Jacaré extinto gigante quebra restrição no esqueleto axial de crocodilianos existentes
- Universidade de Zurique, Instituto e Museu Paleontológico, Suíça
- Departamento de Ciências Biomédicas Comparadas, The Royal Veterinary College, Reino Unido
- Dipartimento de Scienze della Terra, Universidade de Turim, Itália
- Instituto Catalão de Paleontologia Miquel Crusafont, Universitat Autònoma de Barcelona, Espanha
- Museu Paleontológico de Urumaco, Venezuela
- Citado 0
- Visualizações 1326
- Anotações
Citar
como: eLife 2019; 8: e49972 doi: 10.7554 / eLife.49972
Abstrato
O número de vértebras de precaução em todos os crocodilianos existentes
é notavelmente conservador, com nove cervicais, 15 dorsais e dois
sacrais, padrão presente também em seus parentes extintos mais próximos. A contagem vertebral consistente indica um controle rígido do padrão axial pelos genes Hox durante o desenvolvimento. Aqui, relatamos um desvio desse padrão com base em um esqueleto associado do gigante jacaré Purussaurus , um membro da coroa Crocodylia, e vários outros espécimes do Neogene dos neotrópicos do norte. P. mirandai
é o primeiro coroa-crocodyliano a ter três sacrais, duas vértebras
sacrais verdadeiras e uma dorsossacral não patológica e funcional, a
articular-se com o ílio (pelve). O tamanho do corpo gigante deste jacaré está relacionado a alterações locomotoras e posturais.
A configuração iliossacral, uma cinta peitoral mais orientada
verticalmente e a baixa torção da cabeça femoral em relação aos côndilos
são especializações hipotetizadas para orientação mais vertical dos
membros ou suporte de peso.
eLife digest
Todos os crocodilos, jacarés, jacarés e gharials vivos - coletivamente
chamados crocodylians - têm um plano corporal semelhante que se adapta
ao seu estilo de vida como predadores de emboscadas aquáticas. Essa semelhança se estende por todo o corpo, incluindo o esqueleto. Suas espinhas dorsais, por exemplo, sempre têm nove vértebras no pescoço, 15 no tronco e duas na pelve. Crocodylianos extintos intimamente relacionados também organizam seus espinhos da mesma maneira.
Scheyer et al., No entanto, agora relatam que um jacaré extinto chamado Purussaurus mirandai tinha uma coluna vertebral estruturada de maneira diferente de qualquer outro crocodyliano conhecido. Pesando quase três toneladas (~ 2.600 kg), o Purussaurus de 10 metros de comprimento era mais do que o dobro do peso do maior crocodiliano vivo, o crocodilo de água salgada. Quando Scheyer et al.
Ao examinar restos fossilizados da Venezuela, estimados entre 7 e 5
milhões de anos, encontraram uma vértebra extra na área pélvica da
criatura e uma a menos em seu tronco. Scheyer et al.
especula-se que esse arranjo incomum possa ter ajudado a criatura
extinta a suportar seu peso maciço e compensar a tensão imposta ao seu
esqueleto.
Dentro do reino animal, os chamados genes da homeobox determinam como
diferentes estruturas do corpo, incluindo a coluna, se desenvolvem nos
embriões. Mudanças nas quais esses genes são ativos no embrião podem resultar em uma vértebra pélvica extra em humanos e outros animais. Scheyer et al.
Concluímos que mudanças nos limites da atividade dos genes da homeobox
também podem explicar a vértebra pélvica extra nesse jacaré antigo.
Ainda não está claro se outros crocodilianos extintos também tinham vértebras pélvicas extras.
Mas essas novas descobertas provavelmente levarão a mais pesquisas
sobre fósseis gigantes crocodylianos relacionados a serem descobertos.
Essa pesquisa pode ajudar os cientistas a entender melhor a biomecânica
dos crocodilianos e pode levar a novas idéias sobre os jacarés, que
prosperaram nos trópicos do norte da América do Sul nos últimos sete
milhões de anos.
Outras pesquisas nessa área também podem ajudar a explicar como esses
répteis se adaptaram ao ambiente e o papel que desempenham em seus
ecossistemas, atualmente ameaçados pela atividade humana.
Introdução
A fauna crocodiliana Neogene do norte da América do Sul é notável em
termos de riqueza de espécies, níveis de simpatia e especialização
ecomorfológica (por exemplo, Riff et al., 2010 ; Sánchez ‐ Villagra e Aguilera, 2006 ; Scheyer et al., 2013 ; Scheyer e Delfino, 2016 ). Entre uma infinidade de táxons, o Purussaurus
era um jacaré excepcionalmente grande (Alligatoroidea) que vivia nos
neotrópicos do norte da América do Sul no Mioceno médio e tardio (ca.
13–5 Ma).
Seu crânio impressionantemente grande é a base de sua taxonomia, que
abrange três espécies distribuídas em localidades no Brasil, Peru,
Colômbia e Venezuela ( Aguilera et al., 2006 ; Langston, 1965 ; Mook, 1921a ; Mook, 1942 ; Riff et al. , 2010 ; Salas-Gismondi et al., 2015 ). A anatomia pós- craniana do Purussaurus , como a da maioria dos crocodilianos extintos, ainda é pouco conhecida. Relatamos aqui a descoberta de um esqueleto excepcional de Purussaurus mirandai e vários outros restos do Mioceno na Venezuela.
Os archosauria são representados hoje por mais de 10.000 espécies de
aves (coroa Aves), mas menos de 30 espécies de crocodylians (coroa
Crocodylia).
Como taxa irmã, ambos os grupos sofreram profundas mudanças em seus planos corporais ao longo de ca. 250 milhões de anos de história evolutiva.
Ao comparar a morfologia da Archosauria existente, as aves revelam uma
variação muito maior na forma do corpo, forma e função
musculoesquelética, ecologia e estilo de vida;
enquanto as espécies crocodylianas se assemelham muito mais devido ao
estilo de vida anfíbio e carnívoro em geral compartilhado.
Ao examinar os padrões de desenvolvimento subjacentes nos arquossauros,
a diferença na variação também se reflete no padrão axial da coluna
vertebral em Aves e Crocodylia, com o primeiro tendo números vertebrais
de precaução variáveis e o segundo mostrando um padrão conservado ( Mansfield e Abzhanov, 2010 ; Müller et al., 2010 ).
Finalmente, os crocodilianos existentes mostram taxas evolutivas
distintamente baixas em todo o genoma em comparação com as aves, o que
pode estar relacionado a tempos de geração prolongados no antigo clado (
Green et al., 2014 ).
Essas baixas taxas evolutivas podem potencialmente estar subjacentes à
disparidade morfofuncional geralmente mais baixa observada nos planos
corporais crocodilianos pós-cretáceos ( Brusatte et al., 2010 ; Stubbs et al., 2013 ).
O padrão axial de vertebrados por meio da expressão do gene Homeobox ( Hox ) tem sido extensivamente estudado em organismos modelo, incluindo o pintinho, desde os anos 80 (ver Favier e Dollé, 1997 ; McGinnis e Krumlauf, 1992 ; Wellik, 2007 para obter uma visão geral).
Por outro lado, o estudo do padrão de desenvolvimento e das expressões
gênicas associadas em crocodylians (entre outros répteis existentes) só
recebeu atenção recentemente ( Böhmer, 2013 ; Böhmer et al., 2015 ; Mansfield e Abzhanov, 2010 ), com um concentre-se no padrão pré-sacral do corpo. Esses estudos de desenvolvimento, bem como a anatomia comparada ( Hoffstetter e Gasc, 1969 ; Mook, 1921b
), corroboram a contagem preventiva geral de toda a Crocodylia da coroa
como constituída por nove cervicais, 15 dorsais (torácica e lombar) e
dois sacrais. Várias condições patológicas foram relatadas usando dissecção clássica (por exemplo, Baur, 1886 ; Baur, 1889 ; Reinhardt, 1873 ; Reinhardt, 1874 ).
Aqui, apresentamos todo o material axial e apendicular relevante do caimanino gigante extinto Purussaurus mirandai
, incluindo uma pontuação revisada de caracteres e análise filogenética
para as espécies, como o primeiro caso não patológico na
coroa-Crocodylia que se desvia da contagem prudencial altamente
conservada de o grupo.
A comparação com os crocodilianos existentes patológicos (por exemplo, a
presença de vértebras transitórias lombossacrais de malformação
congênita) e crocodilianos não patológicos serviu de base para elucidar
padrões de desenvolvimento para a sacralização da última vértebra dorsal
(ou seja, lombar) nas espécies extintas.
Resultados
Paleontologia sistemática
Crocodylia Gmelin, 1789
Alligatoridae Gray, 1844
Caimaninae Brochu, 2003 (seguindo Norell, 1988 )
PURUSSAURUS Barbosa-Rodrigues, 1892
P. mirandai Aguilera, Riff e Bocquentin-Villanueva, 2006
Material de holótipo: UNEFM-CIAAP-1369, compreendendo o crânio e o
material da mandíbula associado, além de um fêmur e ísquio, que segundo Aguilera et al. (2006) foi coletado na localidade de El Hatillo (ver Scheyer e Delfino, 2016 para obter informações sobre a localidade).
Novo material referido: AMU-CURS-541, um esqueleto associado, mas
desarticulado, preservando o material craniano e grande parte do
pós-crânio, que foi incorporado normalmente com o abdome no sedimento. O espécime foi atribuído preliminarmente a Purussaurus cf. P. mirandai ( Scheyer e Delfino, 2016
), mas dada a forma geral e as proporções da mandíbula inferior (veja
abaixo), o contorno lateral levemente ondulado do dentário, a pré-maxila
baixa e o jugal baixo e esbelto (indicando um crânio bastante plano)
perfil) é aqui referido como Purussaurus mirandai . A forma da mandíbula, os dentes e a linha dentária em geral também são congruentes com a do holótipo de P. mirandai . Até o momento, não há indicação para a presença de uma segunda espécie de Purussaurus na Formação Urumaco. O contexto filogenético das espécies entre Caimaninae é mostrado na Figura 1 .
Figura 1 com 1 suplemento ver todos
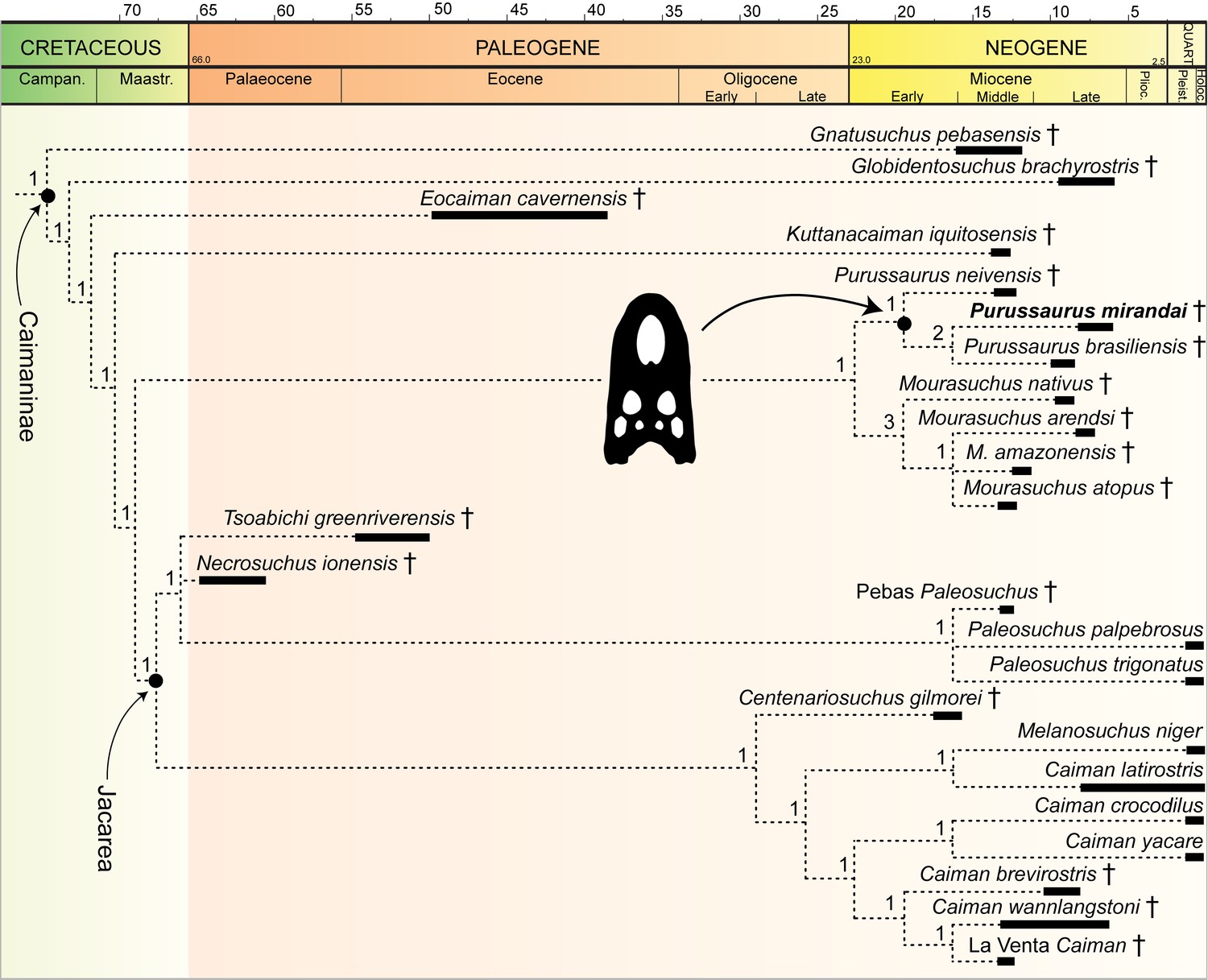
Análise filogenética com pontuação atualizada de Purussaurus mirandai com base no AMU-CURS-541 (consulte Materiais e métodos para explicações).
Aqui, apenas um trecho do clado Caimaninae da estrita árvore de consenso é mostrado como estrutura filogenética de P. mirandai (em negrito). Os valores de suporte da Bremer são fornecidos acima dos ramos. Para o restante da topologia, consulte a seção de resultados e a Figura 1 - suplemento à figura 1 (consulte também Salas-Gismondi et al., 2015 : figura suplementar S6).
Localidade e idade: A amostra provém de sedimentos do Membro Superior
da Formação Urumaco (Mioceno tardio), da localidade 'Norte El Picache'
(11 ° 15 '09,00' 'N; 70 ° 13' 40,00 '' W), Urumaco , Estado de Falcón,
Venezuela.
Sua característica mais recente encontrada na região sacral (junção da
coluna vertebral e da pelve) expande a diversidade morfológica do
esqueleto axial nos crocodilianos da coroa e sugere mudanças no
desenvolvimento correlacionadas às demandas biomecânicas de suporte e
locomoção, provavelmente correlacionadas ao tamanho corporal gigante.
Além disso, inferimos que o tamanho gigantesco de 10 a 12 metros de
comprimento total do corpo e massa relacionada desses animais (ver
também Aureliano et al., 2015 ) influenciaram a morfologia peitoral e pélvica peculiar.
A seguir, fornecemos uma descrição abreviada dos ossos da cintura e da região sacral de P. mirandai pertinentes à discussão ( Figura 2 , Figura 2 - suplemento de figura 1 , Figura 3 ; Figura 3 - suplemento de figura 1 ).
Uma descrição exaustiva de todas as amostras estudadas, incluindo os
ossos pós-cranianos do AMU-CURS-541, será publicada em outro local.
Figura 2 com 1 suplemento ver todos
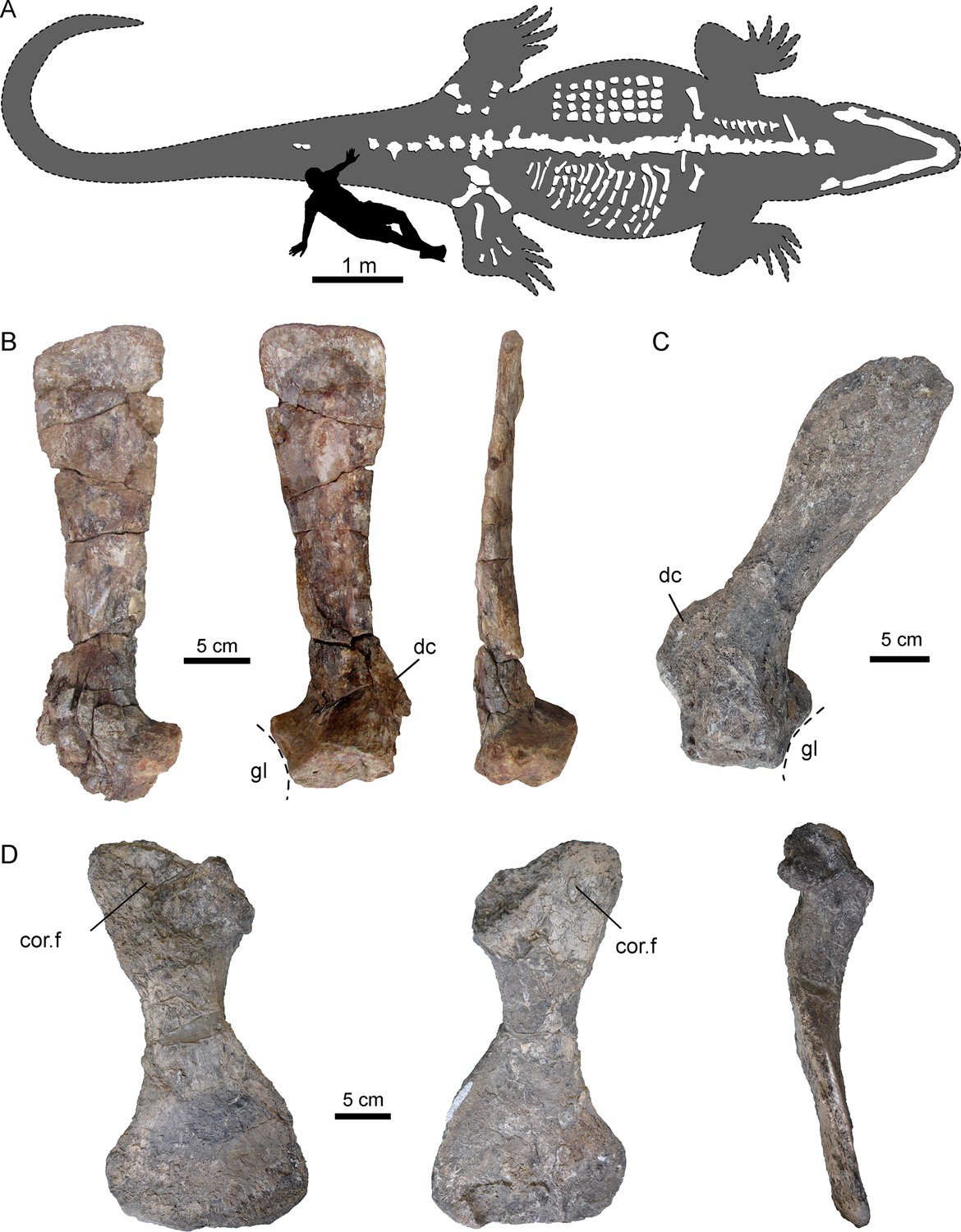
Ossos peitorais selecionados de AMU-CURS-541 e UNEFM-CIAAP-1367 de Purussaurus da Formação Urumaco da Venezuela.
( A ) Reconstrução interpretativa do contorno corporal completo de P. mirandai (AMU-CURS-541) mostrando os ossos pós-cranianos preservados e montados e a mandíbula inferior em posição viva experimental. Osteodermas (na parte superior do tronco) e costelas (na parte inferior do tronco) não estão em posição de vida. O segundo autor (SO) serve como escala (veja a Figura 2 - suplemento da figura 1 ). ( B ) Escápula esquerda do AMU-CURS-541 em vista lateral, medial e posterior. ( C ) Escápula direita do Purussaurus cf. P. mirandai (UNEFM-CIAAP-1367) em vista medial. ( D ) Coracoide direito (UNEFM-CIAAP-1367) em vista dorsomedial, ventrolateral e anterior. Abreviações: cor.f, forame coracoide; dc, crista deltoide da escápula; gl, fossa glenóide.
Figura 3 com 2 suplementos ver todos
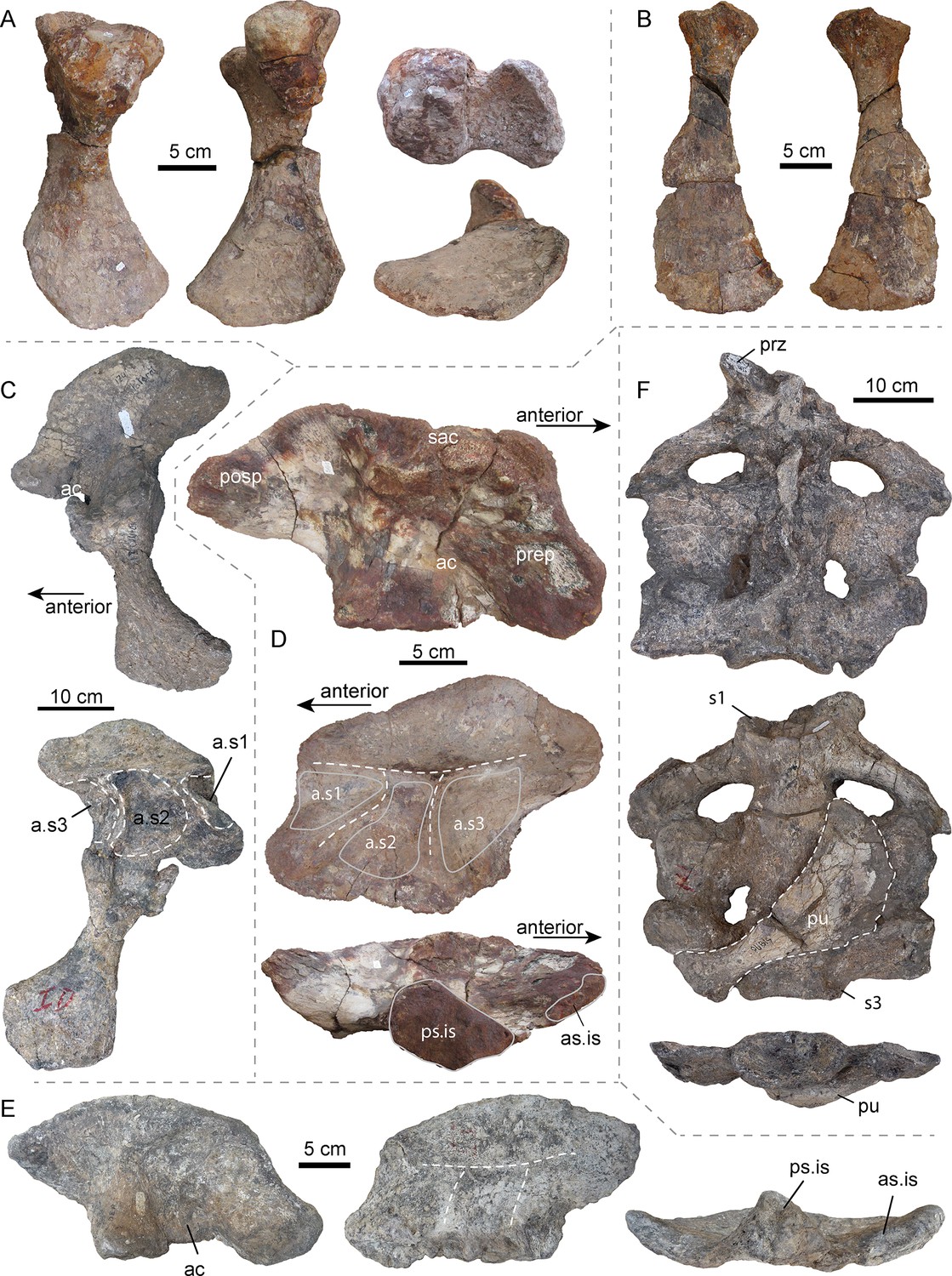
Ossos pélvicos e sacrais selecionados de AMU-CURS-541 e outros espécimes de Purussaurus da Formação Urumaco da Venezuela.
( A ) Ísquio esquerdo (AMU-CURS-541) nas vistas posterior, anterior, dorsal (acima) e ventral (abaixo). ( B ) Púbis direito (AMU-CURS-541) em vista ventral e dorsal. ( C
) ílio esquerdo e ísquio articulados (não holótipo, ossos adicionais
acessados sob UNEFM-CIAAP-1369) em vista lateral e medial. Observe as três grandes depressões separadas por sulcos, ou seja, locais de fixação para três costelas sacrais (a.s1 a a.s3). ( D ) ílio direito (AMU-CURS-541) nas vistas lateral, medial e ventral. ( E ) ílio direito (UNEFM-CIAAP-1367) nas vistas lateral, medial e ventral. Linhas brancas pontilhadas indicam as cristas desgastadas que separam as facetas da articulação das costelas sacrais. ( F
) Região sacral (não holótipo, ossos adicionais acessados sob
UNEFM-CIAAP-1369) consistindo em três sacrais articulados nas vistas
dorsal, ventral e posterior. Observe o púbis direito (em vista dorsal) anexado ventralmente aos sacrais. Abreviações: ac, acetábulo; a.s1-a.s3, locais de fixação para três costelas sacrais; as.is, superfície articular anterior do ílio; posp, processo pós-acetabular; preparação, processo pré-acetabular; prz, prezygapophysis; ps.is, superfície articular posterior para ísquio; pu, púbis; s1-s3: vértebras sacrais / costelas 1–3; saco, crista supraacetabular.
Cintura escapular
Dos elementos da cintura peitoral ( Figura 2
), as escápulas isoladas são conhecidas de duas amostras (AMU-CURS-541;
UNEFM-CIAAP-1367) e um coracóide bem preservado e isolado é conhecido
na UNEFM-CIAAP-1367. Semelhante a jacarés existentes, como Alligator mississippiensis e Caiman crocodilus , Purussaurus mirandai possui lâminas escapulares estreitas.
Em contraste com os táxons existentes, a espécie extinta tem as
escápulas orientadas mais dorsalmente e um pouco posteriormente, bem
como coracóides ventromedialmente e levemente posteriores.
A lâmina longa, plana e estreita, que é desencadeada por uma constrição
da articulação proximal e da fossa glenóide, se estende apenas
levemente em direção à extremidade distal. A crista deltóide (acrômio de Cong, 1998 ) da escápula de Purussaurus
da Formação Urumaco (bem preservada no UNEFM-CIAAP-1367) é um processo
muito robusto e distinto, e de expansão próximo-distal semelhante à
largura da articulação escapular. o coracóide.
O coracóide possui uma haste proeminente e robusta, uma faceta
articular estreita para a escápula, um forame coracóide ovóide próximo à
faceta articular e uma parte distal alargada e alargada.
Cinturão pélvico
Em geral, esses elementos ( Figura 3 ) se assemelham aos de outros crocodilianos em geral, mas também mostram algumas especializações. Como em A. mississippiensis (por exemplo, Brochu, 1999 ), o ílio de P. mirandai (AMU-CURS-541, UNEFM-CIAAP-1369) apresenta um recuo fraco na margem dorsal do processo pós-acetabular. O mais impressionante é que, no entanto, a superfície medial do Purussaurus ilium mostra três concavidades rugosas separadas por cristas formando um 'π' ( Figura 3C-E ), em vez de um 'τ' como nos crocodilianos existentes com duas vértebras sacrais.
Essas concavidades são os locais de fixação das costelas sacrais e
ficam próximas umas das outras, seu tamanho responde à largura
anteroposterior de cada uma das três costelas sacrais, como é visível no
UNEFM-CIAPP 1369. Todas as quatro ilias recuperadas atribuíveis ao Purussaurus do Urumaco A formação mostra os três locais de articulação separados por sulcos ósseos baixos.
Até o momento, há apenas informações comparativas limitadas sobre a
morfologia da cinta pélvica de outras formas crocodylianas gigantes. O ílio direito bem preservado do UCMP 38012 (holótipo) de Mourasuchus atopus de La Venta, Colômbia, no entanto, com 137 mm de comprimento ( Langston, 1965 ); muito menor que o purussauro ; também tem cristas em forma de 'π' em seu lado medial. Dada a falta de outras amostras que corroborem o potencial de três sacrais, tratamos de maneira conservadora a condição em Mourasuchus como inconclusiva.
Em alguns crocodylians mekosuchine (ie, morfotipo descrito como 'formas pélvicas três e quatro'; veja também Figura 3 - suplemento de figura 2
), a separação da forma 'τ' não é óbvia, porque os locais de fixação
das duas costelas sacrais estão mudando separados, criando assim um
espaço intermediário ( Stein et al., 2017 ).
Esse espaço, no entanto, consiste em uma superfície óssea lisa e plana,
que não deve ser confundida com um local de fixação adicional rugoso e
escavado.
O púbis é em forma de machado em P. mirandai, com um eixo delgado e lâmina distal de queima. A superfície óssea proximal do púbis difere da de outros crocodilianos ( Claessens e Vickaryous, 2012 ; Mook, 1921b ; Stein et al., 2017 ), em que a parte lateral da superfície é angulada.
O ísquio é conhecido no holótipo UNEFM-CIAAP-1369 e no material não holótipo de P. mirandai , bem como em AMU-CURS-528 e AMU-CURS-541.
O ísquio possui um eixo delgado e, proximalmente, duas facetas de
articulação do ílio, que formam o acetábulo, e um escudo largo
distalmente alargado. Medialmente, os ísquios têm uma margem de contato direta.
Região sacral
Sacrais isolados são conhecidos nos espécimes AMU-CURS-541 ( Figura 3 - suplemento à figura 1
) e MCNC-URU-111–72V, mas é o sacro articulado completo no
UNEFM-CIAAP-1369 (material não holotípico), constituído por três
vértebras sacrais moderadamente preservadas, mas articuladas, as mais
informativas.
Todas as três vértebras sacrais têm o centro e as costelas sacrais
preservados em articulação, mostrando a morfologia detalhada da cintura
pélvica e a conexão do ílio com as três vértebras sacrais no Purussaurus . As costelas sacrais da primeira vértebra sacral (= dorsossacral) são inclinadas posteriormente.
As costelas sacrais do segundo sacral se estendem lateralmente e as
costelas sacrais do terceiro sacral são inclinadas anteriormente.
Dos três sacrais, as costelas do meio sacral apresentam a maior
expansão ântero-posterior, seguida pela terceira sacral e depois pela
primeira sacral, as menos expandidas.
Com exceção de uma única pré-zigofofia, as outras zigapofises e os
espinhos neurais dos sacrais estão ausentes ou fortemente distorcidos.
Dos sacrais isolados, o terceiro centro
sacral (= sacro primordial 2) do AMU-CURS-541 mostra as superfícies
articulares melhor preservadas, indicando que o côndilo anterior é de
forma redonda e achatada (em vez de fortemente convexa), enquanto o
cotyle posterior é ligeiramente oval e côncavo. O cotyle também mostra uma borda marginal espessa, mas uma flange parece estar ausente ( Figura 3 - suplemento de figura 1D, E e G ). Todas as amostras acima mencionadas são consideradas adultos grandes e variam apenas de forma modesta.
A superfície articular posterior do último centra vertebral da série
sacral (isto é, sacral primordial 2) do material não holotípico acessado
pelo UNEFM-CIAAP-1369 tem 12 cm de largura e 6 cm de altura (a
distância entre as extremidades da costela sacral é de 30 cm ), enquanto
que o sacral primordial 2 do AMU-CURS-541 tem cerca de 12,8 cm de
largura e 8,8 cm de altura (a distância entre as extremidades da costela
sacral é de 28 cm, mas as extremidades não estão completas).
A superfície articular posterior com forte intemperismo da costela
sacral do MCNC-URU-111–72V tem 9 cm de largura e 5,5 cm de altura (as
extremidades da costela sacral estão separadas por 36 cm).
Dadas as margens posteriores fortemente côncavas e a inclinação
posterior de suas costelas sacrais, e em comparação com a série sacral
completa do UNEFM-CIAAP-1369 descrita acima, esse espécime é
identificado como um dorsossacro isolado, que é o primeiro sacral da
série, como bem ( Figura 3 - suplemento de figura 1J – L ).
Análises filogenéticas
A descrição e análise osteológicas forneceram 43 caracteres que
poderiam ser pontuados com base no AMU-CURS-541, com 19 (15
pós-cranianos, quatro mandibulares) novos em comparação à pontuação
disponível anteriormente de P. mirandai ( Salas-Gismondi et al., 2015 )
Análise filogenética com AMU-CURS-541 mesclada com escores de P. mirandai
A análise TNT recuperou as 20 árvores mais parcimoniosas (comprimento
da árvore = 687 passos; IC = 0,383, IR = 0,806), para as quais uma
árvore de consenso rigoroso foi calculada. A análise ( Figura 1 ) mostra a melhor resolução daqueles aqui realizados. Produziu uma topologia semelhante à apresentada por Salas-Gismondi et al. (2015) : fig. 4) com as seguintes exceções: em Gavialoidea, o Gryposuchus pachakamue 'Pebas gavialoid' ( Salas-Gismondi et al., 2016 ) caiu em uma politomia com Siquisiquesuchus , Ikanogavialis , Piscogavialis , Gavialis e um clado de Gryposuchus (consistindo em G. croizati e G. colombianus ). O clado Purussaurus - irmã do gênero Mourasuchus - é totalmente resolvido, sendo P. neivensis a irmã do clado P. mirandai - P. brasiliensis . O clado Jacarea é melhor resolvido em comparação com a análise original de Salas-Gismondi et al. (2015) , mas o apoio de Bremer aos clades de Caimaninae também foi geralmente baixo ( Figura 1 ; Figura 1 - suplemento de figura 1 ).
Análise adicional com AMU-CURS-541 adicionada como taxon terminal separado
Esta análise recuperou 60 árvores mais parcimoniosas (comprimento da
árvore = 687 etapas; índice de consistência CI = 0,383, índice de
retenção RI = 0,806), para as quais foi calculada uma árvore de consenso
estrito. A análise mostra uma topologia semelhante à apresentada no texto principal (ver também Salas-Gismondi et al., 2015 , fig. 4). O agrupamento irmão de P. mirandai e P. brasiliensis não pôde ser recuperado e, em vez disso, o clado Purussaurus entrou em colapso em uma politomia.
Análises adicionais com AMU-CURS-541 mescladas com pontuações de P. mirandai e Melanosuchus fisheri incluíram
Esta análise produziu 160 árvores parcimoniosas (comprimento da árvore = 688 passos; IC = 0,382, IR = 0,805). O estrito consenso dessas árvores mostrou a mesma topologia que em uma análise anterior de Salas-Gismondi et al. (2015) , com exceção da politomia com Siquisiquesuchus , Ikanogavialis , Piscogavialis , Gavialis , um grado de Gryposuchus dentro de Gavialoidea e um clado de Jacarea mal resolvido.
Discussão
Com a descrição do AMU-CURS-541 (incluindo cerca de 100 elementos
preservados) e outras amostras da Formação Urumaco, agora estão
disponíveis novos dados sobre a anatomia pós-craniana do caimanino
gigante Purussaurus mirandai . Nossas análises filogenéticas permitidas por esses dados destacam a monofilia do Purussaurus , como táxon irmã do clã Mourasuchus , e ambos os grupos como clã irmã dos jacarés do grupo da coroa ( Figura 1 ) em Caimaninae. A relação do grupo irmão de P. mirandai e P. brasiliensis persiste após a pontuação revisada de P. mirandai com AMU-CURS-541. A melhor resolução do clado Jacarea também indica que a pontuação questionável de Melanosuchus fisheri é provavelmente responsável pela politomia encontrada por Salas-Gismondi et al. (2015) . Os estudos de Bona et al., 2018 e Souza-Filho et al., 2019 encontraram problemas semelhantes, com o primeiro - excluindo Me. fisheri - tendo um clado Jacarea melhor resolvido, e o último - incluindo Eu. fisheri - mostrando menos resolução geral, especialmente no clado Jacarea. Análises futuras devem usar a pontuação atualizada de P. mirandai e recomenda-se uma revisão crítica das pontuações de Melanosuchus fisheri e Melanosuchus niger .
Região peitoral
A escápula e o coracóide permaneceram separados no Purussaurus , mesmo em espécimes grandes ( Figuras 2 e 4 A), enquanto que nos jacarés existentes, o fechamento da sinondrose escapulocoracóide começa relativamente cedo na ontogenia.
Esse fechamento precoce foi sugerido como uma característica peramórfica de Caimaninae ( Brochu, 1995 ), bem como um apoio ambíguo ao caráter do grupo ( Brochu, 1999 ).
A lâmina da escápula é orientada apenas ligeiramente posteriormente, e o
coracóide é orientado ventromedialmente e ligeiramente posteriormente,
dando a toda a cintura peitoral uma aparência bastante vertical ou reta
(subvertical) na vista lateral ( Figura 4A ). A sutura escapulocoracóide na espécie Purussaurus
é estreita em comparação com a das espécies existentes, porque ambos os
ossos da cintura peitoral carecem de uma expansão anterior proximal ( Figura 4A ).
A crista deltóide muito ampla e robusta da escápula do Purussaurus indica um local de origem bem desenvolvido do músculo deltoideus clavicularis ( Meers, 2003 ; = M. deltoideus scapularis inferior sensu Brochu, 1999 ). O deltoideus clavicular provavelmente reforçou a ancoragem do úmero na cintura escapular. Expansões análogas da inserção desse músculo na porção proximal do úmero no Stratiotosuchus crocodiliforme baurusuchida foram citadas como evidência para melhorar a função do membro parasagital ( Riff e Kellner, 2011 ). A crista deltóide também pode ter hospedado partes das inserções dos músculos levator scapularis e trapezius / cucullaris ( Cong, 1998 ), que deveriam ter funções relacionadas à estabilização / mobilização da cintura peitoral ou do pescoço.
Figura 4
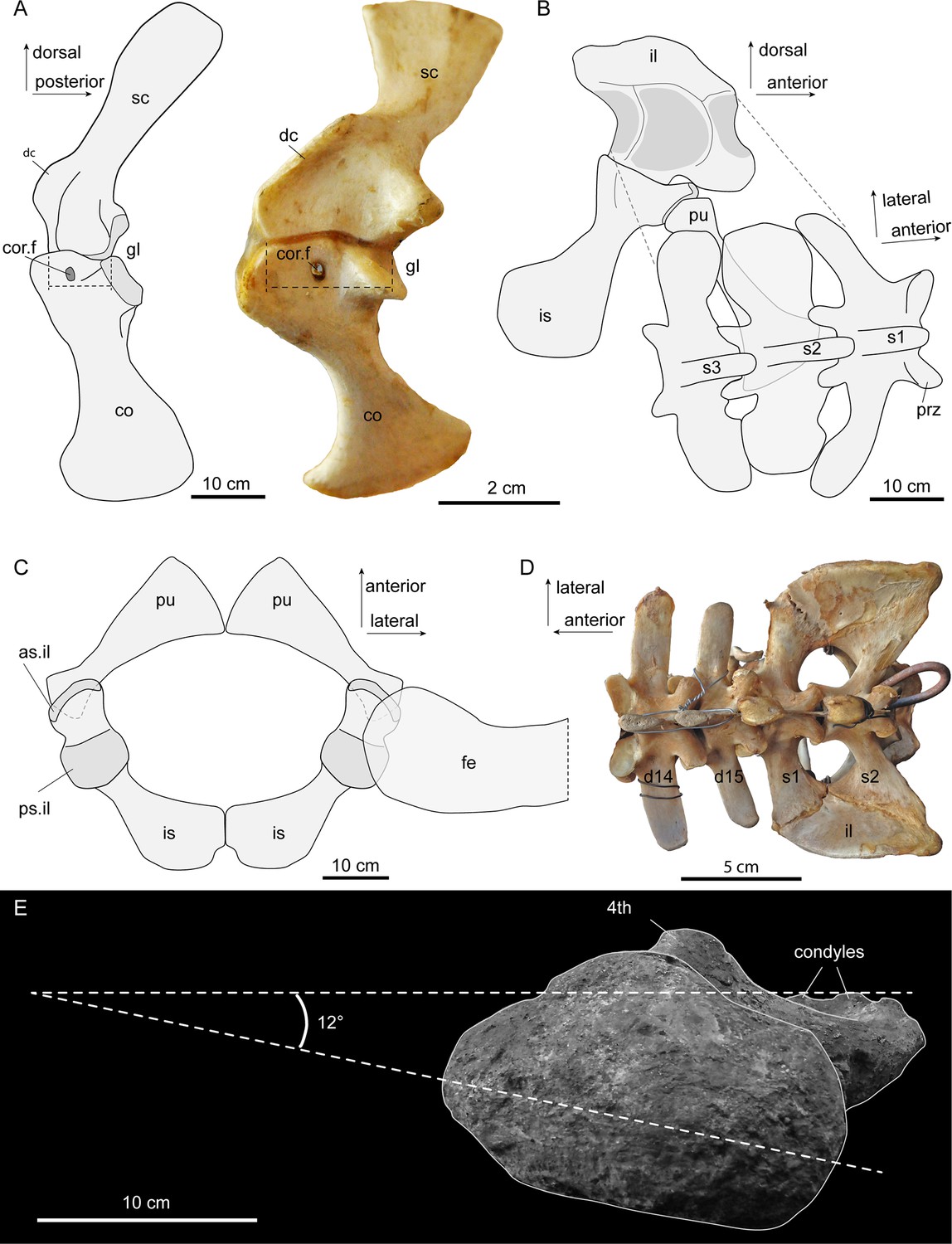
Esboços interpretativos da articulação da cintura em Purussaurus , com base nos ossos da cintura de várias amostras (AMU-CURS-541; UNEFM-CIAAP 1367; UNEFM-CIAAP-1369) em comparação com os ossos existentes selecionados.
( A ) Cintura peitoral direita em vista medial em comparação com uma cintura peitoral esquerda (espelhada) de um Crocodylus niloticus existente. Linhas pontilhadas indicam a largura da articulação óssea entre a escápula e o coracóide. ( B ) cintura pélvica esquerda em vista medial e porção vertebral sacral em vista dorsal em ângulo. ( C ) Púbis e ísquios (ossos foram espelhados para finalização) e cabeça femoral sobreposta em vista dorsal. ( D ) Transição dorsosacral da coluna vertebral em um jacaré Caiman existente. ( E ) Vista proximal do fêmur, revelando baixa torção (12 °) da cabeça femoral ao plano dos côndilos distais. Abreviações: as.il, superfície articular anterior do ílio; 4º , quarto trocanter; co, coracóide; cor.f, forame coracóide; d14-d15, 14ª e 15ª vértebra dorsal; dc, crista deltóide; fe, fêmur; gl, fossa glenóide; ílio, ílio; é ísquio; pu, púbis; prz, prezygapophysis; ps.il, superfície articular posterior para ílio; s1-s3, vértebras sacrais e costelas 1–3; sc, escápula.
Região pélvica e sacral
Os ossos da cintura pélvica do Purussaurus articulam-se de maneira semelhante à dos crocodilianos existentes ( Figura 4B, C ; ver Claessens e Vickaryous, 2012 ), com uma articulação dupla entre ílio e ísquio e uma articulação única do púbis com a margem anterior do ísquio. O ângulo da faceta articular proximal no púbis em P. mirandai
é peculiar, porque a superfície articular padrão nos táxons
crocodilianos tem um contorno sub-circular e uma única superfície
articular fracamente concavoconvexa com o ísquio (por exemplo, Claessens e Vickaryous, 2012 ) A parte lateral angular da faceta articular proximal no Purussaurus pubis é, portanto, impossível de articular com o ísquio quando a parte medial está em articulação ( Figura 4C ). As áreas de inserção medial entre os púbis distais esquerdo e direito foram provavelmente um contato pontual ( Figura 4C ); comparado aos crocodilianos existentes, o contato medial entre os ísquios foi reduzido em comprimento.
Além dos sacrais isolados, a região sacral completa com os três sacrais do UNEFM-CIAAP-1369 é conhecida por P. mirandai.
Juntamente com as evidências dos locais de articulação medial na ília,
há um forte apoio à condição 'três sacrais' neste jacaré extinto como
uma característica única dentro da coroa-Crocodylia ( Böhmer et al., 2015 ; Mook, 1921b ; Romer, 1956 ). Com base na comparação com os esqueletos crocodylianos existentes (por exemplo, Crocodylus niloticus , Caiman yacare ; Figura 4D
) e as formas das costelas sacrais e locais de fixação associados na
ília, mostramos que a última vértebra dorsal (ou seja, lombar) foi
sacralizada em um dorsossacro em P. mirandai . Isso leva a uma contagem vertebral de nove cervicais, 14 dorsais e três sacrais. A primeira caudal em P. mirandai
possui, como nos crocodilianos existentes, uma forma de centro
biconvexo, com os processos transversos sendo orientados posteriormente.
A cabeça femoral do fêmur de P. mirandai
mais bem preservado parece ter uma orientação mais direcionada
medialmente que na maioria das outras coroas-Crocodylia, a apenas ~ 12 °
em relação ao eixo mediolateral dos côndilos femorais ( Figura 4E ).
Embora a distorção óssea e o desgaste dos côndilos possam ter alterado
essa orientação, a baixa torção da cabeça femoral em comparação com os
côndilos contrasta mesmo com a orientação relativamente medial de 36 °
em Stratiotosuchus ou com os ângulos maiores encontrados em crocodilianos existentes ( Riff e Kellner, 2011 relataram um ângulo de 52 ° em uma amostra de Caiman yacare ).
Anomalias vertebrais congênitas
Em humanos, a condição patológica de uma vértebra de transição
lombossacra de malformação congênita (LSTV) é generalizada e
frequentemente estudada na literatura médica e veterinária. Propõe-se que esta condição esteja ligada a sintomas de dor lombar referidos como síndrome de Bertolotti em humanos (veja Holm et al., 2017 ; Jancuska et al., 2015 ) para visões gerais) ou predisponha a síndrome da cauda equina em cães (por exemplo, Flückiger et al., 2006 ).
Os mecanismos de desenvolvimento subjacentes a essas patologias ocorrem
precocemente durante a embriogênese, quando a expressão do gene Homeobox , que é Hox8 , Hox10 e Hox11
(os dois últimos também desempenham um papel importante no padrão de
membros), induz o padrão axial da dorsal posterior (= lombar), regiões
sacral e caudal anterior do embrião ( Casaca et al., 2014 ; Swinehart et al., 2013 ; Wellik, 2007 ; Wellik e Capecchi, 2003 ).
O Hoxc8 é expresso em toda a região dorsal (= toracolombar) do esqueleto crocodiliano, mas não na região sacral ( Böhmer et al., 2015 ; Böhmer, 2013 ; Mansfield e Abzhanov, 2010 ). Sabe-se que pequenas mudanças heterocrônicas na interação das expressões do gene Hox causam mudanças drásticas no desenvolvimento ( Gérard et al., 1997 ), como nas expressões Hoxa10 e Hoxa11 na região sacral ( Carapuço et al., 2005 ). O último estudo mostrou que alterações no tempo de expressão do Hoxa10 e Hoxa11
no mesoderma pré -omítico versus expressão nos somitos influenciavam o
desenvolvimento das costelas, a sacralização e a formação caudal na
região sacral.Como tal, a expressão de Hoxa10
no mesoderma pré -omítico levou à formação de vértebras sem costelas,
enquanto a mesma expressão nos somitos levou a vértebras com costelas;
da mesma forma, a expressão de Hoxa11 no mesoderma pré -omítico causou sacralização, enquanto nos somitos induziu a formação caudal.
Entre os crocodilianos existentes, inúmeras condições patológicas que afetam o esqueleto axial foram descritas ( Kälin, 1933 ; Rothschild et al., 2012
). Segundo informações, essas alterações incluem alterações no número e
na identidade das vértebras, como a adição vertebral na série dorsal,
deslocamento da região sacral, sacralização dos últimos dorsais e
primeiros caudais ( Baur, 1886 ; Baur, 1889 ; Reinhardt, 1873 ; Reinhardt, 1874
), mas geralmente não eram fornecidas imagens ou desenhos, e apenas
ocasionalmente os espécimes estudados eram diretamente referidos (por
exemplo, Baur, 1889),
p. 240) Uma articulação unilateral de um processo transversal esquerdo
da última vértebra dorsal hemisacralizada com o ílio esquerdo também foi
recentemente observada e figurada em um espécime patológico de Alligator mississippiensis
, enquanto discutia o aparecimento independente repetido de
dorsossacrais não patológicos entre os arquossossauros triássicos ( Griffin). et al., 2017 ). O último estudo discutiu o papel potencial dos genes Hox , especialmente os paralogais Hox11
, e as mudanças no tempo de expressão dos mesmos, para compartimentar
as séries dorsal e sacral e mudar a fronteira entre eles no fitossomo Smilosuchus da Formação Triássica Chinle Superior do nordeste do Arizona , EUA.
Em
nossa amostra de materiais comparativos, descobrimos um último dorsal
sacral hemisacralizado em uma amostra juvenil do jacaré-anão Palaeosuchus palpebrosus ( Figura 5
; amostra RVC-JRH-PP4). Este espécime exibe os ossos da cintura e
elementos estilopodiais em posição / articulação da vida e, portanto,
serve bem para comparação com os fósseis de Purussaurus . Possui uma ampla sincrondose escapular e uma cabeça femoral com torção mais forte ( Figura 5C ; com 53,5 ° semelhante ao ângulo relatado por Caiman em Riff e Kellner, 2011 ) em comparação ao plano condilar distal ( Figura 5C, D ). Conforme relatado para o espécime de A. mississippiensis (Griffin et al., 2017 ) mencionado acima, o sacro primordial 1 de P. palpebrosus ( Figura 5D, E
) mostra o desenvolvimento de um flange raso no dorso-sacro e um
ligeiro deslocamento posterior de sua base de costela sacral no lado
onde a hemisacralização ocorreu.
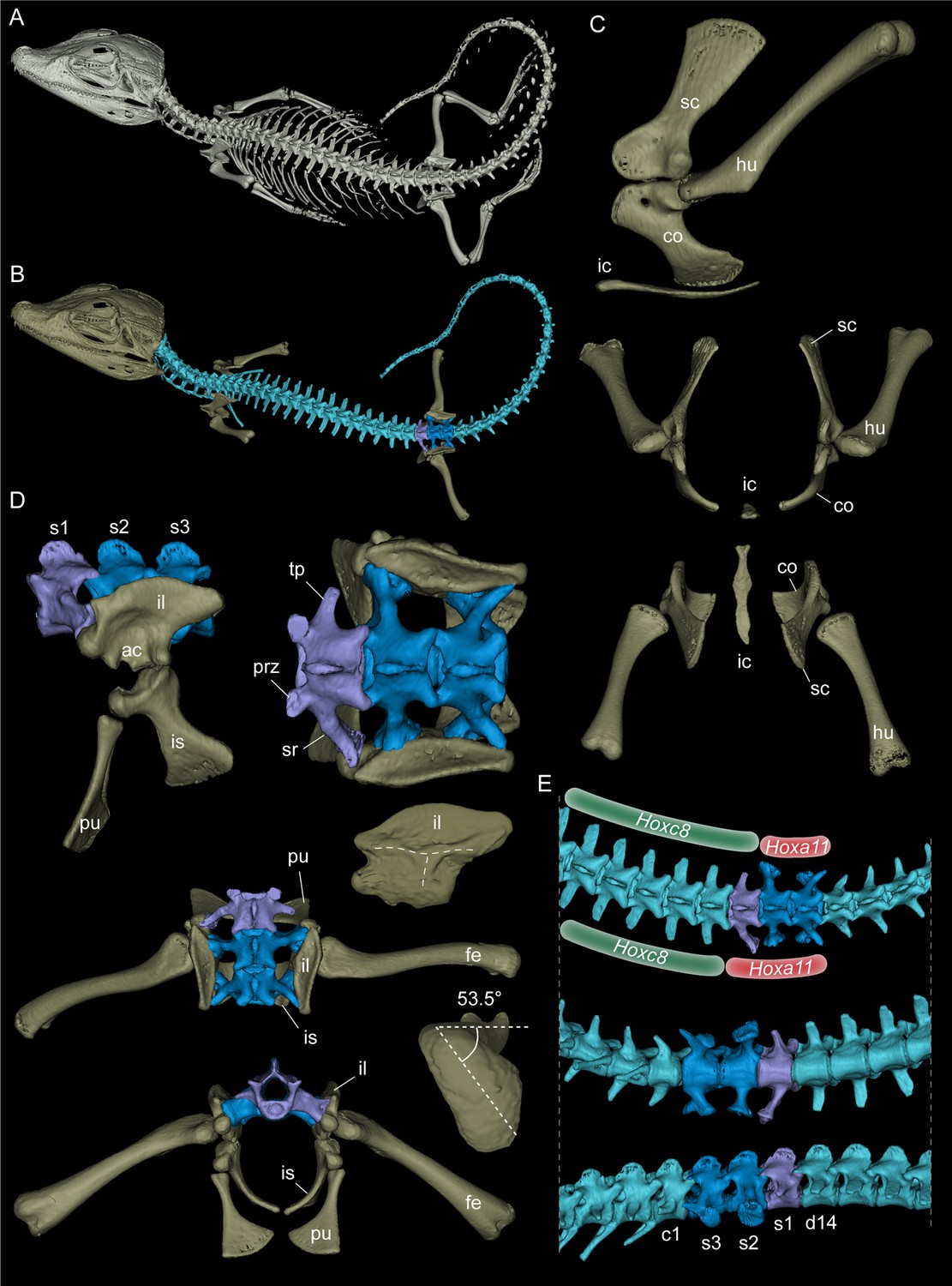
Modelos 3D renderizados de um jacaré-anão juvenil Palaeosuchus palpebrosus (RVC-JRH-PP4; sem escala).
( A ) Renderização da amostra completa. ( B
) Modelo com apenas o esqueleto apendicular axial e parcial (em vista
dorsal) realçado. As vértebras são mostradas em azul claro, exceto as da
região sacral, nas quais o dorsossacro em lavanda e os sacrais
verdadeiros ou primordiais em azul escuro. ( C ) Elementos da cintura peitoral na vista lateral esquerda, anterior e dorsal. ( D)
Região sacral em vista lateral esquerda, dorsal (e dorsal com fêmur
incluído) e vista anterior. Observe assimetria do dorso-sacro. A vista
medial do ílio direito revela as cristas 'τ' que separam as duas
superfícies articulares das costelas sacrais. O lado medial do ílio
esquerdo (não mostrado) parece semelhante, porque a costela sacralizada
do dorso-sacro articulava-se apenas levemente com a margem anterior do
ílio, não deixando cicatriz articular profunda. No fêmur direito, foi
medido um ângulo de torção de 53,5 ° da cabeça em relação ao plano dos
côndilos distais. ( E
) Concentre-se nas séries dorsal posterior, sacral e caudal anterior do
esqueleto em vista dorsal, ventral e lateral direita. A mudança
assimétrica inferir dos limites do domínio de HoxC8 e Hoxa11levando a malformação congênita vértebra de transição lombossacra nesta amostra de P. palpebrosus . Abreviações: ac, acetábulo; c1, primeira vértebra caudal; co, coracóide; D14, 14 th
vértebra dorsal; fe, fêmur; húmus; ic, interclavícula; ílio, ílio; é
ísquio; prz, prezygapophysis; pu, púbis; s1-s3: vértebras sacrais /
costelas 1–3; sc, escápula; sr, costela sacral; TP, processo
transversal.
Além disso, outro espécime patológico de P. palpebrosus
(MACV-6139; imagem cortesia de Mariano Padilla Cano) mostra uma
condição patológica 'três sacral' na qual o tamanho e a forma das
costelas sacrais e os locais de articulação nas superfícies mediais da a
ilia varia entre os lados esquerdo e direito ( Figura 5 - suplemento de figura 1 ).
Para nosso conhecimento, Purussaurus mirandai é o primeiro membro do grupo da coroa Crocodylia com três sacrais não patológicos (ver Müller et al., 2010 ). Condição semelhante foi relatada no crocodilomorfo notossuco terrestre Notosuchus
do final do Cretáceo da Argentina, que possui o segundo e o terceiro
fusões sacrais, em alguns neosuchianos atopossaurídeos com três sacrais
não fundidos ( Fiorelli e Calvo, 2008 ; Nobre e Carvalho, 2013 ; Pol, 2005 ; Tennant e Mannion, 2014
) e, em alguns Machimosaurini teleosauroides, um grupo de
crocodilomorfos marinhos de grande porte do Jurássico, nos quais o
primeiro caudal foi sacralizado (por exemplo, Johnson et al., 2018; Jouve et al., 2016 ). Todos esses grupos, no entanto, diferem fortemente do plano corporal conservado dos crocodilianos existentes.
Interpretações morfofuncionais
Molnar et al. (2014) identificaram a articulação lombossacra (entre dorsal 15 e sacral 1) da coluna vertebral de Crocodylus niloticus
como tendo a maior rigidez da articulação intervertebral, que foi
sugerida como benéfica no suporte de uma cauda grande e no combate às
forças dos membros posteriores ( Willey et al. 2004 ). Baseado em um estudo de cinco espécimes pequenos (entre 40 e 50 cm de comprimento), de Alligator mississippiensis , Willey et al. (2004)
descobriram que o centro de massa estava situado em cerca de 70% ao
longo do comprimento do tronco e a massa da cauda em até 28% da massa
corporal total. Além disso, Aureliano et al. (2015) estimaram os maiores espécimes de Purussaurus brasiliensis ter atingido mais de 12 m de comprimento e mais de oito toneladas métricas de massa.
Dado
o comprimento de 145 cm da mandíbula inferior preservada e o tamanho
das vértebras com comprimento de até 11,8 cm, estima-se que o
AMU-CURS-541 tenha variado entre oito e dez metros de comprimento total.
Infelizmente, o fêmur
associado ao AMU-CURS-541 foi apenas parcialmente preservado,
dificultando as estimativas de tamanho com base nesse elemento
esquelético ( Farlow et al., 2005 ). Comparações com outras amostras de Purussaurusda
Formação Urumaco indicam, no entanto, que o AMU-CURS-541 não é o maior
indivíduo recuperado até agora. No AMU-CURS-20, a maior vértebra
preservada (13,6 cm de comprimento do centro) é cerca de 15% maior em
comparação com as vértebras do AMU-CURS-541. Além disso, as dimensões do
fêmur direito associado e razoavelmente completo do AMU-CURS-20 (54 cm
de comprimento; em comparação com 51 cm do fêmur do holótipo
UNEFM-CIAAP-1369 de P. mirandai ) estão entre os maiores fêmures crocodylianos ( Salas-Gismondi et al., 2007 relataram um fêmur de 54,5 cm de comprimento do Purussaurus
do Peru). Com 210 mm, o fêmur do AMU-CURS-20 possui uma circunferência
mínima similar do eixo do fragmento femoral MNN G102–2 do imperador Sarcosuchus , usado porFarlow et al. (2005) para estimar o comprimento total desse espécime entre 7,2 e 9,1 m.
Utilizamos o conjunto de dados e o código do software R de O'Brien et al. (2019) com entrada de P. mirandai na filogenia conforme a Figura 1
e com um comprimento de ramo de 0,01 (assumindo 25% de massa corrigida
conforme seus métodos) para estimar a massa corporal total e o
comprimento total da amostra AMU-CURS-541 ( largura da cabeça ~ 0,88 m).
Com base nessa abordagem mais conservadora, nossa análise estimou 1686 a
2637 kg e 7,11 a 8,01 m para massa corporal total e comprimento,
respectivamente (das faixas interquartis inferiores para as superiores
da análise bayesiana). Isso se compara favoravelmente com outros grandes
Crocodyliformes, como Sarcosuchus imperator (conforme O'Brien et al., 2019cálculos
de 'longirostrina'), estimados em 1976-2981 kg e 8,50-9,47 m (Tabela
5). Comparado com a aparência mais maciça e robusta de P. brasiliensis , o espécime mais ' gracioso ' de P. mirandai AMU-CURS-541 poderia ter a cauda sozinha pesando até 472-738 kg (com base em Willey et al., 2004 ; valores que excedem a maioria das massas médias de crocodilianos existentes ( Grigg e Kirschner, 2015 ); o excepcionalmente grande 'Lolong', um antigo Crocodylus porosus com 6,17 m de comprimento, foi relatado em 1075 kg: Britton et al., 2012 ).
Independentemente
da estimativa de massa utilizada, especulamos que a condição dos três
sacrais, juntamente com a robustez dos ossos dos membros posteriores
encontrados em P. mirandai, estivesse relacionada ao tamanho gigante e à massa corporal desses animais ( Aureliano et al., 2015 ; Salas-Gismondi et al., 2007
). Um sacro mais longo, com três em vez de apenas dois sacrais
articulados com o ílio, expande a conexão entre o esqueleto axial e o
esqueleto apendicular posterior, aumentando potencialmente a
estabilidade dessa região na coluna vertebral e permitindo uma melhor
transmissão de forças através da bacia pélvica e caudal regiões.
Dados comparativos de outros crocodilianos gigantes, no entanto, são escassos ou permanecem pouco descritos (por exemplo, Deinosuchus : Colbert e Bird, 1954 ; Holland, 1909 ; Rivera-Sylva et al., 2011 ; Schwimmer, 2002 ; Euthecodon : Storrs, 2003 ; Gryposuchus : Langston, 1965 ; Riff e Aguilera, 2008 ; Laganosuchus : Sereno e Larsson, 2009 ; Mourasuchus : Langston, 1965 ; Sarcosuchus : Dridi, 2018 ; Sereno et al., 2001)
Assim, a influência do tamanho e da massa corporal na região sacral dos
crocodilianos gigantes permanece em grande parte inexplorada. Desses
táxons, pelo menos um ílio bem preservado de Mourasuchus atopus mostra um padrão 'π' de cristas em sua superfície medial (UCMP 38012, Figura 3 - suplemento de figura 2
). Isso pode indicar a presença de três sacrais nesse caimanino gigante
adicional, se confirmado por outras amostras. A condição 'três sacral'
aqui descrita é puramente de origem funcional, pois Mourasuchus e Purussaurus são gigantes (embora provavelmente possuam
massas corporais distintamente diferentes), ou também pode haver um
sinal filogenético envolvido, pois ambos os táxons são gêneros irmãos
dentro de Caimaninae .
Outros
estudos de Crocodylomorpha / Archosauria inferiram que ilia expandida e
sacralização se correlacionam com uma postura mais ereta dos membros e /
ou melhor suporte contra a gravidade (por exemplo, Riff e Kellner, 2011
). A evidência acima da região iliossacral corrobora outras
especializações para orientação mais vertical dos membros ou
simplesmente suporte de peso em P. mirandai , incluindo a cinta peitoral mais orientada verticalmente e a baixa torção da cabeça femoral em relação aos côndilos.
Conclusões
As regiões peitoral e pélvica de Purussaurus mirandai
mostram uma morfologia peculiar em comparação com os táxons existentes,
incluindo um contato escapulocoracóide estreito, crista deltóide larga e
proeminente na escápula, que serve como origem muscular, cintura
peitoral mais orientada verticalmente e 'três sacrais' autapomórficos
condição na pelve, aqui interpretada como ligada ao tamanho gigante do
corpo. Infere-se que os mecanismos subjacentes do desenvolvimento,
especialmente um tempo anterior de expressão de HOX11 e supressão parcial de Hox10 ( Wellik e Capecchi, 2003 ), resultando em domínios deslocado de HoxC8 e HOX11 , levou à formação do dorsosacral em P. mirandai. A descoberta e o exame de espécimes adicionais de Purussaurus
com preservação excepcional para documentar a variação de gênero e de
outros crocodilianos gigantes levariam a uma melhor compreensão das
peculiaridades anatômicas desses táxons. Tais estudos devem inspecionar o
lado medial do ílio para fornecer informações indiretas sobre a
contagem sacral.
materiais e métodos
Os espécimes de Purussaurus
utilizados neste estudo derivam de diferentes localidades no final da
Formação Mioceno Urumaco, Estado de Falcón, Venezuela. O AMU-CURS-541,
um espécime coletado em 2013, consiste em material craniano, maxilar
inferior e pós-craniano associados. Nosso estudo também incluiu:
UNEFM-CIAAP-1367 (uma amostra amplamente não descrita, incluindo
elementos da cintura peitoral e pélvica), UNEFM-CIAAP-1369 (material
referente ao holótipo e não holótipo, este último incluindo um sacro
articulado completo), MCNC-URU- 111–72V e MCNC-URU-158–72V (assembléias
mistas de Purussaurus
), AMU-CURS-020 (dois fêmures associados a duas vértebras) e
AMU-CURS-528 (material pós-craniano associado a restos desarticulados de
crânio).
Os materiais comparativos existentes incluíam amostras de Crocodylus e Gavialis em PIMUZ, Caimaninae das coleções MACUT, bem como material armazenado e digitalizado pelo RVC (veja abreviações abaixo).
Análise filogenética
Solicite um protocolo detalhado
Os
numerosos elementos pós-cranianos do novo espécime AMU-CURS-541 foram
usados para marcar novamente alguns caracteres até então desconhecidos
para a espécie Purussaurus mirandai . Todos os personagens são retirados de Salas-Gismondi et al. (2015) , uma matriz amplamente baseada e modificada de Brochu (2011) e Jouve et al. (2008) .
Três análises de parcimônia máxima foram realizadas usando o TNT v. 1.5 ( Goloboff e Catalano, 2016 ). Na primeira análise, o AMU-CURS-541 foi classificado como um táxon terminal separado, além das três espécies de Purussaurus: P. mirandai , P. neivensis e P. brasiliensis . Para a segunda análise, as novas pontuações baseadas no AMU-CURS-541 foram adicionadas às pontuações anteriores de P. mirandai
. Os índices de consistência e os valores de suporte da Bremer foram
calculados no TNT v. 1.5, usando os scripts 'stats.run' e 'Bremer.run'
(baixados em 11.11.2018 em http://phylo.wikidot.com/tntwiki ; veja também http: //gensoft.pasteur.fr/docs/TNT/1.5/)
e o último foi verificado manualmente pelo colapso das topologias de
árvores através da inclusão de sucessivas árvores subótimas. Nas duas
análises filogenéticas, o táxon extinto Melanosuchus fisheri foi removido devido a inconsistências recentemente reveladas entre o holótipo e o material referido ( Bona et al., 2017 ; Foth et al., 2018 ). Na terceira análise, foi utilizada a pontuação atualizada de P. mirandai , exceto Me. fisheri
foi deixado para ver como a adição ou remoção desse táxon influenciou
os resultados. Em todas as análises, a pesquisa heurística (pesquisa
tradicional; espaço 60000 árvores na memória, semente aleatória = 1,
modo de reconexão por bisseção de árvore (TBR) ativado)) foi executada
com Bernissartia fagesiicomo grupo externo, 1000 repetições adicionais aleatórias em sequência e 100 árvores salvas por replicação. Os caracteres eram igualmente ponderados, desordenados e definidos como não aditivos.
Pontuações para análises filogenéticas
Solicite um protocolo detalhado
Escores de 201 caracteres utilizados para P. mirandai atualizados pelos escores de AMU-CURS-541 na matriz de Salas-Gismondi et al. (2015) :
1 110 ?? 00001 0? 0? 0 11110 01 ??? 111 0 11? 01? ? 1100
00? 2? ? 1? 11? 1201 100? 1 1001? 0? 112? 0021? 10000 11100
10 00 ??? 0000 00011 11? 1? 11021 0? 111 110? 1 10200 10112
000 ?? 10104 ????? ?????0011? 2,00? 10 ?? 0 01000 00 ?? 1 00? 10 0
Escores de 201 caracteres usados para AMU-CURS-541 como táxon terminal na matriz de Salas-Gismondi et al. (2015) :
1 110 ?? 00001 0? 0? 0 11110 0 ???? 111 11? 01? ? 1100
00? 2? 1 ?? 2 ?? 00 001 1001? 0? ????? ????? ????? ?????
????? ????? ????? ????? ????? ????? ????? ????? ????? ?????
????? ????? ????? ????? ????? ????? ????? ????? ????? 1? ?
Abreviações institucionais. AMU-CURS,
Solicite um protocolo detalhado
Coleção
Paleontológica da Alcaldía Bolivariana de Urumaco, Estado Falcón,
Venezuela; MACUT, Coleções do Museu de Anatomia Comparada da
Universidade de Turim, hospedado pelo Museu Regional de Ciências
Naturais de Turim, Itália; MACV, Museu de Anatomia Comparada dos
Vertebrados, Universidade Complutense de Madri, Espanha. MCNC, Museu de
Ciências Naturais de Caracas, Venezuela; MNN, Museu Nacional do Níger,
Niamey, Níger; PIMUZ, Instituto e Museu Paleontológico, Universidade de
Zurique, Suíça; QM, Queensland Museum, Brisbane, Austrália; RVC, Royal
Veterinary College, Londres, Reino Unido; UNEFM-CIAAP, Universidade
Nacional Experimental Francisco de Miranda / Centro de Investigações
Antropológicas, Arqueológicas e Paleontológicas, Coro, Venezuela.
Referências
- 1 Um novo Purussaurus gigante (Crocodyliformes, Alligatoridae) da Formação Mioceno Superior Urumaco, VenezuelaJornal de Paleontologia Sistemática 4 : 221–232.https://doi.org/10.1017/S147720190600188X
- 2
- 3
- 4 Revisão meiner Mitteilungen im Zool. Anz., Mit NachträgenZoologischer Anzeiger 12 : 238–243.
- 5 Fakultät Für GeowissenschaftenCorrelação entre código Hox vertebral e morfologia vertebral em arquossauros: implicações para a evolução vertebral em dinossauros sauropodomorph, Fakultät Für Geowissenschaften, Munique, Ludwig-Maximilians-Universität München.
- 6 Novas idéias sobre o código Hox vertebral dos arquossaurosEvolution & Development 17 : 258–269.https://doi.org/10.1111/ede.12136
- 7
- 8 Um novo paleoceno crocodyliano do sul da Argentina lança luz sobre o início da história dos caimaninosAnais da Royal Society B: Biological Sciences 285 : 20180843.https://doi.org/10.1098/rspb.2018.0843
- 9 Aqui está um dragão: tamanho excepcional em um crocodilo de água salgada ( Crocodylus porosus ) das FilipinasHerpetological Review 43 : 541-546.
- 10 Heterocronia no escapulocoracóide crocodilianoJournal of Herpetology 29 : 464–468.https://doi.org/10.2307/1565002
- 11 Filogenética, taxonomia e biogeografia histórica de AlligatoroideaLivro de memórias 6 : 9–100 da Sociedade de Paleontologia de Vertebrados .https://doi.org/10.2307/3889340
- 12 Relações filogenéticas de Necrosuchus ionensis Simpson, 1937 e o início da história dos caimaninosRevista Zoológica da Sociedade Linnean 163 : S228 – S256.https://doi.org/10.1111/j.1096-3642.2011.00716.x
- 13 Padrões macroevolutivos na radiação evolutiva de arquossauros (Tetrapoda: Diapsida)Transações com Ciências da Terra e do Meio Ambiente da Sociedade Real de Edimburgo 101 : 367–382.https://doi.org/10.1017/S1755691011020056
- 14 Os genes Hox especificam tipos vertebrais no mesoderma pré -omíticoGenes & Development 19 : 2116-2121.https://doi.org/10.1101/gad.338705
- 15 Controlando a expressão e a atividade do gene Hox para construir o esqueleto axial dos vertebradosDinâmica do Desenvolvimento 243 : 24–36.https://doi.org/10.1002/dvdy.24007
- 16
- 17 Um crocodilo gigantesco do alto leito cretáceo do TexasNovitates do Museu Americano 1688 : 1–22.
- 18 A anatomia grossa de Alligator sinensis Fauvel (Di 1 Ban)Pequim: Ke xue chu ban ela.
- 19 Novos fósseis do gênero gigante pholidosaurid Sarcosuchus do início do Cretáceo da TunísiaJournal of African Earth Sciences 147 : 268–280.https://doi.org/10.1016/j.jafrearsci.2018.06.023
- 20 Dimensões femorais e tamanho corporal de Alligator mississippiensis: estimando o tamanho de mesoucrocodilianos extintosJournal of Vertebrate Paleontology 25 : 354–369.https://doi.org/10.1671/0272-4634(2005)025[0354:FDABSOorgeous2.0.CO;2
- 21 Funções de desenvolvimento de genes Hox de mamíferosReprodução Humana Molecular 3 : 115–131.https://doi.org/10.1093/molehr/3.2.115
- 22 Novos restos de Notosuchus terrestris Woodward, 1896 (Crocodyliformes: mesoeucrocodylia) do final do Cretáceo de Néuquen, Patagônia, ArgentinaArquivos do Museu Nacional, Rio de Janeiro 66 : 83–124.
- 23 Uma vértebra de transição lombossacra no cão predispõe à síndrome da cauda eqüinaVeterinary Radiology & Ultrasound 47 : 39–44.https://doi.org/10.1111/j.1740-8261.2005.00103.x
- 24
- 25
- 26
- 27
- 28.
- 29
- 30 Vértebras e costelas de répteis modernosIn: C Gans, AdA Bellairs, T. S Parsons, editores. Biologia dos Reptilia . Londres: Morfologia A. Academic Press. 201-310.
- 31 Deinosuchus hatcheri , um novo gênero e espécie de crocodilo dos leitos do rio Judith, em MontanaAnais do Carnegie Museum 6 : 281–294.
- 32.
- 33 Uma revisão das vértebras transitórias lombossacrais sintomáticas: Síndrome de BertolottiRevista Internacional de Cirurgia da Coluna 9 : 42.https://doi.org/10.14444/2042
- 34 Re-descrição do 'Steneosaurus' obtusidens Andrews, 1909, um incomum crocodilomorfo teleossóide macrófago do Jurássico Médio da InglaterraZoological Journal of the Linnean Society 182 : 385–418.https://doi.org/10.1093/zoolinnean/zlx035
- 35
- 36.
- 37. Über Skeletanomalien bei CrocodilidenZoomorphology 32 : 327–347.https://doi.org/10.1007/BF00403078
- 38. Crocodilos fósseis da Colômbia e a história cenozóica dos crocodilos na América do SulPublicações da Universidade da Califórnia em Ciências Geológicas 52 : 1–169.
- 39. Expressão de Hox no jacaré americano e evolução do padrão axial archosaurianoJornal de Zoologia Experimental Parte B: Evolução Molecular e de Desenvolvimento 314B : 629–644.https://doi.org/10.1002/jez.b.21364
- 40.
- 41 Musculação crocodylian do membro anterior e sua relevância para ArchosauriaO registro anatômico 274A : 891–916.https://doi.org/10.1002/ar.a.10097
- 42. Teste experimental e morfométrico da relação entre morfologia vertebral e rigidez articular em crocodilos do Nilo ( Crocodylus niloticus )Journal of Experimental Biology 217 : 758–768.https://doi.org/10.1242/jeb.089904
- 43 Brachygnathosuchus braziliensis , um novo crocodilo fóssil do BrasilBoletim 44 : 43–49 do Museu Americano de História Natural .
- 44 Notas sobre o esqueleto pós-craniano na CrocodiliaBoletim 44 : 67–100 do Museu Americano de História Natural .
- 45 Um novo crocodilo fóssil da ColômbiaAnais do Museu Nacional dos Estados Unidos 91 : 55–58.https://doi.org/10.5479/si.00963801.91-3122.55
- 46.
- 47
- 48. Abordagens Cladísticas da Paleobiologia Aplicadas à Filogenia dos AligatorídeosNew Haven: Universidade de Yale.
- 49. Alometria da largura da cabeça de Crocodylian e predição filogenética do tamanho do corpo em crocodyliforms extintosBiologia Organizacional Integrativa 1 : obz006.https://doi.org/10.1093/iob/obz006
- 50. Restos pós-cranianos de Notosuchus terrestris Woodward (Archosauria: Crocodyliformes) do alto Cretáceo da Patagônia, ArgentinaAmeghiniana 42 : 21.
- 51 Anomalier I Krydevirvel hos KrokodilerneVidenskabelige Meddelelser Frå Den Naturhistoriske Forening I Kjöbenhavn 45 : 221–228.
- 52 Nas anomalias das vértebras sacras, que são os crocodilosJournal de Zoologie 3 : 308–312.
- 53 Amazônia, paisagem e evolução de espéciesFauna neotropical de crocodilo e tartaruga no norte da América do Sul, Amazônia, paisagem e evolução de espécies, Londres, Blackwell Publishing.
- 54
- 55 Crocodyliformes baurusuchídeos como imitadores de terópodes: pistas do crânio e morfologia apendicular de Stratiotosuchus maxhechti (Cretáceo Superior do Brasil)Revista Zoológica da Sociedade Linnean 163 : S37 – S56.https://doi.org/10.1111/j.1096-3642.2011.00713.x
- 56. A Deinosuchus riograndensis (Eusuchia: Alligatoroidea) de Coahuila, norte do MéxicoRevista Mexicana de Ciências Geológicas 28 : 267–274.
- 57
- 58.
- 59 Crocodilos do mioceno médio do Arco Fitzcarrald, Peru da Amazônia, Instituto Geológico e Minero de España, MadriArtigo apresentado no 4º Encontro Europeu de Paleontologia e Estratigrafia da América Latina Cuadernos Del Museo Geominero.
- 60 Comunidade crocodiliana hiperdiversa do Mioceno revela dinâmica trófica peculiar em mega-zonas úmidas proto-amazônicasProceedings of the Royal Society B: Biological Sciences 282 : 20142490.https://doi.org/10.1098/rspb.2014.2490
- 61
- 62 Vertebrados neogênicos de Urumaco, Estado de Falcón, Venezuela: Diversidade e importânciaJornal de Paleontologia Sistemática 4 : 213-220.https://doi.org/10.1017/S1477201906001829
- 63.
- 64 A falecida fauna caimanina do mioceno (Crocodylia: Alligatoroidea) da Formação Urumaco, VenezuelaPalaeontologia Electronica 19 : 1–57.https://doi.org/10.26879/657
- 65 Rei dos Crocodylians: A Paleobiologia de DeinosuchusBloomington: Indiana University Press.
- 66.
- 67
- 68 Nova espécie caimanina (Crocodylia, alligatoroidea) da Formação Solimões do Brasil e a filogenia de CaimaninaeJournal of Vertebrate Paleontology 38 : e1528450.https://doi.org/10.1080/02724634.2018.1528450
- 69
- 70 Mioceno tardio - fauna crocodiliana do início do Plioceno de Lothagam, sudoeste da Bacia de Turkana, QuêniaIn: MG Leakey, JM Harris, editores. Lothagam: O alvorecer da humanidade na África Oriental . Nova York: Columbia University Press. 137-159.
- 71 Disparidade morfológica e biomecânica de arquossauros de linha de crocodilo após a extinção Triássica finalAnais da Royal Society B: Biological Sciences 280 : 20131940.https://doi.org/10.1098/rspb.2013.1940
- 72
- 73
- 74 Padrões Hox do esqueleto axial dos vertebradosDynamics Developmental 236 : 2454–2463.https://doi.org/10.1002/dvdy.21286
- 75
- 76 O conto da cauda: função dos membros e mecânica locomotora em Alligator mississippiensisJournal of Experimental Biology 207 : 553–563.https://doi.org/10.1242/jeb.00774
Carta de decisão
- Diethard TautzEditor sénior; Instituto Max-Planck de Biologia Evolutiva, Alemanha
- John A LongEditor de Revisão; Universidade de Flinders, Austrália
- Michelle StockerRevisor
- Chris BrochuRevisor; Universidade de Iowa, Estados Unidos
No interesse da transparência, o eLife publica as solicitações de
revisão mais substanciais e as respostas dos autores que as acompanham.
Resumo de aceitação:
Este
manuscrito destaca o crescente reconhecimento de que a variação na
morfologia fóssil é comum e muitos táxons quebram as restrições
propostas anteriormente à morfologia que resultam em pensamento
tipológico. A anatomia
sacral deste crocodiliano gigante é descrita como compreendendo três
sacrais, quebrando a restrição existente de que os crocodilianos têm a
condição ancestral de apenas dois sacrais e adicionando à lista de
táxons não dinossauros extintos que têm evolução pélvica mais complexa
do que o reconhecido anteriormente. Os autores discutem isso no contexto dos efeitos conhecidos das mudanças na expressão do gene Hox , tornando este um possível exemplo de polimorfismo morfológico mediado pelo gene Hox que permitiu a adaptação ao tamanho corporal excepcional.
Carta de decisão após revisão por pares:
Obrigado
por enviar o seu artigo "Gigante jacaré extinto quebra a restrição no
esqueleto axial dos crocodilianos existentes" à consideração da eLife . Seu
artigo foi revisado por dois revisores e a avaliação foi supervisionada
por um Editor de Revisão e por Diethard Tautz como Editor Sênior. As
seguintes pessoas envolvidas na análise de sua submissão concordaram em
revelar sua identidade: Michelle Stocker (Avaliadora nº 1); Chris Brochu (revisor # 2).
Os revisores discutiram as revisões entre si e o Editor de Revisão
redigiu esta decisão para ajudá-lo a preparar um envio revisado.
Resumo:
O manuscrito recebeu críticas favoráveis que solicitam apenas pequenas revisões, conforme listado abaixo. Ambos os revisores concordam que é importante compreender melhor a evolução crocodiliana mesozóica.
Revisor # 1:
Este
manuscrito destaca o crescente reconhecimento de que a variação na
morfologia fóssil é comum e muitos táxons quebram as restrições
propostas anteriormente à morfologia que resultam em pensamento
tipológico. A anatomia sacral do Purussaurus mirandai
é descrita como compreendendo três sacrais, quebrando a restrição
existente de que os crocodilianos têm a condição ancestral de apenas
dois sacrais e adicionando à lista de táxons não dinossauros extintos
que têm evolução pélvica mais complexa do que o reconhecido
anteriormente. Os autores
fazem um bom trabalho ao descobrir a anatomia relevante (a Figura 4 é
especialmente interessante!) E discutir aspectos funcionais e de
desenvolvimento dessa morfologia. Não vejo razão para que este manuscrito não possa ser publicado com uma revisão rápida. Bom trabalho!
Revisor # 2:
Este é um manuscrito muito interessante. É certo que fiquei cético quando vi o título pela primeira vez. Eu
vi o estranho espécime moderno de crocodylia com um sacro adicional -
um dorsal sacralizado ou um caudal sacralizado - envolvido no quadril. Os autores fizeram um bom trabalho ao argumentar de forma convincente que a condição dos três sacros é consistente em P. mirandai e pode ter uma distribuição mais ampla entre os jacarandás de corpo grande intimamente relacionados.
O texto é geralmente claro. Existem alguns erros gramaticais, mas os argumentos dos autores nunca foram claros.
Revisões essenciais:
- Figura 2: Como a morfologia do ílio é essencial para seus argumentos,
seria mais eficaz separar H, I e J como uma figura separada (faça o 2 e
coloque o material da cintura peitoral e a reconstrução em uma nova
Figura 3) para enfatizar sua importância e aumentar as partes da figura.
- Subseção “Cinturão pélvica”, segundo parágrafo: eu imagino esse
Morfotipo 4 para que o leitor possa comparar e contrastar por si
próprio.
- Subseção “Região sacral”: você pode fornecer alguma informação sobre a morfologia das faces do centro além do tamanho? Existem flanges nas bordas laterais das faces do centro?
- Posso tirar algumas fotos em algum lugar de um espécime de Alligator mississippiensis
com dorsal ou caudal sacralizado. O mesmo pode ser verdade para outras
espécies para as quais vi grandes amostras (por exemplo, Crocodylus acutus, Crocodylus niloticus / suchus
). Eles ainda podem estar no formato de slide de 35 mm, mas vou olhar.
Ficaria feliz em compartilhar essas imagens com os autores, se eles
estiverem interessados.
Também vou verificar se tenho alguma foto utilizável do Euthecodon ilium. Os pós-cranianos de Euthecodon
são poucos e distantes entre as coleções da África Oriental,
principalmente por causa da maneira como os crocodilianos são
historicamente coletados na região. O material pós-craniano foi coletado
apenas recentemente e geralmente é coletado na superfície - o que
significa que muitas vezes não pode realmente ser associado a nenhuma
espécie em particular. Dito isto - há um esqueleto articulado de Euthecodon
no chão em Koobi Fora. Recebi algumas fotos do espécime há alguns anos e
vejo um ílio. Voltarei com os autores se eles estiverem interessados
em algo disso.
- Eu recomendaria dividir a Figura 2 em várias figuras. Alguns
dos detalhes destacados no foco do trabalho - as facetas das costelas
sacrais na ileia em particular - seriam muito mais fáceis de ver se eram
mostrados em tamanho maior. Pode ser bom, por exemplo, mostrar os elementos peitorais, a ilia e os sacrais como figuras separadas.
(À parte as figuras, a Figura 1 na versão do manuscrito que revi foi
truncada à direita. Suspeito que esse seja um artefato da conversão do
manuscrito para o formato PDF, mas talvez seja bom checar novamente.)
- Algumas análises adicionais dos caimaninos fósseis foram publicadas, por exemplo, Bona et al. em Protocaiman (que os autores citaram) e Cossette e Brochu em Bottosaurus . Duvido
fortemente que a inclusão de tais táxons tenha impacto nos resultados
gerais deste trabalho, mas pode valer a pena considerar a adição desses
táxons - principalmente porque o ílio de Bottosaurus harlani é conhecido.
(Não compro a possível sinonímia de M. fisheri e M. niger
. Acho que são táxons separados e devem ser tratados como tal. Mas,
novamente, isso não mudaria nada do que os autores estão dizendo.)
https://doi.org/10.7554/eLife.49972.sa1 Resposta do autor
Revisões essenciais:- Figura 2: Como a morfologia do ílio é essencial para seus argumentos, seria mais eficaz separar H, I e J como uma figura separada (faça o 2 e coloque o material da cintura peitoral e a reconstrução em uma nova Figura 3) para enfatizar sua importância e aumentar as partes da figura.
De acordo com a sugestão do revisor, dividimos a Figura 2 original em
uma nova Figura 2 (cintura peitoral) e Figura 3 (cintura pélvica e
sacro) com imagens ampliadas.
- Subseção “Cinturão pélvica”, segundo parágrafo: eu imagino esse Morfotipo 4 para que o leitor possa comparar e contrastar por si próprio.
Uma figura e desenho interpretativo dos morfotipos 3 e 4 foram incorporados na Figura 3 - suplemento à figura 2.
- Subseção “Região sacral”: você pode fornecer alguma informação sobre a morfologia das faces do centro além do tamanho? Existem flanges nas bordas laterais das faces do centro?
Adicionamos duas frases ao texto em “região sacral” para descrever as
facetas de articulação do centro sacral, especialmente do sacral
primordial 2 bem preservado do AMU-CURS-541.
- Posso tirar algumas fotos em algum lugar de um espécime de Alligator mississippiensis com dorsal ou caudal sacralizado. O mesmo pode ser verdade para outras espécies para as quais vi grandes amostras (por exemplo, Crocodylus acutus, Crocodylus niloticus / suchus). Eles ainda podem estar no formato de slide de 35 mm, mas vou olhar. Ficaria feliz em compartilhar essas imagens com os autores, se eles estiverem interessados.
Adicionamos outro espécime de Palaeosuchus palpebrosus
à nova Figura 5 - suplemento à figura 1 que mostra uma condição
3-sacral patológica. Agradecemos ainda ao revisor pela oferta de
fornecer imagens adicionais, mas o objetivo do nosso artigo não é
fornecer uma visão abrangente das aberrações ou patologias do
desenvolvimento na região sacral dos crocodilianos existentes. Por isso,
gostamos de não adicionar mais imagens aqui.
Também vou verificar se tenho alguma foto utilizável do Euthecodon ilium. Os pós-cranianos de Euthecodon são poucos e distantes entre as coleções da África Oriental, principalmente por causa da maneira como os crocodilianos são historicamente coletados na região. O material pós-craniano foi coletado apenas recentemente e geralmente é coletado na superfície - o que significa que muitas vezes não pode realmente ser associado a nenhuma espécie em particular. Dito isto - há um esqueleto articulado de Euthecodon no chão em Koobi Fora. Recebi algumas fotos do espécime há alguns anos e vejo um ílio. Voltarei com os autores se eles estiverem interessados em algo disso.
Agradecemos ao revisor por todas as informações sobre o Euthecodon . Como apontado, a maior parte do material de Euthecodon é isolada e a amostra em campo ainda não foi descrita e calculada. Portanto, mantemos o texto como está no final da Discussão.
- Eu recomendaria dividir a Figura 2 em várias figuras. Alguns dos detalhes destacados no foco do trabalho - as facetas das costelas sacrais na ileia em particular - seriam muito mais fáceis de ver se eram mostrados em tamanho maior. Pode ser bom, por exemplo, mostrar os elementos peitorais, a ilia e os sacrais como figuras separadas.(À parte as figuras, a Figura 1 na versão do manuscrito que revi foi truncada à direita. Suspeito que esse seja um artefato da conversão do manuscrito para o formato PDF, mas talvez seja bom checar novamente.)
De acordo com a sugestão do revisor, dividimos a Figura 2 original em uma nova Figura 2 e 3 (veja o comentário acima). Não queremos dividir ainda mais os números para cumprir também as diretrizes da revista. O truncamento da imagem ocorreu devido a um erro de dimensionamento que foi corrigido na nova versão da Figura 1.
- Algumas análises adicionais dos caimaninos fósseis foram publicadas, por exemplo, Bona et al. em Protocaiman (que os autores citaram) e Cossette e Brochu em Bottosaurus. Duvido fortemente que a inclusão de tais táxons tenha impacto nos resultados gerais deste trabalho, mas pode valer a pena considerar a adição desses táxons - principalmente porque o ílio de Bottosaurus harlani é conhecido.
Estamos cientes do artigo de Cossette e Brochu sobre Bottosaurus , bem como Souza-Filho et al., 2019 sobre Acresuchus pachytemporalis. Este
último não possui a região sacral ou um ílio preservado, nem a
morfologia e a posição filogenética do espécime podem ser verificadas
por nós até o momento. Portanto, evitamos adicioná-lo ao manuscrito.
(Não compro a possível sinonímia de M. fisheri e M. niger. Acho que são táxons separados e devem ser tratados como tal. Mas, novamente, isso não mudaria nada do que os autores estão dizendo.)
É
claro que o revisor tem o direito de expressar sua opinião sobre isso -
evitamos modificar o texto e a figura a esse respeito; como afirmado no texto, M. fisheri é um táxon difícil e foi podado a partir da análise mostrada na Figura 1 em conformidade.
https://doi.org/10.7554/eLife.49972.sa2 Artigo e informações do autor
Detalhes do autor
Torsten M Scheyer

John R Hutchinson
Financiamento
Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (149506)
- Torsten M Scheyer
Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (162775)
- Torsten M Scheyer
Generalitat de Catalunya (Programa CERCA)
- Massimo Delfino
Agência Estatal de Investigação (CGL2016-76431-P)
- Massimo Delfino
Programa-Quadro Horizonte 2020 (# 695517)
- John R Hutchinson
Os financiadores não tiveram nenhum papel no desenho do estudo, coleta e
interpretação dos dados, nem na decisão de enviar o trabalho para
publicação.
Agradecimentos
Agradecemos
à Alcadía Bolivariana de Urumaco e ao Instituto do Patrimônio Cultural
da Venezuela (IPC) pela autorização para coletar e estudar as amostras.
Hyram Moreno (MCNC), Gina Ojeda e Camilo Morón (UNEFM-CIAAP) e José
Hernández (Museu Paleontológico de Urumaco) por seu valioso apoio. Elena
Gavetti e Franco Andreone (Museu Regional de Ciências Naturais de
Torino) agradecem o empréstimo de material osteológico comparativo.
Austin Hendy (LACM), Maurice Joss (ZHDK), Daniel Núñez (PIMUZ) e Alex
Hastings (VMNH) são reconhecidos por discussões frutíferas. Paula Bona
(Universidade La Plata) e Rodolfo Salas-Gismondi (Universidade Peruana
Cayetano Heredia, Lima) forneceram imagens e informações sobre espécimes Caiman existentes . Pat Holroyd (Universidade da Califórnia em Berkeley) gentilmente forneceu imagens deMourasuchus atopus , imagens de Carl Mehling (AMNH) de Deinosuchus , imagens de Mariano Padilla Cano (MACV) de Palaeosuchus palpebrosus , enquanto Tyler Keillor (Univ. Chicago) é agradecido por fornecer imagens de Sarcosuchus imperator
. O programa TNT é gentilmente disponibilizado através do patrocínio da
Willi Hennig Society. Agradecemos à Crocodiles of the World (Brize
Norton, Reino Unido) por fornecer os Paleosuchus escaneadosespécime. Finalmente,
agradecemos aos revisores Michelle Stocker e Chris Brochu, os editores
John Long e Diethard Tautz, bem como Judith Recht pelos comentários que
levaram à melhoria do manuscrito. Este
estudo foi financiado pela Fundação Nacional de Ciência da Suíça
(doações 149506 e 162775 ao TMS) e pela Generalitat de Catalunya
(Programa CERCA) e pela Agência Estatal de Investigação Espanhola
(CGL2016-76431-P, AEI / FEDER, EU to MD). O
JRH foi financiado pelo Conselho Europeu de Pesquisa (CEI) no âmbito do
programa de pesquisa e inovação Horizonte 2020 da União Europeia
(contrato de concessão # 695517).
Editor sénior
- Diethard Tautz, Instituto Max-Planck de Biologia Evolutiva, Alemanha
Editor de Revisão
- John A Long, Universidade de Flinders, Austrália
Revisores
- Michelle Stocker
- Chris Brochu, Universidade de Iowa, Estados Unidos
Histórico da publicação
- Recebido: 5 de julho de 2019
- Aceito: 24 de outubro de 2019
- Versão do registro publicada: 27 de novembro de 2019 (versão 1)
direito autoral
© 2019, Scheyer et al.
Este artigo é distribuído sob os termos da Creative Commons Attribution License , que permite o uso e redistribuição irrestritas desde que o autor e a fonte originais sejam creditados.
Este artigo é distribuído sob os termos da Creative Commons Attribution License , que permite o uso e redistribuição irrestritas desde que o autor e a fonte originais sejam creditados.
Nenhum comentário:
Postar um comentário
Observação: somente um membro deste blog pode postar um comentário.