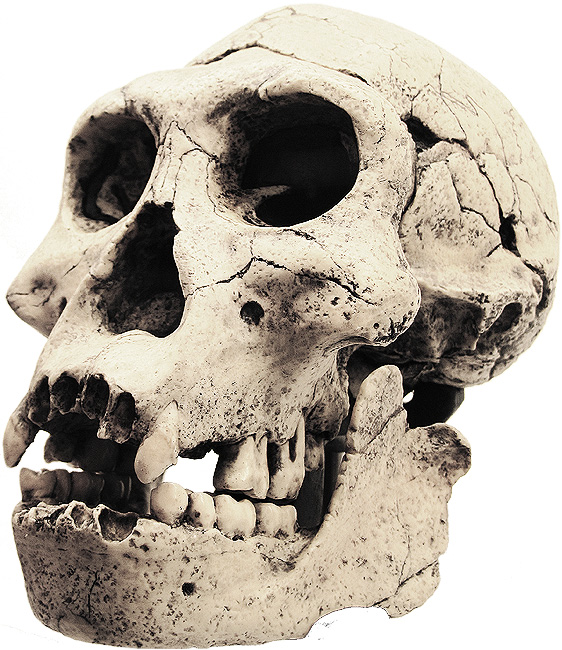
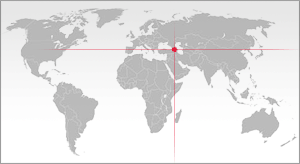
Homo erectus georgicus é o nome da subespécie usada
às vezes para descrever crânios e mandíbulas fósseis encontrados em
Dmanisi, na Geórgia. É conhecido como 'Dmanisi Man' e foi inicialmente
proposto como uma subespécie do Homo erectus, mas agora é classificado
como uma espécie separada. Um esqueleto parcial foi descoberto em 2001.
Homo erectus georgicus
Os fósseis têm cerca de 1,8 milhões de anos. Os restos foram descobertos
em 1991 por David Lordkipanidze. Implementos e ossos de animais foram
encontrados ao lado dos antigos restos humanos. O volume cerebral é de
aproximadamente 600 centímetros cúbicos - aproximadamente metade do
tamanho dos humanos modernos. É o menor cérebro hominídeo encontrado
fora da África, além do Homo floresiensis. Posteriormente, foram
encontrados quatro esqueletos fósseis, mostrando uma espécie primitiva
em seu crânio e parte superior do corpo, mas com espinhos e membros
inferiores relativamente avançados, proporcionando maior mobilidade.
Homo georgicus era bípede e diminutivo em tamanho - a altura masculina
média era de aproximadamente 4 pés. Padrões de desgaste dentário revelam
uma dieta onívora. Os sítios arqueológicos não mostram evidências do
uso do fogo.
Os fósseis de Dmanisi também podem representar um desenvolvimento social
e cultural significativo: um osso da mandíbula desdentada de 1,8 milhão
de anos de um caçador-coletor pode ser evidência de altruísmo e
compaixão - a sobrevivência neste momento, neste clima e nessa dieta,
não seguiu o padrão. regras estritas de adaptação evolutiva. Este
indivíduo foi cuidado por outros no grupo social.
O Homo georgicus pode representar uma ligação entre o Homo habilis e o
Homo erectus. Esta espécie pode ter sido o primeiro hominídeo a se
instalar na Europa devido a sua descoberta na costa oriental do Mar
Negro.
The Evolutionary Tree of Hominids - Millions of years before present
Fósseis atribuídos ao Paranthropus
aethiopicus foram encontrados em sítios da África Oriental que foram
datados entre 2,7 e 2,3 milhões de anos atrás (m.a.). Especificamente,
esta espécie foi encontrada na Etiópia (na bacia do rio Omo), no Quênia
(no oeste de Turkana) e na Tanzânia (em Laetoli). Fósseis de P.
aethiopicus exibem uma mistura de características na caixa craniana (as
partes do crânio que envolvem o cérebro) que se assemelham a Au.
afarensis com características faciais e dentários que são muito
semelhantes aos do Paranthropus boisei e Paranthropus robustus.
Juntamente com a sua idade (que sucede A. afarensis e precede P. boisei e
P. robustus), estas características do P. aethiopicus ajudaram os
cientistas a compreender as origens dos australopitecos robustos (P.
aethiopicus, P. boisei e P. robustus ). Consenso sobre as relações
evolutivas precisas entre estas espécies e entre estas espécies e
hominídeos anteriores, entretanto, não foi alcançado.
P. aethiopicus é conhecido apenas de restos do crânio;
nenhuma parte pós-craniana (partes do esqueleto que não inclua o crânio)
foi atribuída a esta espécie. Embora uma maxila e vários dentes (alguns
dos quais foram encontrados com osso associado da mandíbula inferior)
representando P. aethiopicus tenham sido encontrados, o fóssil mais
informativo é um crânio quase completo (crânio menos mandíbula inferior)
do Quênia. Este crânio em desdentados (desdentado) (número de catálogo
KNM WT 17000) é apelidado de “Crânio negro” porque os sedimentos em que
foi enterrado mancharam-no de preto.
O crânio de P. aethiopicus possui
uma mistura interessante de características que são mais semelhantes ao
Au. afarensis e características que mais se assemelham a P. robustus e
P. boisei. Os recursos compartilhados com o Au. afarensis incluem uma
face prognática (proeminente) e uma capacidade craniana relativamente
pequena (uma estimativa do tamanho do cérebro com base no volume do caso
cerebral; a capacidade cranial estimada de P. aethiopicus está na
extremidade inferior do intervalo de Au. afarensis) .
A morfologia
(tamanho e forma) das articulações temporomandibulares (a articulação
entre a mandíbula e o crânio) em P. aethiopicus e Au. afarensis também é
muito semelhante. Enquanto a dentição como um todo difere muito da
encontrada em Au. afarensis (veja abaixo), os dentes anteriores
(frontais) em P. aethiopicus, como os de Au. afarensis, são
relativamente grandes (comparados com aqueles encontrados nos
australopitais robustos). A crista sagital (uma crista óssea no topo do
crânio que se estende da frente para trás no meio do crânio, ao qual o
músculo temporal - um grande músculo mastigador que fecha a boca - se
liga) em P. aethiopicus também é semelhante àquela. encontrado em A.
afarensis.
Especificamente, a crista sagital de P. aethiopicus é mais
pronunciada na parte de trás do crânio, como em Au. afarensis, sugerindo
que, como A. afarensis, P. aethiopicus enfatizou a parte posterior do
músculo temporal; este fato é corroborado por outras semelhanças entre
Au. afarensis e P. aethiopicus nas marcações feitas no dorso do crânio
pelo músculo temporal. É importante notar que as características que P.
aethiopicus compartilha com Au. afarensis não são vistos em espécies do
gênero Paranthropus e estas características, em geral, não são vistas em
Australopithecus africanus.
Muitas características do crânio do P. aethiopicus se parecem mais
com aquelas exibidas por P. boisei e P. robustus. Por exemplo, os ossos
zigomáticos (bochecha) estão posicionados muito para frente e as margens
externas do rosto projetam-se bem para frente do meio da face, criando a
aparência de uma face “abaulada” (na qual as partes externas do projeto
da face tão à frente que eles obscurecem o buraco do nariz quando visto
de lado), característica dos outros australopitecos robustos. Como nos
demais australopitecos robustos, os ossos do palato de P. aethiopicus
são espessos. Os dentes pré-molares e molares em P. aethiopicus são
muito grandes - similares em tamanho aos outros australopitecos robustos
e muito maiores que em A. afarensis - e os pré-molares são muito mais
parecidos com os molares do que no A. afarensis.
A morfologia de P. aethiopicus tem uma relação direta com as
relações evolutivas entre as primeiras espécies de hominídeos. Em
particular, P. aethiopicus é importante para entender a origem dos
australopitecos robustos, bem como a relação dessas espécies com o Au.
afarensis e Au. africanus. Antes da descoberta do "crânio negro", os
pesquisadores sustentaram que Au. O africanus foi o ancestral de P.
robustus e P. boisei.
Esta descoberta, no entanto, lançou dúvidas sobre
este cenário porque representava uma espécie de hominina que era
contemporânea ao Au. africanus e que exibiram muito mais primitivo
(características compartilhadas encontradas no seu ancestral, neste caso
Au. afarensis) morfologia craniana. Alguns pesquisadores agora sugerem
que existiam duas linhagens distintas de australopitais robustos - uma
na África do Sul, representada por P. robustus, cujo antepassado,
segundo esse cenário, é o A. africanus, e outro na África Oriental,
representado por P. boisei, cujo antepassado é P. aethiopicus (cujo
antepassado, neste ponto de vista, é A. afarensis).
As semelhanças
entre P. robustus e P. boisei, desse ponto de vista, são evidências de
que ambas as espécies adquiriram independentemente características
relacionadas à mastigação de alimentos duros. No entanto, a maioria
filogenética (relacionada às relações evolutivas entre espécies) analisa
o grupo P. boisei e P. robustus juntos e sugere que o ancestral comum
dessas duas espécies era provavelmente mais derivado (possuindo
características não compartilhadas com seu ancestral, neste caso,
características compartilhadas com P. robustus e P. boisei) que P.
aethiopicus.
Independentemente da perspectiva que os cientistas adotam,
um consenso surgiu de que o registro fóssil de hominina entre 3,0 e 2,0
m.a. testemunhou um grau relativamente alto de homoplasia e reversão. Em
outras palavras, qualquer que seja o cenário filogenético correto, as
evidências atuais sugerem que algumas das características compartilhadas
por diferentes espécies não refletem relações evolutivas próximas
(homoplasia, por exemplo, características relacionadas à produção de
grandes forças mastigatórias, encontradas nos australopitecos robustos e ,
em certa medida, em A. africanus; ver ensaio sobre A. africanus para
mais detalhes) e algumas características evoluíram de um estado para
outro e depois de volta ao estado original (reversão).
Um registro
fóssil mais completo do intervalo de tempo entre 3.0 e 2.0 mya (em
particular, um registro fóssil melhor de P. aethiopicus) bem como um
melhor entendimento da diversidade morfológica incluída dentro de Au.
africanus - veja ensaio sobre Au. africanus) ajudará a esclarecer estas
questões filogenéticas não resolvidas.
PALEONTOLOGIA GERAL EXPLICA Atendendo aos pedidos!
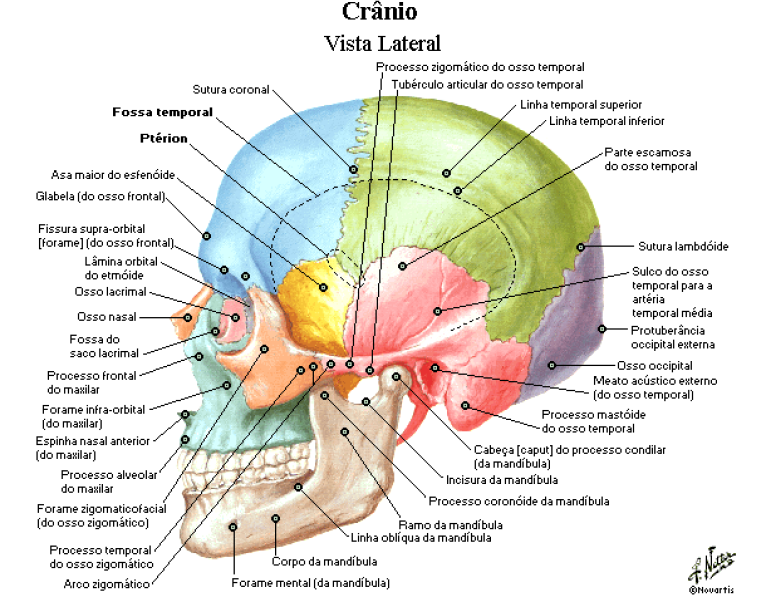
A maioria das articulações do crânio é do tipo fibrosa, apenas uma articulação é sinovial, sendo assim, dos 22 ossos do crânio, apenas um é móvel, a mandíbula.
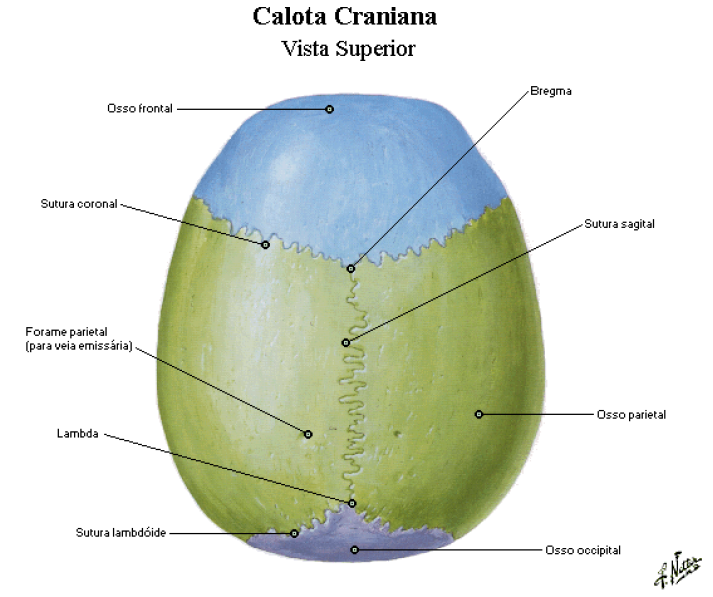
CALVÁRIA
É a parte superior do crânio, também conhecida como abóbada craniana.
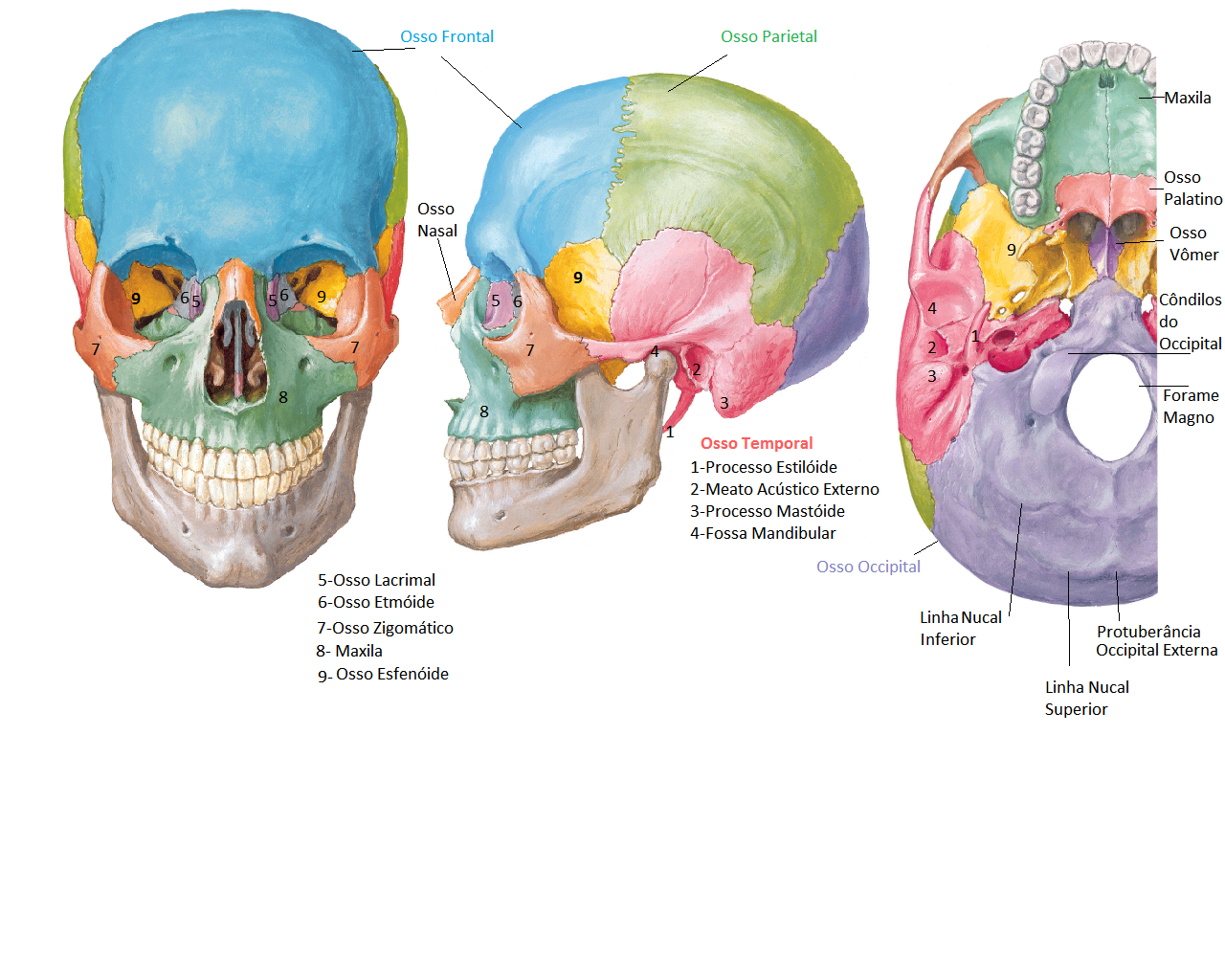
É constituída pelos ossos: parietais, frontal, occipital e temporal.
A calvária contém diversos forames diminutos,
por onde passam veias, conhecidas como veias emissárias, comunicando o
sistema venoso intracraniano com o sistema venoso extracraniano.
O revestimento externo da calvária é o couro cabeludo, formado por cinco camadas:
A camada mais externa é formada pela pele; subjacente a ela a camada de tecido conectivo denso; aponeurose (gálea aponeurótica); tecido conectivo frouxo, que permite a mobilidade das três camadas localizadas superficialmente, nessa camada localiza-se as veias emissárias; e o pericrânio.
As suturas são as articulações que promovem a união dos ossos da calvária (veja abaixo).
No crânio do recém-nascido, existe uma grande quantidade de tecido fibroso nessas áreas, formando as fontanelas ou fontículos do crânio. São seis fontículos:
Anterior (bregmático), posterior (lambdóide), ântero-lateral (esfenoidal) e póstero-lateral (mastóideo).
Quase todos os fontículos sofrem sinostose no primeiro ano de vida, sendo o fontículo anterior o que mais tarda a desaparecer, seu fechamento ocorre por volta do 2º ao 3º ano de vida.
Fontanelas ou fontículos
As fontanelas ou fontículos são espaços situados entre os ossos do crânio dos recém-nascidos e fetos. São vulgarmente chamados “moleiras“. Desaparecem quando se completa a ossifícação dos ossos do crânio.
No crânio do feto e recém-nascido, onde a ossificação ainda é incompleta, a quantidade de tecido conjuntivo fibroso interposto é muito maior, explicando a grande separação entre os ossos e uma maior mobilidade.
É isto que permite, no momento do parto, uma redução bastante apreciável do volume da cabeça fetal pelo “cavalgamento“, digamos assim, dos ossos do crânio. Esta redução de volume facilita a expulsão do feto para o meio exterior.
O
bebê ainda não atingiu o desenvolvimento completo do seu crânio no
momento do nascimento e as fontanelas permitem que o osso do crânio
continue a crescer até chegar ao seu tamanho adulto.
NETTER: Frank H.. Atlas de Anatomia Humana. 2ed. Porto Alegre: Artmed
Se
observar atentamente um crânio de feto em vista superior, um outro
fato pode ser notado: em alguns pontos a separação entre os ossos é
maior pela presença de maior quantidade de tecido conjuntivo fibroso.
tipos de fontanelas
Existem quatro tipos de fontanelas nos recém-nascidos:
Ântero-lateral (Esfenoidal);
Póstero-lateral (Mastoidea)
Essas duas primeiras são simétricas e se localizam de cada lado do crânio.
Posterior, que se fecha já no segundo mês de vida do recém-nascido.
Por fim, a fontanela Anterior, que se fecha mais tarde, por volta dos 18 meses.
Esta última é utilizada para avaliar a ocorrência de hipertensão craniana, bem como o estado de desidratação do bebê.
NETTER: Frank H.. Atlas de Anatomia Humana. 2ed. Porto Alegre: Artmed
Com o passar do tempo, os ossos da cabeça vão crescendo até que se encostam e “colam”
uns nos outros, fechando a fontanela quando a criança tem cerca de 1
ano e meio de idade. Se fechar antes de 1 ano, é necessário procurar o
pediatra.
A fontanela posterior é muito pequena e fecha perto dos dois meses de idade.
A fontanela anterior é maior, mede cerca de dois dedos. Contudo, o tamanho é muito variável entre bebés e fecha entre os 12e os 18 meses. Uma das suas características é a pulsação (batimentos rítmicos), originada pela proximidade de vasos arteriais.
Embora
já tenha sido descrita anteriormente, seria interessante lembrar que
na idade avançada pode ocorrer ossificação do tecido interposto (sinartrose), fazendo com que as suturas, pouco a pouco, desapareçam e, com elas, a elasticidade do crânio.
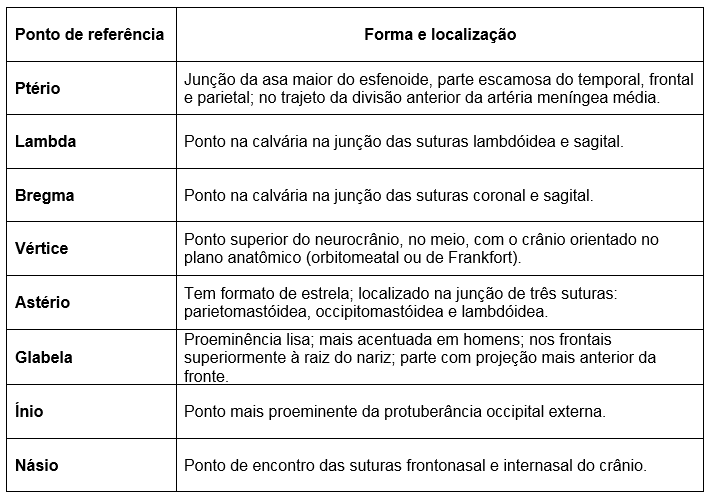
pontos craniométricos
Cada extremidade do fio da sutura sagital é identificada como um ponto ou área com o nome específico.
A extremidade anterior dessa sutura é denominada bregma;
E a extremidade posterior chama-se lambda;
Os ptérios direito e esquerdo são pontos de junção dos parietais, temporais e asas maiores do esfenoide;
Os astérios direito e esquerdo são pontos posteriores à orelha, onde as suturas escamosa e lambdóidea se encontram.
Tratam-se
de seis reconhecidos pontos ósseos usados em cirurgia ou outros casos
em que os pontos de referência específicos para medições cranianas são
necessários.
BONTRAGER: Kenneth L.; John P. Manual Prático de Técnicas e Posicionamento Radiográfico. 8 ed. Rio de Janeiro: Elsevier, 2015.
BONTRAGER: Kenneth L.; John P. Manual Prático de Técnicas e Posicionamento Radiográfico. 8 ed. Rio de Janeiro: Elsevier, 2015.
MOORE: Keith L. Anatomia orientada para a clínica. 7. ed. Rio de Janeiro: Guanabara Koogan, 2014.
Suturas
São
encontradas entre os ossos do crânio. A maneira pela qual as bordas dos
ossos articulados entram em contato é variável, reconhecendo-se:
Suturas planas = união linear retilínea ou aproximadamente retilínea, ex: internasal e interpalatina;
Suturas escamosas = união
em bisel (Corte enviesado na aresta de uma peça. O mesmo que
chanfradura = marca na pele provocada por corte de faca ou de navalha;
cicatriz), ex: temporo-parietal;
Suturas serreadas ou serrátil= união em linha “denteada”, ex: inter-parietal ;
Suturas esquindilese = Também chamada de cunha e sulco por assim se apresentarem às superfícies ósseas, ex: esfenóide e vômer.
NETTER: Frank H.. Atlas de Anatomia Humana. 2ed. Porto Alegre: Artmed
As articulações, ou juntas, do crânio são denominadas suturas e são classificadas como articulações (sinartroses) fibrosas.
A sutura coronal ou frontal separa o osso frontal dos dois ossos parietais;
A separação entre estes dois ossos na linha média é a sutura sagital;
Na parte posterior, a sutura lambdóidea separa os ossos parietais do osso occipital;
As suturas escamosas são formadas pelos cruzamentos inferiores dos dois ossos parietais com os respectivos ossos temporais.
NETTER: Frank H.. Atlas de Anatomia Humana. 2ed. Porto Alegre: Artmed
NETTER: Frank H.. Atlas de Anatomia Humana. 2ed. Porto Alegre: Artmed
Como visto anteriormente, as suturas são as articulações que promovem a união dos ossos da calvária.
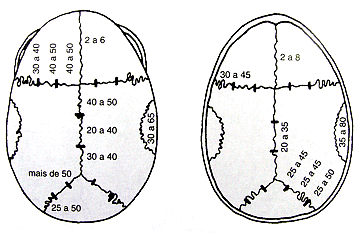
A sutura sagital localizada entre os ossos parietais (sofre sinostose entre 20º e 30º ano de vida).
Sutura coronal localizada entre os ossos frontal e parietais (sofre sinostose entre 30º e 40º ano de vida).
Sutura lambdóidea localizada entre os ossos occipital e parietal (sofre sinostose entre 40º e 50º ano de vida).
Estimativa de idade em anos para fechamento das suturas cranianas.
Esquerda: face externa. Direita, vista interna.
Referências Bibliográficas
BONTRAGER: Kenneth L.; John P. Manual Prático de Técnicas e Posicionamento Radiográfico. 8 ed. Rio de Janeiro: Elsevier, 2015.
MOORE: Keith L. Anatomia orientada para a clínica. 7. ed. Rio de Janeiro: Guanabara Koogan, 2014.
NETTER: Frank H. Netter Atlas De Anatomia Humana. 5.ed. Rio de Janeiro, Elsevier, 2011
SOBOTTA: Sobotta J. Atlas de Anatomia Humana. 21ed. Rio de Janeiro: Guanabara Koogan, 2000.
Homo erectus
Restos do Homo erectus são encontrados em toda a África e no oeste e leste da Ásia (a leste até a ilha de Java, na Indonésia). Outros fósseis, atribuídos por alguns estudiosos a essa espécie, foram encontrados na Europa, até o norte da Inglaterra. O Homo erectus teve um longo mandato; os fósseis mais antigos do Homo erectus são datados de cerca de 1,8 milhão de anos atrás, enquanto os mais jovens fósseis atribuídos a essa espécie datam de aproximadamente 300 mil anos atrás (ka). O Homo erectus é importante para o estudo da evolução humana porque é a primeira espécie a ser encontrada fora da África e apresenta muitas características anatômicas (especialmente o tamanho do cérebro e aspectos do esqueleto pós-craniano - isto é, as partes do esqueleto abaixo da cabeça) que refletem a evolução em direção ao padrão visto no Homo sapiens.
A taxonomia (a nomeação e atribuição de espécies) do Homo erectus é controversa. Alguns estudiosos afirmam que existem diferenças importantes entre os representantes asiáticos e africanos desta espécie. Em particular, esses cientistas afirmam que as características do crânio (crânio menos mandíbula inferior) - por exemplo, dentes grandes, quilhas sagitais (áreas estreitas de osso espesso que se estende desde apenas as sobrancelhas até a parte posterior do crânio) e maciço do neurocrânio ( a parte do crânio que cobre e protege o cérebro) e a face - são encontradas apenas em fósseis asiáticos de H. erectus.
Para esses paleoantropólogos, essa evidência sugere que as amostras asiática e africana representam espécies separadas; o nome "Homo ergaster" é dado aos fósseis africanos para formalizar essa distinção em nível de espécie. No entanto, como outros pesquisadores argumentam, muitas dessas características também são encontradas em alguns fósseis de H. erectus na África, sugerindo que toda a amostra constitui uma espécie única, o Homo erectus. Neste site (e para o restante deste ensaio), a última interpretação será empregada - ou seja, "Homo erectus" será usado para descrever a amostra inteira (africana e asiática).
Muitas das características que distinguem o Homo erectus de outras espécies de hominídeos - espécies anteriores e posteriores - são vistas no crânio. Estima-se que o tamanho médio do cérebro do Homo erectus tenha sido aproximadamente 900 centímetros cúbicos (cc), que é maior que o Homo habilis, mas menor que o do Homo heidelbergensis e outras formas posteriores. O tamanho do cérebro do Homo erectus é insignificantemente maior do que no Homo habilis quando é considerado em relação ao tamanho do corpo - isto é, o tamanho do cérebro aumentou substancialmente no Homo erectus, mas porque o tamanho do corpo também aumentou, o tamanho relativo do Homo erectus o cérebro não é consideravelmente maior que o do Homo habilis. O tamanho absoluto no cérebro, no entanto, causou mudanças no caso do cérebro; por exemplo, a caixa craniana é maior que no Homo habilis, mas menor que nas homininas posteriores.
A caixa craniana do Homo erectus também é muito longa em relação à sua altura, dando ao crânio uma forma de bola de futebol quando visto de lado. A caixa craniana e a face e as mandíbulas do Homo erectus eram muito pesadamente construídas, com ossos grossos e espessamentos extremos ao longo de algumas das suturas do crânio (onde dois ossos cranianos se conectam). Por exemplo, as sobrancelhas eram massivamente construídas e contínuas na face e grandes proeminências ósseas existiam na parte de trás do crânio (o toróide occipital e o torso angular). Devido a essas proeminências, o crânio do Homo erectus é pentagonal quando visto de trás, com a área mais ampla coincidindo com a parte inferior do crânio.
Além de seu uso para definir as espécies, os restos cranianos também foram usados para estudar a maneira como o Homo erectus cresceu e se desenvolveu até a idade adulta. Apesar do extenso estudo, os estudiosos discordam sobre como as evidências que envolvem este assunto devem ser interpretadas. Evidências dos dentes dos fósseis do Homo erectus sugerem que o Homo erectus ainda não havia desenvolvido o cronograma único de crescimento exibido pelo Homo sapiens. Usando evidências de crânio fóssil, no entanto, alguns cientistas afirmam que o H. erectus possuía características exclusivamente humanas, como o surto de crescimento juvenil. Assim, atualmente não está claro se o padrão de crescimento encontrado no Homo sapiens evoluiu no Homo erectus ou em uma espécie posterior.
The postcranial remains of this taxon have traditionally been interpreted as being very similar to those of Homo sapiens. The only notable difference is that the postcranial remains of Homo erectus are generally thicker and more massively-built than those of H. sapiens; all other features, though, point to striking similarities between the two species. The stature of Homo erectus, for example, has been suggested to be very similar to living humans and the hindlimb (leg) of Homo erectus
is much longer than in earlier forms for which good evidence exists.
Therefore, the proportion of the hind- to forelimb length (leg to arm)
in Homoerectus is considered to be almost identical to that of Homo sapiens. Because of these similarities, most scholars believe that Homo erectus was the first hominin species that was capable of a bipedal form of locomotion indistinguishable from that of Homo sapiens. Recent research, however, suggests that key fossils assigned to Homo erectus
were much shorter than previously thought and argues that modern human
stature may not have been present in Homo erectus. To wit, recent
fossil finds in Kenya suggest that Homoerectus was
quite variable in size and that sexual dimorphism (differences between
males and females within a species) may have been greater than
previously believed.
Whether it was due to its possession of human-like gait or not, Homo erectus is the first hominin species to have been found outside of Africa. Many sites bearing Homo erectus fossils have been discovered in China and Indonesia. In general, these sites are later than Homo erectus sites in Africa. At a site called Dmanisi in the Republic of Georgia, however, fossil assigned to Homo erectus have been found in sediments dated to 1.7 mya. The fossils at Dmanisi suggest that bands of Homo erectus
began migrating out of Africa at a time very close its origin.
Interestingly, though, the postcranial remains from Dmanisi are more
primitive (more similar to earlier species; e.g., the hindlimb was
relatively longer) than those found at many other sites.
The ability of Homo erectus to colonize land outside of
Africa was also largely dependent on the new technology this species
invented to deal with its surroundings. Homo erectus was the first species to actively control fire; the first good evidence for controlled fire is from a Homo erectus site in Israel and is dated to roughly 780 ka. Homo erectus
also invented new technology for making stone tools. The types of
tools used by this species are called the Acheulean industry.
The
Acheulean industry consists largely of handaxes (large, teardrop-shaped
artifacts) and cleavers (large artifacts with chips removed on either
side of one end to create a sharp edge), but other tool types are also
included. Handaxes are particularly emblematic of the Acheulean
industry. These artifacts are bifacially worked (chips removed from
both sides of the original rock), creating sharp edges on most
surfaces. Handaxes have been traditionally considered to be tools, but a
growing minority of scientists now argues that it was the flakes of
stones that were removed from the original rock that were the tools that
were ultimately utilized for hunting, butchery, and other tasks.
Handaxes are found much less abundantly at sites in eastern Asia. This
fact may be due to underrepresentation of Homo erectus site in
the region (due, in turn, to lack of scientific efforts in the region
and/or a real dearth of sites) or to the fact that other materials
(e.g., bamboo) that do not preserve in the archaeological record
replaced handaxes. It is also possible that Homo erectus lost the
tradition of making handaxes as it migrated from Africa to eastern
Asia. No consensus has been reached on this subject.
As relações evolutivas entre o Homo erectus e outras espécies hominíneas não estão totalmente resolvidas. Muitos estudiosos acreditam que o Homo erectus evoluiu do Homo habilis, provavelmente na África Oriental. Achados recentes no Quênia, no entanto, aumentaram o alcance temporal do Homo habilis, sugerindo que as duas espécies se sobrepunham muito no tempo e fazendo com que alguns cientistas questionassem a ligação direta entre o Homo habilis e o Homo erectus. É geralmente aceito, no entanto, que o Homo heidelbergensis evoluiu do Homo erectus na África e se espalhou pelo Velho Mundo substituindo populações posteriores do Homo erectus, da mesma forma que o Homo sapiens substituiu o Homo heidelbergensis e o Homo neanderthalensis três quartos de milhão de anos. mais tarde (ver ensaios sobre Homo sapiens e Homo neanderthalensis).
O registro arqueológico do Pleistoceno Superior / Holoceno Superior no Brasil: um banco de dados georreferenciado
Entre 13.000 e 8.000 14C BP, o leste da América do Sul foi
colonizado por uma população estável e diversificada de
caçadores-coletores.
A escavação arqueológica nos últimos vinte anos tem
produzido evidências cada vez mais consistentes de ocupação em
diferentes regiões do Brasil desde o final do Pleistoceno, com datas
pelo menos contemporâneas ao Clovis Horizon na América do Norte. Isto é
abordado documentando e analisando a quantidade, qualidade e
distribuição das datas 14C arqueológicas do Brasil durante este período.
Um total de 277 datas de 90 sites são tabulados, mapeados e incluídos
na análise. Durante o Pleistoceno Superior houve uma fase pioneira de
colonização humana, com dispersão para o interior através dos principais
sistemas fluviais.
Posteriormente, o Holoceno Primitivo viu a primeira
fase de assentamento estabelecido do interior do Brasil. Parece haver um
limiar arqueológico atingido em ca. 10.500 anos 14C BP: o número de
sites aumenta, há evidências de liquidação de todos os principais biomas
e há evidências claras de diversidade cultural inter-regional.
Oldest non African Stone tools found in China
July 14, 2018
Um artigo científico e um comentário na
edição de 11 de julho da revista Nature relatam a descoberta no sul da
China de ferramentas de pedra datadas de 2,1 milhões de anos atrás.
Por
muitos anos, a mais antiga evidência de ancestrais humanos fora da
África foi encontrada em Dmanisi, na República da Geórgia, e datada de
cerca de 1,8 milhão de anos atrás (mya).
O trabalho, de autoria de Z. Zhu, do Laboratório de Geologia Marítima e
Marítima, Instituto de Geoquímica de Guangzhou, Academia Chinesa de
Ciências, Guangzhou, China e seus colegas, pode ser encontrado
“Foi dada atenção considerável
ao namorar a primeira aparição de hominídeos fora da África. A mais
antiga evidência esquelética e artefactual para o gênero Homo na Ásia
atualmente vem de Dmanisi, na Geórgia, e é datada de aproximadamente
1,77 a 1,85 milhões de anos atrás (Ma) 1.
Dois incisivos que podem
pertencer ao Homo erectus vêm de Yuanmou, sul da China, e são datados de
1,7 Ma2; a evidência mais antiga é um crânio de H. erectus de Lantian
(Gongwangling) - datado recentemente de 1,63 Ma3 - e os primeiros
fósseis de hominídeos da cúpula de Sangiran em Java, datados de cerca de
1,5 a 1,6 Ma4.
Os artefatos de Majuangou III5 e Shangshazui6 na bacia
Nihewan, norte da China, também foram datados de 1,6 a 1,7 Ma. Aqui
relatamos um Pleistoceno Inferior e uma seqüência de artefatos em grande
parte contínua de Shangchen, que é uma localidade paleolítica
descoberta recentemente no Platô de Loess do sul da China, perto de
Gongwangling no condado de Lantian.
O local contém 17 camadas de
artefatos que se estendem desde o paleossolo S15 - datado de
aproximadamente 1,26 Ma - até o L28, que damos cerca de 2,12 Ma. Esta
descoberta implica que os homininos deixaram a África mais cedo do que o
indicado pelas evidências de Dmanisi. ”
Antes de ler este artigo, no entanto, pedimos aos leitores interessados que visitem o comentário de John Kappelman intitulado “Ancient Hiominin Arrival in Asia”.
Este comentário examina todos os aspectos relevantes da datação de
artefatos, tipos de ferramentas, uso de ferramentas, etc. De
fotografias, as ferramentas parecem ser da tecnologia de Olduwan (veja
“Craftsmanship Antecipado, disponível na Home Page em becomehuman.org).
Não houve avanço na complexidade de fabricação de ferramentas até a
chegada das ferramentas biface em torno de 1,6 mya. Kappelman, em seu
comentário, discute a estimativa de quanto tempo levaria os povos
caçadores-coletores a migrar da África para a China.
Deve-se notar que nenhum osso foi encontrado com esses artefatos. Os
autores e comentaristas assumiram que os fabricantes de ferramentas eram
o Homo erectus e não um Australopitecino contemporâneo. Se assim for,
esta é uma data muito anterior, em 300.000 anos, para o H. erectus fora
da África. Kappelman também discute as forças push-pull que atuam sobre
os povos migrantes: empurrado por trás pela expansão das populações;
puxado pelos recursos do jogo, que também estão migrando.
terça-feira, 28 de agosto de 2018
Biologia evolutiva do desenvolvimento explica origem das formas biológicas
Disciplina procura entender como ocorre a variação entre as espécies que gera toda a diversidade da vida
Integrada
com biologia molecular, evolução, genética, entre outras áreas, a
biologia do desenvolvimento estuda organismos como os tunicados – Foto:
Wikimedia Commons
.
Embora tenha surgido há aproximadamente 3,8 bilhões de anos, a vida na
Terra ainda tem muito o que aprender sobre suas próprias origens. Foi
somente no século 19, durante a famosa viagem do navio HMS Beagle à
América do Sul, que o cientista inglês Charles Darwin esboçou o que
ficaria conhecido como a teoria da evolução por meio da seleção natural.
Para o naturalista, o conceito de seleção natural envolveria entender
como algumas características que são hereditárias se tornaram mais
comuns em gerações sucessivas de uma população de organismos que se
reproduzem e, ao mesmo tempo, que outras características tornam-se menos
comuns.
Pouco mais de cem anos desde a apresentação da hipótese para
cientistas do mundo todo, a teoria é uma das bases do campo que ficou
conhecido como biologia evolutiva do desenvolvimento (em inglês evo-devo). A subdivisão da biologia estuda a origem e a descendência das espécies, bem como suas mudanças ao longo do tempo.
Apesar de ter começado no continente, o estudo da evolução das
espécies feito nos países latino- americanos ainda busca visibilidade
internacional. Não por acaso, o professor do Departamento de Zoologia do
Instituto de Biociências (IB) da USP, Federico David Brown Almeida,
especializado na área da evo-devo, foi o responsável por uma edição especial do Journal of Experimental Zoology Part B., lançada no início deste ano.
A edição, na qual Brown foi editor convidado, foi dedicada à pesquisa
em biologia evolutiva do desenvolvimento na América Latina.
“A biologia do desenvolvimento é uma disciplina relativamente nova que começou nos anos 1980 como consequência do boom
molecular”, explica ele. Foi somente nas últimas duas décadas do século
20 que o desenvolvimento ganhou destaque maior no campo da evolução, a
partir da descoberta de genes que regulam o desenvolvimento embrionário
nos chamados “organismos-modelo”. A descoberta, de acordo com o docente,
permitiu aos cientistas estudar genes em vários grupos animais.
“É uma disciplina que se integra com biologia molecular, evolução,
genética, entre outras áreas mais velhas da biologia”, esclarece ele, ao
enfatizar que a pergunta principal da disciplina é explicar como se
originam as formas biológicas. Ou seja, entender quais são suas origens
do ponto de vista genético do desenvolvimento e “como ocorre a variação
entre os animais para gerar toda a diversidade que a gente tem na vida”,
ilustra o professor.
Visibilidade na América Latina
Foto: Reprodução
De acordo com Brown, a edição especial foi o resultado de uma
conferência que aconteceu na Califórnia há pouco mais de dois anos.
Durante o evento, se formou a Sociedade Pan-Americana de Biologia
Evolutiva do Desenvolvimento, com pesquisadores norte- americanos e
latino-americanos.
“Eu estava envolvido na formação desse novo grupo e os outros
pesquisadores me convidaram para organizar um workshop da América Latina
para entender por que a área está tão pouco representada na nossa
região”, relembra Brown.
A partir do encontro, participantes latino- americanos solicitaram um
espaço aos editores das principais publicações da área para apresentar
seus trabalhos e dar maior visibilidade à pesquisa da região. Como
editor associado, Brown, que na USP é líder do Laboratório de Evo-Devo
do IB, foi atrás de pesquisadores no Brasil e na América Latina que
compuseram um mosaico de tudo que é produzido no continente.
No volume foram incluídos revisões, artigos históricos e diversos
trabalhos experimentais. “Tratamos desde a parte epistemológica, de
história da ciência, até propostas novas e trabalhos experimentais com
animais da nossa região”, sumariza ele.
Ao todo, seis países contribuíram com os 13 artigos do volume.
Brasileiros colaboraram com um artigo introdutório e uma revisão
histórica de evo-devo na América Latina, além de uma série de
estudos funcionais e do desenvolvimento de uma estrutura anatômica
(filamentos pélvicos) do peixe pulmonado da América do Sul (peixe
amazônico) e uma análise do desenvolvimento dos olhos em castas de
abelhas, feita por especialistas da USP, no campus de Ribeirão Preto.
Federico David Brown Almeida, do Instituto de Biociências (IB) da USP – Foto: Reprodução
No geral, os artigos focam em organismos característicos do
continente, que são “pouco estudados, interessantes e originais por
isso”, explica o professor.
“Foi bem interessante porque incluímos tanto plantas quanto animais,
já que temos, na América Latina, grupos fortes nas duas áreas”, revela
Brown ao salientar que não é comum na evo-devo unir botânica e
zoologia em um mesmo volume. ”A biologia do desenvolvimento estava
dividida e foi predominantemente dominada por modelos animais. E a
comunidade de plantas, que estudava Biologia do Desenvolvimento, estava
associada à áreas mais fisiológicas”, lembra ele.
No entanto, ao trazer a botânica para o volume, o professor defende
que aproximar os campos provoca um enriquecimento da pesquisa na área.
“Em plantas temos grupos que estão estudando a identidade das estruturas
florais, a diversidade da arquitetura floral, por exemplo”, destaca.
Por meio de estudos da nossa flora, especialistas perceberam que alguns
modelos estabelecidos para plantas já não se mantêm mais e estão em
constante mudança.
Laboratório de evo-devo
Formado por um grupo relativamente pequeno, o chamado Laboratório de
Evo-Devo do IB existe faz três anos. Seu projeto principal busca
entender a origem evolutiva das células-tronco. Nesse campo,
pesquisadores também procuram compreender os chamados processos de
regeneração. “Queremos saber como essas células evoluíram e como as
mudanças nos processos de células-tronco podem estar envolvidas em
processos regenerativos de alguns animais”, explica o especialista.
“Estamos usando vários invertebrados, sobretudo tunicados, um subfilo
de animais marinhos”, elabora Brown. Os tunicados são animais que
evolutivamente estão próximos dos vertebrados e portanto compartilham
muitas similaridades em seu desenvolvimento inicial. “Neste grupo,
depois do desenvolvimento larval, você tem várias espécies coloniais que
evoluíram. Vários grupos dentro desse filo são coloniais, então eles
conseguem se clonar formando indivíduos diferentes, como as plantas”,
esclarece o professor. .
Espécie de tunicado – Foto: Wikimedia Commons
.
Apesar de serem um grupo de animais que se desenvolvem como vertebrados,
os tunicados conseguem ter o que os especialistas chamam de
plasticidade e se reproduzem assexuadamente por brotação.
“Queremos entender como nós perdemos essas capacidades regenerativas,
essas capacidades de brotar, regenerar, que tinham nossos ancestrais”,
revela Brown. Alternativamente, o objetivo da pesquisa atual do
laboratório é entender como evoluíram as capacidades regenerativas e de
brotamento neste grupo específico de animais.
O laboratório atualmente é formado por apenas oito membros.
Uma teoria unificada
“Foram três concursos para o Departamento de Biologia do
Desenvolvimento da USP até a chegada de um candidato apropriado”,
recordou Brown, que veio do Equador para trabalhar no Brasil. “Somos
poucos os que atuam nessa linha de pesquisa na América Latina”, pontua.
A área, que é forte tanto nos EUA quanto na Europa, possui uma grande
tradição no velho continente, onde se formou a primeira sociedade de evo-devo do mundo.
Foto: Reprodução
Entretanto, “o estudo começou lá, mas o objeto foi estudado aqui na
América Latina”, reforça o pesquisador, ao salientar que a proposta da
edição especial é deixar claro para os demais cientistas da área que a
América Latina tem contribuído com várias novas ideias sobre evolução e é
um continente que pode explorar sua diversidade natural para continuar
fazendo descobertas. “Os modelos de animais e plantas que temos aqui são
inumeráveis e com adaptações fenomenais, coisas realmente únicas que
precisam de mais visibilidade e investimento local”, aponta ele.
A área da Biologia do Desenvolvimento, por ser relativamente nova,
ainda conta com poucos laboratórios já estabelecidos. E, na América
Latina, os poucos locais de estudo também sofrem com a falta de
comunicação.
“Estamos acostumados a colaborar e interagir com pesquisadores do
chamado Primeiro Mundo, mas não entre nós. Nos falta esse tipo de
interação para gerar um grupo forte”, teoriza o docente.
O Journal of Experimental Zoology Part B. publicou artigos
de vários grupos da América Latina, dentre eles, as pesquisas de Augusto
Flores, Gabriel Marroig, Hussam Zaher, Klaus Hartfelder, Márcia
Bitondi, Miguel Rodrigues, Nanuza Luiza de Menezes, Tiana Kohlsdorf e
Zilá Luz Simões, especialistas da USP
O futuro, para o professor Brown, está em formular, junto com
pesquisadores da Ecologia, uma teoria de como o ambiente está envolvido
nos processos do desenvolvimento, como ele pode influenciar esse
desenvolvimento e como esses desenvolvimentos passam por processos
adaptativos. A partir daí, especialistas em evo-devo poderão
trabalhar com base em uma “teoria nova, sólida e forte”, que unirá a
área e ampliará ainda mais suas possibilidades de novas descobertas