overlying non-integumentary melanosomes (lower left). j
Size-specific layers of non-integumentary melanosomes, reflecting
vertical superposition of different internal tissues in vivo. Scale
bars, 60 mm (a), 50 mm (b), 20 mm (c), 10 mm (d, f), 2 µm (g–j)
The
melanosomes occur as layers in association with phosphatized soft
tissues interpreted as the mid-dermal Eberth–Katschenko (E–K) layer.
This interpretation is discussed in detail in ref.12;
in brief, the fossil layer is identical to the E–K layer of extant
frogs in terms of its anatomical distribution, thickness, collagen-rich
composition, the geometry of its upper and lower surfaces, and the
presence of perforations identical in diameter and geometry to the
centrifugal pillars of the dermis. Selective phosphatization of this
tissue in the fossils reflects its phosphate-rich composition in vivo.
This phosphatized tissue layer is both under- and overlain by layers of
melanosomes (‘internal’ and ‘external’, respectively). These soft tissue
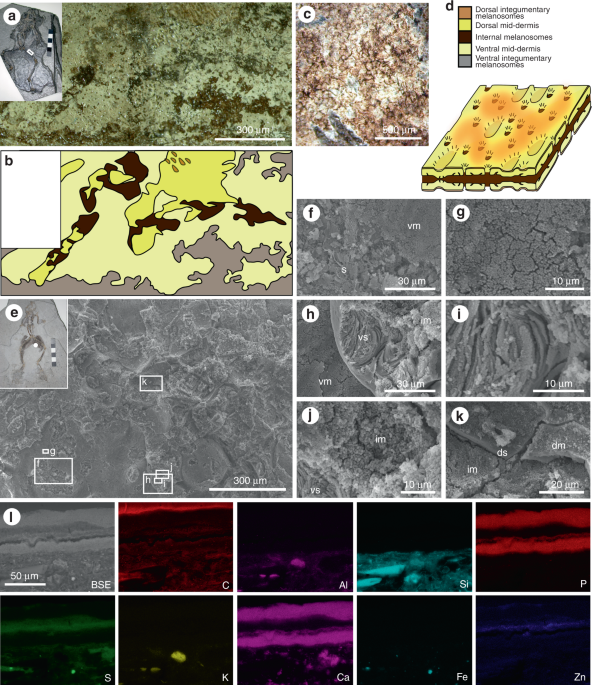
layers have different characteristics in hand specimen (Fig. 4):
the outer melanosome layer is light brown in colour, 2–5 µm thick, can
be patchy or continuous and can preserve patterning (Fig. 4c);
the phosphatized mid-dermis is white to cream in colour, 15–30 µm
thick, and continuous; and the inner melanosome layer is dark brown in
colour, 15–200 µm thick, and continuous. Both the outer melanosome layer
and the phosphatized mid-dermal layer are present on dorsal and ventral
sides of the specimen.
Fig. 4
Layered soft tissues in the hindlimbs of fossil frogs from Libros. a–b MNCN 63775. Light micrograph of area indicated in inset, showing a plan view of the soft tissues in the thigh. b Interpretative drawing of soft tissue layers present, based on light microscopy. c Patterning in the integumentary melanosome layer. d
Three-dimensional schematic illustration of the preserved 3D structure
of the layered soft tissues in the Libros frogs, based on data presented
in ref.12 and this figure. The terms ‘dorsal’ and ‘ventral’ are for illustrative purposes only. e–l MNCN 63798. e Scanning electron micrograph of soft tissues from the region indicated in the inset. f–k Details of regions indicated in e. f Sediment (s) underlying the ventral melanosome layer (vm). g Detail of ventral melanosomes. h Ventral layer of phosphatized skin (vs) overlain by non-integumentary melanosomes (im) and underlain by ventral melanosomes. i Detail of collagen fibres of phosphatized skin. j Detail of non-integumentary melanosomes. k Dorsal layer of phosphatized skin (ds) overlain by dorsal melanosome layer (dm). l
Elemental maps of polished sections through the soft tissues from the
thigh. Melanosome layers are defined by C, S and Zn. SE: secondary
electron micrograph of region analysed, dm: dorsal melanosome layer, s:
sediment, vm: ventral melanosome layer, vs: ventral skin layer. Scale
bars, 300 μm (a), and same scale in b, 500 μm (c), 300 µm (e), 30 µm (f, h), 10 µm (g, i, j), 20 µm (k), 50 μm (l)
Our
results demonstrate that the melanosomes in different layers are from
different tissues. Melanosomes in the outer layer are 0.35–0.5 µm
spheres; the inner layer includes sub-layers comprising 0.9–1.1 µm
ovoids and ca. 0.5 µm spheres, respectively (Fig. 3i)12,13. The outer melanosome layer derives from the upper dermis and epidermis, and the thicker inner layer, from internal tissues.
Other amphibians from Libros and other localities27,28
also exhibit discrete layers of melanosomes. Thin patchy layers over
the body outline represent integumentary melanosomes; non-integumentary
melanosomes form a thicker layer that is usually confined to the torso
(Fig. 3a–f), where they can define the positions of internal organs12,27,28,29. Critically, 93% of the 73 Libros frogs studied possess non-integumentary melanosomes (see Supplementary Data 1) and in 25%, the latter extend from the torso into the limbs29 (Fig. 4).
Thus, it cannot be assumed a priori that melanosomes from the body
outlines of fossil vertebrates represent only integumentary melanosomes.
A mechanism to discriminate integumentary and non-integumentary
melanosomes is required.
Tissue-specific melanosome geometries
Our results show that melanosomes vary in geometry among body tissues within individual taxa (Table 3); except for the liver, lung and spleen of Osteopilus (which have similar geometries), these differences are statistically significant (Table 4).
Critically, differences in the sizes of melanosomes from the skin and
the liver (the primary source of melanosomes) are statistically
significant for all three taxa (Table 4).
We predict that collapse of soft tissues during decay would generate
diagnostic spatial distributions of melanosomes, typically a thick layer
of melanosomes sandwiched between thin layers of melanosomes of
different geometry; where melanosomes have not redistributed, limbs
should show a thin layer of melanosomes of uniform size. Fossil frogs
from Libros exhibit the predicted vertical partitioning of melanosomes
(Fig. 3g–i): non-integumentary and integumentary melanosomes differ significantly in geometry (Student’s t-test p = 3.42e−13 (length); p = 7.96e−8 (aspect ratio)); non-integumentary melanosomes from adjacent layers differ in geometry (p = 1.67e−17 (length); p = 1.28e−8 (aspect ratio)).
Table 3 Geometry of melanosomes in extant Xenopus, Kaloula and Osteopilus
This
study highlights a potential risk in the use of fossil melanosomes to
infer original coloration in ancient animals. Melanosomes preserved
within the body outline of a fossil animal may not have imparted
integumentary coloration.
Contrary to suggestions that melanosomes decay rapidly5,
our experiments confirm that vertebrate melanosomes routinely survive
decay. Analysis of modern frogs shows that non-integumentary melanosomes
are highly abundant and most remain in situ during early decay provided
carcasses are not disturbed. Many fossils, especially those deposited
rapidly after death in a quiescent environment, and especially those
buried soon after death, may thus be expected to retain original
distributions of non-integumentary melanosomes. Our fossil analysis
demonstrates, however, that under certain circumstances,
non-integumentary melanosomes can redistribute from the torso into the
limbs; many frogs from Libros show evidence of disturbance by weak
currents during residence on the lake floor, viz. disarticulation of
small distal limb bones and association of fossils with plant fragments29.
Redistribution should thus be anticipated in contexts where the
biostratinomy of the fossils or the sedimentological context indicates
current activity.
Even if they remain in situ, melanosomes
preserved in the torso will derive from both the internal organs and the
skin and thus cannot be assumed to reflect integumentary pigmentation.
Compounding this, the plane of splitting typically passes through
preserved soft tissues (as evidenced by similar distributions of soft
tissues on both part and counterpart of a specimen; Fig. 5a–f),
thus exposing non-integumentary melanosomes on the surface of a fossil.
Importantly, preservation of non-integumentary melanosomes does not
necessarily preclude preservation of original integumentary colour
patterning (Fig. 4c),
especially where melanosomes have not redistributed and in body regions
where redistribution is unlikely. The latter includes feathers that do
not overlap the torso, in particular where melanosomes are localized to
the barbules and are embedded in an organic matrix, i.e., the degraded
remains of the feather keratin. Studies of fossil melanin should
consider the possibility that variations in tone in fossil soft tissues
comprising melanosomes may reflect the positions of internal organs
where melanosomes have remained in situ, or the presence of layers of
melanosomes redistributed from internal organs, not integumentary
patterning.
Fig. 5
Distribution of soft tissues in part and counterpart of fossil amphibian specimens. In a–f
the plane of splitting passes medially through the soft tissues, thus
exposing non-integumentary melanosomes at the surface and producing
near-identical distributions of soft tissues in part and counterpart. In
g, the dissimilar distribution of soft tissues in part and
counterpart demonstrates that the plane of splitting is not precisely
medial within the soft tissues; non-integumentary melanosomes may still
be exposed at the surface. a MNCN 63781. b MNCN 63848. c MNCN 63793. d MNCN 63864. e IPS 16465, Institut de Paleontología de Sabadell Miquel Crusafont, Sabadell, Barcelona, Spain. f MGB-33179a, Museu de Geologia de Barcelona, Barcelona, Spain. g MPV-1934-RMa, Museu de Paleontología de Valencia, Valencia, Spain. a–f are larval P. pueyoi from Libros; g is Chelotriton from the Middle Miocene of Rubielos de Mora, Spain. Scale bars, 10 mm
Our
results show that melanosomes from the internal organs and skin in
extant frogs can, in some instances, be discriminated based on geometry.
Sympathetic to this, the fossils studied show vertical partitioning of
melanosomes into size-specific layers. These observations form a basis
for assessing the suitability of fossil specimens for studies of
integumentary coloration. Ideal specimens are those that retain original
spatial distributions of melanosomes, evident in SEM images as vertical
and lateral separation of melanosomes of different geometries within
the preserved soft tissues. Such specimens will characteristically show
definition of internal organs by dark tones in hand specimen and offer
the best opportunity to prevent erroneous sampling of non-integumentary
melanosomes. Layering of melanosomes has also been identified in the
eyespots of fossil animals and interpreted as a taxonomic signal30.
A single visually uniform layer of melanosomes (potentially with mixing
of melanosomes of different size and geometry) in fossil specimens,
however, may be original or result from melanosome dispersal.
Diverse
fossil vertebrate taxa preserve visual evidence for non-integumentary
melanosomes and thus our results have broader applications within
vertebrates, including taxa that possess feathers or hair. Examples
include the conspicuous dark patch in the anterior torso of the
ichythyosaur Stenopterygius quadriscissus31 and dark patches in the torso of the bat Palaeochiropteryx tupadiodon32. Numerous examples in fossils from the Jehol biota include the dark patches in the torso of Sinosauropteryx prima33, Sinornithosaurus34 and Epidexipteryx35. Preliminary analyses of extant reptiles confirm that melanosomes are abundant in various internal tissues (Fig. 6).
Future studies of diverse extant vertebrates will characterize the
preservation potential, abundance and geometry of non-integumentary
melanosomes, and will test the feasibility of other mechanisms to
identify the source of melanosomes, e.g., the chemistry of the
melanosomes and their embedding matrix. Discriminating between the
melanosomes from different sources is essential to accurate
reconstruction of melanin-based pigmentation in fossil vertebrates.
Fig. 6
Melanosomes in extant reptiles. Scanning electron micrographs of extant black iguana (Ctenosaura similis (Squamata: Corytophanidae), a–c) and the common basilisk (Basiliscus basiliscus (Squamata: Iguanidae), d–f). a, d Connective tissue. b, e Liver. c Kidney. f Spleen. Scale bars, 5 μm
The morphological decay of frogs has been studied in detail previously36.
Our laboratory experiments had two objectives: (1) to assess whether
non-integumentary melanosomes survive early decay, and (2) to assess
whether these melanosomes redistribute within a carcass during early
decay. We conducted a series of pilot experiments using extant frogs to
identify the most appropriate conditions for our experiments; the goal
was not to attempt to replicate the full complexity of natural settings37
but to identify conditions in which carcasses would remain intact
during decay, allowing the decay of melanosomes within the body cavity
to be tracked. Adult specimens of the African-clawed frog, Xenopus laevis (Anura: Pipidae), the Asian-painted frog, Kaloula pulchra (Anura: Microhylidae), and the Cuban tree frog, Osteopilus septentrionalis
(Anura: Hylidae) were euthanased via immersion in water with added
3 g/l tricaine methane sulphonate. The first pilot experiment used
natural lake water plus an inoculum of lake sediment and was carried out
at 25 °C. This experiment was unsuccessful as all carcasses bloated
rapidly due to high rates of gas production by decay bacteria and
ruptured within 96 h, releasing internal tissues (and their melanosomes)
into the decay medium. The second pilot experiment used distilled water
with no sediment and was carried out at 15 °C, but all carcasses
ruptured within 1 week. The third pilot experiment involved freezing
freshly killed carcasses at −80 °C for 24 h in order to inhibit the
growth of (and rate of gas production by) decay bacteria and thus
prevent bloating; carcasses were then decayed as per the second phase of
pilot experiments. Carcasses in this experiment did not rupture.
Dissection of specimens after 3 days revealed that all major organs
showed a similar fidelity to those in specimens that had not been
frozen; there is no evidence that freezing affected the rate of tissue
degradation and/or the redistribution of non-integumentary melanosomes.
The final experiment used five specimens of Xenopus (four wild type, one albino) and two specimens each of Kaloula and Osteopilus.
Frog carcasses were frozen at −80 °C for 48 h, placed in individual
glass vessels containing 650 ml of distilled water and degraded at
15± 1 °C for 12 weeks in a Memmert IPS260 constant temperature
incubator.
Reptiles for scanning electron microscopy (see below) were supplied as carcasses.
Ethics
The
authors have complied with all relevant ethical regulations. Euthanasia
of frogs was approved by the University of Bristol Animal Ethics
Committee via authorization UIN/13/017 and carried out by the University
of Bristol Animal Services Unit.
Histology
The abundance of melanin in internal organs was assessed using histological sections. Small (ca. 5 mm3)
samples of liver, lung and spleen were dissected from freshly killed
frog specimens and the volume of each organ (and that of the entire
frog) calculated by immersion in water. Samples of degraded tissue were
also removed from within the abdominal body cavity of degraded
specimens. Tissue samples were fixed in 10% neutral buffered formalin
overnight and dehydrated in the following ethanol: water mixtures:
distilled water (3 × 20 min), 30% ethanol (20 min), 50% ethanol
(20 min), 70% ethanol (20 min), 90% ethanol (20 min) and 100% ethanol
(2 × 20 min). Dehydrated samples were immersed in histolene-clearing
agent overnight and embedded in paraffin wax. Histological sections
(9 μm thick) were stained using Fontana-Masson (FM) using standard
protocols17.
Image analysis
In
histological sections stained using Fontana-Masson, melanosomes appear
black. In order to assess the abundance of melanosomes in each organ,
the area of tissue occupied by melanosomes was calculated by analyzing
digital images of histological sections using the Threshold Colour
function in ImageJ (www.imagej.nih.org).
Ten images were analyzed for each tissue type in each taxon. These data
were used to estimate the volume of melanosomes in each organ based on
the measured organ volume (see above) and thickness of histological
sections (9 µm).
The mean thickness of the internal melanosome layer in fossil frogs from Libros is 25 μm12.
The volume of a 25-µm-thick layer corresponding to the torso, and
entire body, respectively, was calculated for each extant taxon.
Fluorescence microscopy
Acridine
orange staining is an established technique for discrimination of
bacterial cells and eukaryotic tissue in histological sections17,38. Eukaryotic cells fluoresce green and prokaryotic cells fluoresce red-orange19,38.
The staining response of bacteria, however, varies with physiology:
active bacteria fluoresce red, and those in the stationary growth phase
fluoresce green19,39. Sections of stained decayed tissue were stained using Acridine Orange using standard protocols17,38
and analyzed using an Olympus BX53 upright fluorescence microscope
(×100 objective, N.A. 1.4) with filter cube sets for green (Exciter
filter 470–495, DM505, Barrier filter 510–550 nm) and blue images
(Exciter filter 540–550, DM570, Barrier filter 576–625 nm).
Scanning electron microscopy (SEM)
Samples of fresh and decayed tissue were prepared using standard protocols40
and examined using a Hitachi S-3500N variable pressure SEM at an
accelerating voltage of 15 kV. Long and short axes were measured for
melanosomes from the liver, lung, spleen and skin in the extant frogs,
and from melanosomes in the two layers in the fossils.
Alkaline hydroxide peroxide oxidation (AHPO)
Alkaline
hydrogen peroxide oxidation is a unique chemical assay for melanin that
produces diagnostic chemical markers (e.g., pyrrole-2,3,5-tricarboxylic
acid (PTCA), pyrrole-2,3-dicarboxylic acid (PDCA) and
pyrrole-2,3,4,5-tetracarboxylic acid (PTeCA), which are derived from the
dihydroxyindole parent subunit of the melanin molecule) that allow
identification and quantification of melanin in modern and fossil
materials11,19. Freeze-dried samples of liver, lung and spleen of X. laevis
(9–17 mg) were homogenized in water with a Ten-Broeck homogenizer at a
concentration of 10 mg/ml and 200 µl aliquots were dried in a dessicator
and subjected to acid hydrolysis with 6 M HCl (0.5 ml) at 110 °C for
16 h40.
This acid hydrolysis removes proteins and small molecules that might
interfere with the assay and thus increases specificity of the
biomarkers40. The resulting insoluble materials including melanin were collected by centrifugation and washed once with water as described40. Then the residues were subjected to alkaline hydrogen peroxide oxidation (AHPO)38. Aliquots (100 µl) of X. laevis
suspensions were also subjected to hydroiodic acid hydrolysis to
analyze 4-amino-3-hydroxyphenylalanine (4-AHP), a specific biomarker for
phaeomelanin39.
A sample of soft tissues from a fossil frog from the Late Miocene
Libros biota (MNCN 63776) was finely ground with a mortar and pestle and
weighed. The powder was subjected to acid hydrolysis and then AHPO as
above.
Schweitzer, M. H., Lindgren, J. & Moyer, A. E. Melanosomes and ancient coloration re-examined: a response to Vinther 2015. Bioessays37, 1174–1183 (2015).
Sichel,
G., Scalia, M., Mondio, F. & Corsaro, C. The amphibian Kupffer
cells build and demolish melanosomes: an ultrastructural point of view. Pigment Cell Res.10, 271–287 (1997).
McNamara,
M. E. et al. Soft tissue preservation in Miocene frogs from Libros
(Spain): insights into the genesis of decay microenvironments. Palaios24, 104–117 (2009).
McNamara,
M. E., van Dongen, B. E., Lockyer, N. P., Bull, I. D. & Orr, P. J.
Fossilization of melanosomes via sulfurization. Palaeontology59, 337–350 (2016).
Hu,
D. et al. A bony-crested Jurassic dinosaur with evidence of iridescent
plumage highlights complexity in early paravian evolution. Nat. Commun.9, 217 (2018).
Ito,
S. et al. Usefulness of alkaline hydrogen peroxide oxidation to analyze
eumelanin and pheomelanin in various tissue samples: application to
chemical analysis of human hair melanins. Pigment Cell Melanoma Res.24, 605–613 (2011).
Ito, S. et al. Acid hydrolysis reveals a low but constant level of pheomelanin in human black to brown hair. Pigment Cell Melanoma Res.31, 393-403 (2018).
21.
Wakamatsu, K., Ito, S. & Rees, J. L. Usefulness of 4-amino-3-hydroxyphenylalanine as a specific marker of pheomelanin. Pigment Cell Res.15, 225–232 (2002).
McFeters,
G., Singh, A., Byun, S., Callis, P. & Williams, S. Acridine orange
staining reaction as an index of physiological activity in Escherichia coli. J. Microbiol. Methods13, 87–97 (1991).
Eldar,
A., Bejerano, Y., Livoff, A., Horovitcz, A. & Bercovier, H.
Experimental streptococcal meningo-encephalitis in cultured fish. Vet. Microbiol.43, 1–102 (1995).
Ancans,
J. et al. Melanosomal pH controls rate of melanogenesis,
eumelanin/phaeomelanin ratio and melanosomes maturation in melanocytes
and melanoma cells. Exp. Cell Res.286, 26–35 (2001).
Jarrett, I. & Spearman, R. J. The keratin defect and hair-cycle of a new mutant (matted) in the house-mouse. J. Embryol. Exp. Morphol.5, 103–110 (1957).
Fuller,
B., Spaulding, D. & Smith, D. R. Regulation of the catalytic
activity of preexisting tyrosinase in black and Caucasian human
melanocyte cell cultures. Exp. Cell Res.262, 197–208 (2001).
McNamara,
M. E. et al. Exceptionally preserved tadpoles from the Miocene of
Libros, Spain: ecomorphological reconstruction and the impact of
ontogeny upon taphonomy. Lethaia43, 290–306 (2010).
Špinar, Z. Tertiary Frogs from Central Europe (Academia, Prague, 1972).
29.
McNamara,
M. E. et al. What controls the taphonomy of exceptionally preserved
taxa – environment or biology? A case study using exceptionally
preserved frogs from the Miocene Libros Konservat-Lagerstätte, Spain. Palaios27, 63–77 (2012).
Zhang,
F., Zhou, Z., Xu, X., Wang, X. & Sullivan, C. A bizarre Jurassic
maniraptoran from China with elongate ribbon-like feathers. Nature455, 1105–1108 (2008).
We
thank the University of Bristol Animal Services Unit for supply and
euthanasia of frogs, and Suzanne Crotty, John Cunningham, Grace
Flannery, Stuart Kearns, Sharon Lynch, Steve Martin, Chris Rogers and
Richard Wall for technical assistance. The research was funded by
Enterprise Ireland Enterprise Ireland Basic Research Grant C/2002/138
awarded to P.J.O., NERC Standard Grant NE/I027630/1 awarded to M.J.B.
and by Marie Curie Career Integration Grant PCIG13-GA-2013-618598 and
European Research Council Starting Grant 2014-ERC-STG-637691 awarded to
M.E.M.
Author information
Affiliations
School of Biological, Earth and Environmental Sciences, University College Cork, North Mall, Cork, T23 TK30, Ireland
Maria E. McNamara
& Valentina Rossi
School of Earth Sciences, University of Bristol, Queen’s Road, Bristol, BS8 1RJ, UK
Jonathan S. Kaye
& Michael J. Benton
UCD School of Earth Sciences, University College Dublin, Belfield, Dublin, D04D1W8, Ireland
Patrick J. Orr
Department of Chemistry, Fujita Health University School of Health Sciences, Toyoake, Aichi, 470-1192, Japan
Shosuke Ito
& Kazumasa Wakamatsu
Contributions
M.E.M.
designed the research; J.K. performed histology and statistical
analyses; J.K., M.E.M. and V.R. performed SEM and fluorescence analyses;
S.I. and K.W. performed AHPO analyses; M.E.M., J.K., M.J.B. and P.J.O.
wrote the paper. All authors discussed the results and provided input on
the manuscript.
Open Access
This article is licensed under a Creative Commons Attribution 4.0
International License, which permits use, sharing, adaptation,
distribution and reproduction in any medium or format, as long as you
give appropriate credit to the original author(s) and the source,
provide a link to the Creative Commons license, and indicate if changes
were made. The images or other third party material in this article are
included in the article’s Creative Commons license, unless indicated
otherwise in a credit line to the material. If material is not included
in the article’s Creative Commons license and your intended use is not
permitted by statutory regulation or exceeds the permitted use, you will
need to obtain permission directly from the copyright holder. To view a
copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.




 Open Access
This article is licensed under a Creative Commons Attribution 4.0
International License, which permits use, sharing, adaptation,
distribution and reproduction in any medium or format, as long as you
give appropriate credit to the original author(s) and the source,
provide a link to the Creative Commons license, and indicate if changes
were made. The images or other third party material in this article are
included in the article’s Creative Commons license, unless indicated
otherwise in a credit line to the material. If material is not included
in the article’s Creative Commons license and your intended use is not
permitted by statutory regulation or exceeds the permitted use, you will
need to obtain permission directly from the copyright holder. To view a
copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Open Access
This article is licensed under a Creative Commons Attribution 4.0
International License, which permits use, sharing, adaptation,
distribution and reproduction in any medium or format, as long as you
give appropriate credit to the original author(s) and the source,
provide a link to the Creative Commons license, and indicate if changes
were made. The images or other third party material in this article are
included in the article’s Creative Commons license, unless indicated
otherwise in a credit line to the material. If material is not included
in the article’s Creative Commons license and your intended use is not
permitted by statutory regulation or exceeds the permitted use, you will
need to obtain permission directly from the copyright holder. To view a
copy of this license, visit http://creativecommons.org/licenses/by/4.0/.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.